羌塘高原降水梯度带紫花针茅叶片氮回收特征及影响因素
2016-08-09赵广帅熊定鹏石培礼冯云飞武建双张宪洲曾朝旭
赵广帅,熊定鹏,石培礼,冯云飞,武建双,张宪洲,曾朝旭
1. 中国科学院地理科学与资源研究所, 生态系统网络观测与模拟重点实验室,北京 100101 2. 中国科学院大学,北京 100049
羌塘高原降水梯度带紫花针茅叶片氮回收特征及影响因素
赵广帅1,2,熊定鹏1,2,石培礼1,*,冯云飞1,2,武建双1,2,张宪洲1,曾朝旭1,2
1. 中国科学院地理科学与资源研究所, 生态系统网络观测与模拟重点实验室,北京100101 2. 中国科学院大学,北京100049
摘要:植物回收衰老叶片的氮是植物重要的养分保持和环境适应机制,在寒旱贫瘠的生境更是如此。为了理解降水梯度上植物对高寒贫瘠环境的养分适应特征,研究了羌塘高寒草原优势物种紫花针茅叶片氮回收策略及其与环境因子的关系。结果表明,降水梯度带上紫花针茅叶片具有较高的叶氮水平和氮回收能力。生长季盛期紫花针茅绿叶平均氮含量为(23.87±3.92)g/kg,高于中国草地平均水平(20.9 g/kg)及全球平均值(20.1 g/kg);绿叶氮含量与年降水量(MAP)呈显著负相关,干旱端(西部)绿叶中氮含量明显高于湿润端(东部)。枯叶养分回收后的氮水平(NRP)很低,平均为(6.76±1.42)g/kg,叶片平均氮回收效率(NRE)为(71.25±6.46)%,明显高于中国温带草原和全球的平均水平(46.9%—58.5%)。枯叶中氮回收水平对叶片氮回收效率起决定作用,是维持高养分回收效率的物质基础。NRE与MAP、土壤全氮(TN)和土壤无机氮呈显著负相关;NRP与TN相关性不显著,但与土壤无机氮显著负相关。尽管NRE与NRP呈显著负相关,但二者与绿叶氮含量均没有显著相关性。年均气温、海拔对NRE和NRP影响均不显著。因此,紫花针茅叶片极高的NRE和低NRP反映了它对极端干旱贫瘠环境的养分保持能力,通过内部氮循环来降低养分流失。土壤氮的有效性是影响紫花针茅叶片氮回收能力的关键因子,降水通过影响土壤氮的有效性以及绿叶中氮含量间接影响紫花针茅叶片氮回收效率。
关键词:羌塘高原;降水梯度带;养分回收效率;土壤氮;叶片氮含量
氮是限制植物生长和群落生产力的基本营养元素,在高寒和干旱生态系统更是如此。植物对限制元素的保持通常采取保守策略,植物一方面通过增加根系比例以增加吸收获取养分,另一方面从衰老组织中回收和再利用一部分养分。植物回收衰老叶片中的氮素是一种重要的养分保持机制,确保植物保存和再利用这些养分[1- 3]。在养分贫乏生态系统中,植物的养分需求很大程度是依赖于植物的内循环,即养分的回收[1]。长期以来,叶片氮回收的生态过程及其环境梯度上的空间格局倍受关注,对阐释植物的养分保持策略和对贫瘠环境的适应具有重要意义。
叶片氮回收效率(NRE)和氮回收水平(NRP)是衡量养分回收机制的两个重要指标[4- 6]。已有研究表明,生长在养分贫瘠环境中的植被比肥沃环境中的植被具有更高的NRE或更低的NRP[4, 7-8],原因在于氮的回收主要由土壤氮的有效性[9- 11]或植物氮水平[12-13]决定,在养分有效性低的环境中植物的养分利用趋于保守策略。通常情况下,土壤氮的有效性越高,植物叶片氮含量越高,凋落物的质量也高,而NRE就低,叶片氮回收能力和土壤氮有效性显著负相关[14-15],NRE随绿叶氮含量的增加而降低[5,16]。
土壤氮的有效性与降水量和土壤水分状况存在显著正相关,降水越多的区域土壤氮的有效性越高[17-18]。例如,年平均降水量(MAP)能够解释潘塔哥利亚草地94%净氮矿化的空间变异[19],并且土壤无机氮(铵态氮和硝态氮)与MAP存在显著的正相关[20]。同时,植物叶片氮含量也随降水量上升而增加[21-22]。在降水梯度带上,养分回收效率存在着由干旱端向湿润端递减的趋势[23-24]。Yuan等[6]在综合分析大量前人研究结果的基础上得出NRE随MAP的增加而降低,许多学者通过控制实验也得到类似的结果[9,25]。然而,有些学者研究发现随着MAP增加,土壤氮的有效性不变,甚至是下降[20,26]。尽管氮矿化能力随降水增加而逐渐增加,但植物净初级生产力、有机质以及微生物量等的增加也会导致土壤微生物氮固持增加[27-28]。因此,在降水梯度带上随着水分的增加,氮限制可能逐渐增强,植物叶片氮回收能力随降水梯度带的递变发生逆转[29-30]。植物叶片氮回收能力随水分梯度的变化规律在不同生态系统并不一致。
羌塘高原是西藏高原的主体,平均海拔在4400m以上,气候寒冷干燥,土壤贫瘠,是生态环境最为脆弱的区域。羌塘高原的植被类型较为简单,其优势植被是以紫花针茅 (Stipa purpurea)为优势种的高寒草原[31]。高寒植物适应高寒、贫瘠环境表现出高效的氮利用策略[2]。羌塘高原由东向西存在明显降水梯度带,MAP从最东部700mm左右降低到阿里地区噶尔县的50—80mm左右,在此环境梯度上土壤有机质从4.0%左右降低到1.0%以下,土壤全氮(TN)含量从0.2%降低到0.02%左右,水分及与其密切相关的养分梯度对高寒草原物种分布和群落结构产生更重要影响[32]。目前,青藏高原开展的养分利用和叶片氮回收效率的研究都局限于站点水平,特别是优势植物在环境梯度上的养分回收效率变化规律的研究还很缺乏。为此,人们关注羌塘高原植被优势种紫花针茅在生长季末叶片氮回收效率是如何变化?主要影响因子是什么?
为了研究羌塘高原降水梯度带上紫花针茅对高寒、贫瘠环境适应策略,提出以下假设:(1)羌塘高原土壤氮的有效性和紫花针茅叶片氮含量随降水增加而增加;(2)紫花针茅叶片具有高的氮回收能力,并且从东往西随降水减少不断增加。此外,除了降水、土壤氮的有效性和植物氮素状况对植物叶片氮回收能力产生影响外,气温[6]、海拔[13]等也可能对其产生影响。因此,本文也进一步分析了植物叶片氮回收能力与气温、海拔的关系,揭示生长季末叶片氮回收的关键影响因子。羌塘高原降水梯度带植被生长季末叶片养分回收策略研究不仅可弥补高原养分回收数据的稀缺,也可为理解极端环境下植被养分利用策略提供依据。
1材料与方法
1.1研究区概况
羌塘高原位于西藏自治区西北部,南起冈底斯—念青唐古拉山脉,北抵昆仑山脉,东迄青藏公路西侧E91°左右的内、外流水系分水岭,西止于国境线,面积约60万km2,行政区划上属西藏自治区的那曲与阿里两地区管辖。羌塘高原是我国气候条件最为恶劣、生态环境极为脆弱的典型区域,环境特点主要表现为气候寒冷、干旱、多风,地表沙物质丰富,植被稀疏低矮等,生态容量较低。羌塘高原干旱气候特征显著,蒸发强度普遍>1800mm,年均风速多在3m/s以上,年平均干燥度指数1.6—20[33]。羌塘高原气候寒冷,大部分地区年平均气温< 0 ℃,最冷月(1月)均温为-10—-18 ℃,而最暖月(7月)均温不及14 ℃,全年> 0 ℃的活动积温多在1200℃以下[34]。
1.2野外取样和样品分析
2014年7月至10月期间,沿羌塘高原降水梯度带(那曲县罗玛镇至改则县)共选择12个站点进行紫花针茅叶片和土壤等样品采集,所有站点都位于地势平坦,植物生长良好的地带性植被区域,相邻站点间隔50—80km,并且各站点植物群落以紫花针茅为优势种,确保取样点的代表性。在7月底至8月初,草地生物量达到最大的生长季盛期,由东向西在每个站点随机选择100株左右生长良好的紫花针茅成熟的全展绿叶,每20株样品作为一个重复,共5次重复。同时,在每个站点随机采集3个土壤剖面,土壤剖面间距离500m以上。每个土壤剖面采集0—10cm和10—20cm土层土壤样品,样带上共采集72个土壤样品。9月底至10月初,在草地生长季结束时再在每个相应站点随机选择100株左右紫花针茅,以同样的重复方式采集枯黄的当年生枯叶。

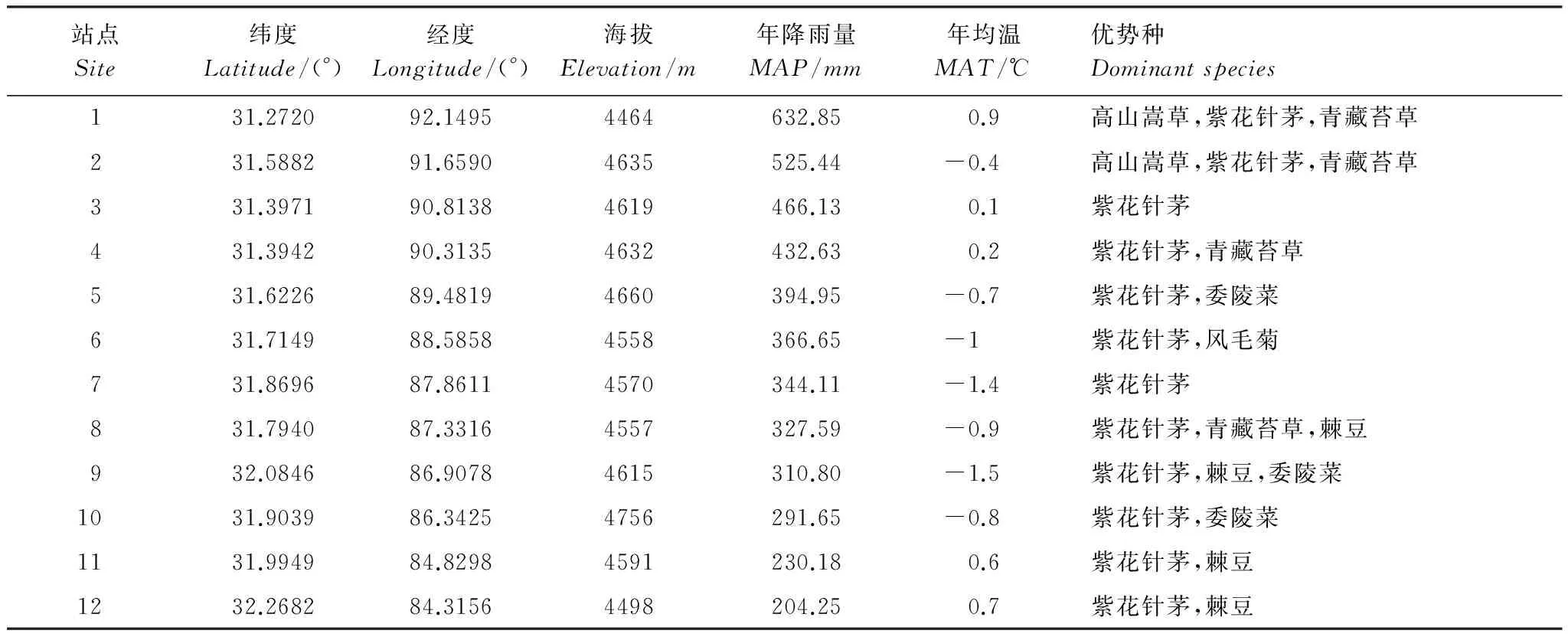
表1 采样点位置及环境特点
1.3数据处理与分析
叶片氮回收效率(NRE)被定义为生长季盛期绿叶、生长季末当年生枯叶间氮含量减少的相对比例,具体计算如下:
NRE=((Ng-Ns)/Ng)×100%
其中,Ng(g/kg)为绿叶的氮含量,Ns(g/kg)为枯叶的氮含量。
叶片氮回收水平(NRP)被定为生长季末当年生枯叶中的氮含量(Ns),它也是反映植物养分回收能力的直接指标[4]。此外,降水和气温数据来自国家气象站和中国科学院拉萨高原生态综合试验站在藏北羌塘高原设置的HOBO自动气象观测站。利用线性回归方法分析植物叶片氮回收能力与降水、气温、土壤养分、叶氮状况等因子之间的关系,显著性水平P<0.05,所有统计分析均在SPSS19.0 (SPSSInc.,Chicago,IL,USA)中进行。同时,为了准确区分降水、土壤养分和叶氮含量等因素对叶片回收效率的影响贡献率,利用结构方程模型进行路径分析(Amos17.0,SPSSInc.,Chicago,IL,USA),计算变量间路径或载荷系数(即回归系数),并通过CR(CriticalRatio)值检验统计结果的显著性。
2结果与分析
2.1紫花针茅叶氮回收效率及其影响因子的空间变异
羌塘高原草地地上净初级生产力(ANPP)与MAP呈极显著的正相关,降水平均每增加100mm,ANPP约增加76.6gm-2a-1(图1)。平均的土壤全氮(TN)含量为(1.23±0.59)g/kg,土壤无机氮含量为(0.02±0.01)g/kg,TN与MAP呈极显著正相关(图1),无机氮与MAP也呈显著正相关,降水越多的地方,土壤全氮和有效氮含量相对越高。
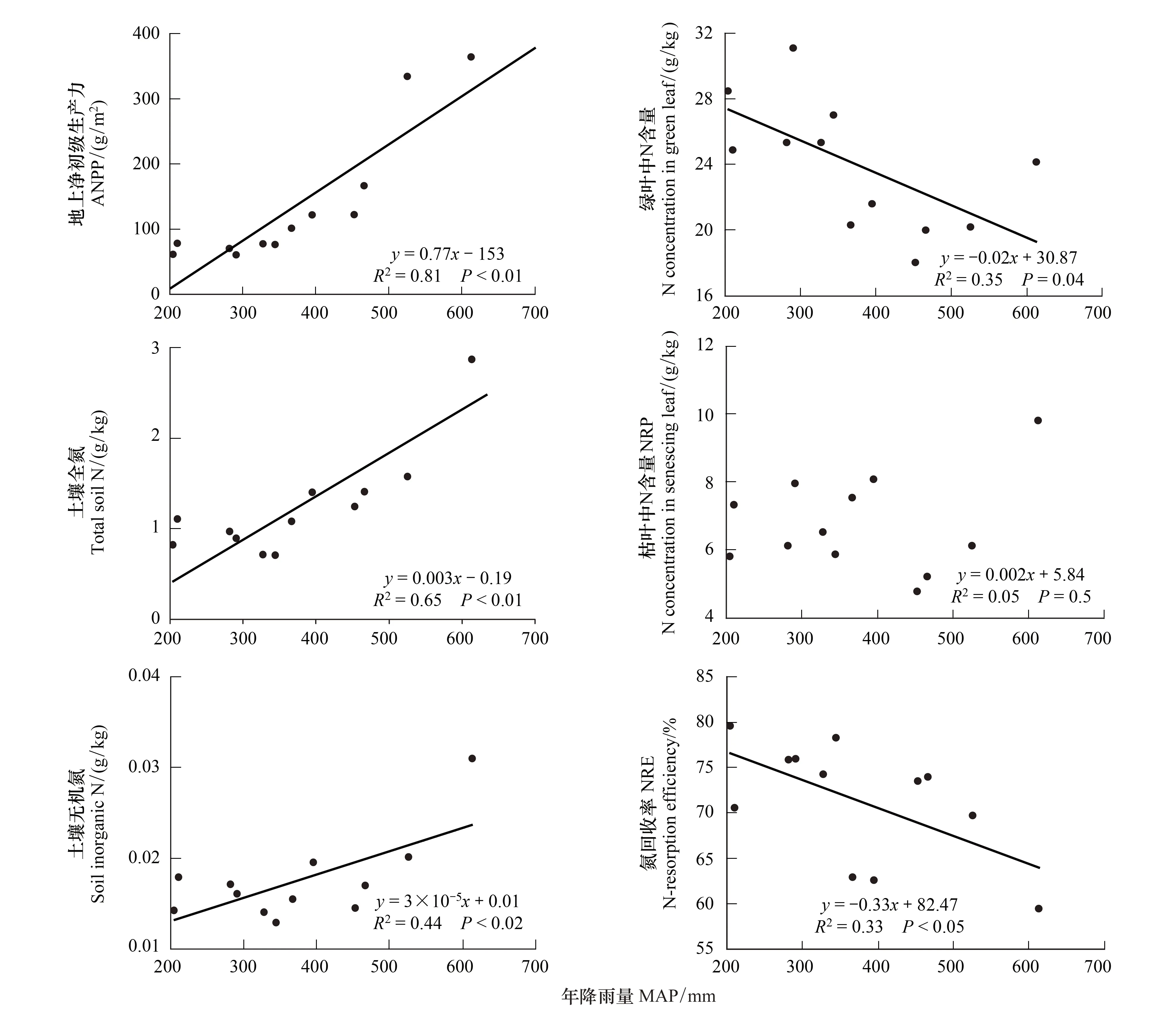
图1 羌塘高原降水梯度带土壤氮含量、叶片氮浓度及回收效率变化格局Fig.1 The changes of soil N, leaf N concentration and N resorption efficiency along the precipitation gradient on the Changtang Plateau
生长季盛期绿叶平均氮含量为(23.87±3.92)g/kg,绿叶氮含量与MAP呈显著负相关关系,干旱端(西部)绿叶中氮含量明显高于湿润端(东部)(图1)。水分梯度带上紫花针茅叶片氮回收效率(NRE)存在明显空间变化,由最东端的59%增加到最西端的80%,样带上平均NRE为(71.25±6.46)%,NRE与MAP显著负相关,随降水增加NRE不断降低(图1)。整个水分梯度带平均的枯叶氮含量(NRP)为(6.76±1.42)g/kg,NRP与MAP没有显著相关性(图1)。
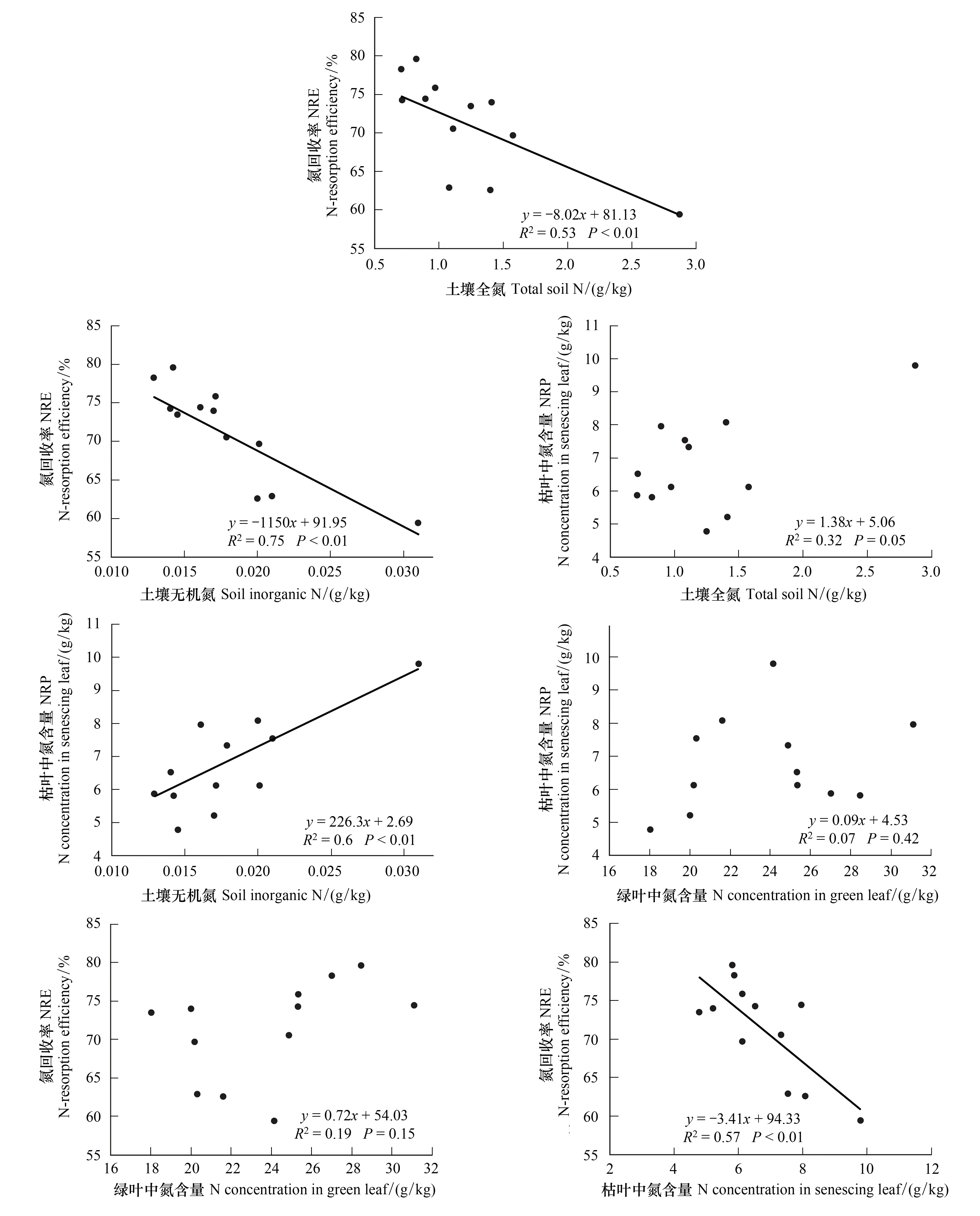
图2 紫花针茅叶片氮回收效率与土壤和叶片氮含量的关系Fig.2 The correlation of leaf nitrogen resorption efficiency of Stipa purpurea with soil N content and leaf N concentration
2.2叶氮和土壤氮含量对紫花针茅叶氮回收能力的影响
叶片NRE与土壤TN呈极显著的负相关,特别是与土壤无机氮相关性更强;NRP与TN没有相关关系,但与土壤无机氮呈极显著正相关(图2)。绿叶氮含量分别与NRP、NRE没有相关性,但NRP与NRE呈极显著负相关(图2)。
2.3气温和海拔对紫花针茅叶氮回收能力的影响
羌塘高原降水梯度带年平均气温(MAT)较低,大部分< 0℃,海拔较高,普遍在4500m以上。NRE(R2= 0.03, P=0.59)和NRP(R2= 0.00, P=0.92)与MAT没有显著相关性;同时,海拔对NRE(R2= 0.04, P=0.53)和NRP(R2= 0.13, P=0.24)也没有显著影响。
2.4不同因素对氮回收效率的贡献率
综合以上线性回归结果,利用结构方程模型分析降水、土壤养分和叶氮含量等因素对叶片氮回收效率的影响贡献率,结果表明MAP对土壤TN和绿叶中氮含量的直接影响最显著(0.81,-0.91),而对其他因子的直接影响很小;土壤TN对NRP直接影响最显著(0.98),而对其他因子的直接影响很小;绿叶中氮含量和NRP对NRE的直接影响最显著,标准路径系数分别为0.75、-0.94(图3)。以上结果说明土壤氮的有效性是影响羌塘高原紫花针茅叶片氮回收能力的关键因子,降水通过影响土壤氮的有效性以及绿叶中氮含量间接影响紫花针茅叶片氮回收效率。
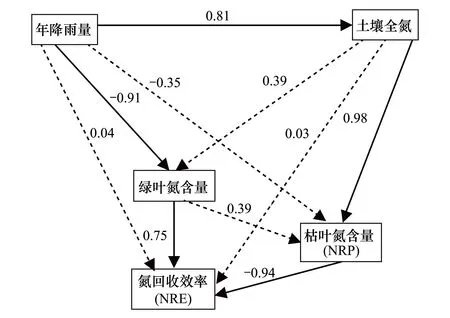
图3 叶片氮回收效率影响因子的结构方程模型分析 Fig.3 The controlling factor analysis of leaf N resorption efficiency using structural equation model 数字代表标准路径系数,其中粗线和粗体数字代表显著性检验P<0.05,虚线代表不显著
3讨论
在羌塘高原从西往东的水分梯度上,本研究验证了随降水量增加土壤全氮及有效氮含量增加,紫花针茅叶氮回收率降低的假设。紫花针茅叶氮回收效率显著高于我国温带草原和世界草地的平均水平,叶片氮回收效率的降低与枯叶养分含量降低密切相关。但与假设相反,生长季盛期绿叶氮含量却随降水量增加而降低。
羌塘高原降水梯度带上土壤TN、土壤无机氮与MAP存在显著正相关关系,说明随着降水增加土壤氮的有效性提高,可能是草地土壤氮矿化能力随降水增加不断增强[17-20,35]导致的。羌塘高原紫花针茅绿叶平均氮含量为(23.87±3.92)g/kg,明显高于中国草地平均水平20.9g/kg[36]及全球平均值20.1g/kg[37],说明低温干旱的高原生态系统紫花针茅具有更高的叶氮水平,是对贫瘠氮限制环境的适应。本研究与Jiang等[2]和Yuan等[6]的结果一致,主要是受低温干旱的影响[1,37-38]。在干旱的条件下植物通过增加叶片内部非光合器官或组织氮的投入,提高细胞内部的渗透压,增强对体内水分的保护作用。同时,低温也会增加叶片氮投入以弥补低效率生理过程。此外,温度升高和降水增加对植被生长的影响可能要大于对土壤氮矿化的影响,从而导致绿叶中氮含量被稀释,叶片氮含量下降。紫花针茅绿叶氮含量与MAP呈显著负相关,干旱端(西部)绿叶中氮含量明显高于湿润端(东部),这与本文的假设刚好相反。羌塘高原紫花针茅绿叶氮含量随降水增加不断降低,原因可能有两方面:一方面,在干旱的条件下植物通过增加叶片内部非光合器官或组织氮的投入,提高细胞内部的渗透压,增强对体内水分的保护作用[39-40],以适应干旱少雨的环境,而当降雨量增加时,水分胁迫逐渐减缓,植物不再需要分配较多的氮到叶片内部非光合器官或组织中,叶片氮含量下降[22];另一方面,降水增加对植被生长的影响可能要大于对土壤氮矿化的影响[20,26],从而导致绿叶中氮含量被稀释,叶片氮含量下降[41]。然而,羌塘高原降水梯度带紫花针茅枯叶氮含量随降水增加无显著差异,因此,可能受叶片氮状况和土壤氮有效性的影响,紫花针茅叶片NRE随降水增加不断降低[6,9,25,41]。通常情况下认为土壤有效氮高的区域植物组织具有更高的氮含量[3,5],然而,羌塘高原气候干旱特征显著,蒸发强度普遍>1800mm,年均风速多在3m/s以上,年平均干燥度指数1.6—20[33],紫花针茅叶片为了增强对体内水分的保护,而增加叶片内部非光合器官或组织氮的投入,提高细胞内部的渗透压[39-40],以适应干旱的环境,从而出现羌塘高原降水少、土壤养分低的一端绿叶氮含量反而高的现象。
羌塘高原紫花针茅叶片具有较高的氮回收能力,平均NRE为(71.25±6.46)%,明显高于李元恒[42]和李玉霖[43]等在内蒙古草原的研究结果,也高于Aerts[1]和Yuan等[6]对全球草地数据的统计结果(分别为58.5%和46.9%)。同时,NRE与MAP呈显著负相关,从东往西随降水减少而不断增加。整个水分梯度带平均的NRP为(6.76±1.42)g/kg,Killingbeck[4]认为当NRP小于7g/kg,叶片几乎达到完全回收水平,因此,羌塘高原降水梯度带紫花针茅叶氮回收效率高是以叶片养分再利用为基础的。紫花针茅叶片极高的NRE和较低的NRP反映了对极端干旱贫瘠环境的适应能力,通过内部氮循环来适应低养分环境[44]。紫花针茅叶片为了维持高的氮投入,干旱端应该具有更高的NRE或更低的NRP,因此,极端干旱的环境可能是造成结果差异的原因。
分析土壤养分对叶片氮回收能力的影响,紫花针茅叶片NRE与土壤TN呈极显著的负相关,特别是与土壤无机氮相关性更强。NRP虽然与土壤TN相关性不显著,但与土壤无机氮呈显著正相关,说明土壤养分会影响植物养分回收能力[9-10],土壤氮的有效性越高,植物叶片NRE越低,叶片氮回收能力和土壤氮有效性显著负相关[14-15]。这也进一步证明生长在养分贫瘠环境中的植被比肥沃环境中的植被具有更高的NRE或更低的NRP[4,7-8]。然而,绿叶氮状况对紫花针茅叶片氮回收能力的影响不明显,这一结果与Aert[1]的研究结论一致,但Kobe等[5]研究结果表明植物绿叶中氮含量越高,相应枯叶中氮含量也高,NRE随绿叶氮含量的增加而降低[16]。Yuan等[3]研究认为NRE与NRP没有显著的相关性,而本研究结果表明NRE与NRP极显著负相关。NRE与绿叶氮含量关系不大,而与枯叶氮含量呈显著负相关,说明枯叶的氮回收水平是影响氮回收效率的内在决定因子。干旱区植物土壤养分有效性低,紫花针茅叶片具有较高的NRE,紫花针茅叶片在衰枯的过程中更多的氮素被重新回收利用,贮藏在根系中[45-46],这一策略可以延长氮素在植物体内驻留的时间。在养分贫瘠的生态系统中较长的氮素驻留时间有利于植物组织养分的维持,可为第2年植物返青创造条件[47]。紫花针茅叶片在衰老的过程中更多的氮素被回收重新利用,导致枯叶中氮素含量更低[48],达到极高回收水平。
羌塘高原水分梯度带内部气温和海拔差异对紫花针茅叶片氮回收能力影响不显著,然而Yuan等[6]通过对全球数据集研究表明NRE随MAT增加不断降低。尽管与其他区域相比,羌塘高原MAT极低,大部分< 0℃,但整个采样梯度带温度差异不显著,最大温差2.4 ℃,因此,羌塘高原MAT对紫花针茅叶片NRE影响不大,同样对NRP的影响也不显著。地形可能影响土壤养分的分布,进而影响植物叶片氮回收能力[49]。然而羌塘高原样带海拔和土壤氮素的分布没有显著的相关性,因此,海拔对采样梯度带紫花针茅叶片NRE和NRP影响不显著。羌塘高原样带位于高原平面上,取样点相对高差不超过300m,地形起伏不大,这是海拔梯度对紫花针茅养分回收能力影响不大的原因。
综上所述,土壤氮的有效性是影响羌塘高原紫花针茅叶片氮回收能力的关键因子,降水通过影响土壤氮的有效性间接影响紫花针茅叶片氮回收效率,同时,枯叶的氮回收水平也对叶片氮回收效率起决定作用。
4结论
羌塘高原降水梯度带温度变化较小,水分是驱动环境梯度的主要关键因子,为研究藏北高原生态过程的生态地理格局提供了一个良好的平台。在羌塘高原降水梯度带上,土壤全氮和有效氮与年降水量呈显著的正相关,湿润端的土壤肥力高于干旱端。受低温干旱环境影响,紫花针茅叶片具有较高的氮含量,且叶氮含量在干旱端高于湿润端;氮回收效率高于全球草地平均水平,与MAP呈显著负相关,枯叶中氮的高效回收利用对叶片氮回收效率起决定作用。土壤氮的有效性是影响羌塘高原紫花针茅叶片氮回收能力的关键因子,降水通过影响土壤氮的有效性以及绿叶中氮含量间接影响紫花针茅叶片氮回收效率。
参考文献(References):
[1]AertsR.Nutrientresorptionfromsenescingleavesofperennials:aretheregeneralpatterns?JournalofEcology, 1996, 84(4): 597- 608.
[2]JiangCM,YuGR,LiYN,GaoGM,YangZP,ShengWP,YuWT.NutrientresorptionofcoexistencespeciesinalpinemeadowoftheQinghai-TibetanPlateauexplainsplantadaptationtonutrient-poorenvironment.EcologicalEngineering, 2012, 44: 1- 9.
[3]YuanZY,LiLH,HanXG,HuangJH,JiangGM,WanSQ.SoilcharacteristicsandnitrogenresorptioninStipa kryloviinativetonorthernChina.PlantandSoil, 2005, 273(1/2): 257- 268.
[4]KillingbeckKT.Nutrientsinsenescedleaves:keystothesearchforpotentialresorptionandresorptionproficiency.Ecology, 1996, 77(6): 1716- 1727.
[5]KobeRK,LepczykCA,IyerM.Resorptionefficiencydecreaseswithincreasinggreenleafnutrientsinaglobaldataset.Ecology, 2005, 86(10): 2780- 2792.
[6]YuanZY,ChenHYH.Global-scalepatternsofnutrientresorptionassociatedwithlatitude,temperatureandprecipitation.GlobalEcologyandBiogeography, 2009, 18(1): 11- 18.
[7]RallhanPK,SinghSP.Dynamicsofnutrientsandleafmassincentralhimalayanforesttreesandshrubs.Ecology, 1987, 68(6): 1974- 1983.
[8]WrightIJ,WestobyM.Nutrientconcentration,resorptionandlifespan:leaftraitsofaustraliansclerophyllspecies.FunctionalEcology, 2003, 17(1): 10- 19.
[9]OleksynJ,ReichPB,ZytkowiakR,KarolewskiP,TjoelkerMG.NutrientconservationincreaseswithlatitudeoforigininEuropean Pinus sylvestrispopulations.Oecologia, 2003, 136(2): 220- 235.
[10]YuanZY,LiLH,HanXG,HuangJH,WanSQ.FoliarnitrogendynamicsandnitrogenresorptionofasandyshrubSalix gordejeviiinnorthernChina.PlantandSoil, 2005, 278(1/2): 183- 193.
[11]AertsR,ChapinIiiFS.Themineralnutritionofwildplantsrevisited:are-evaluationofprocessesandpatterns//FitterAH,RaffaelliDG,eds.SanDiego,CA:AdvancesinEcologicalResearch.AcademicPress, 1999: 1- 67.
[12]CoteB,FylesJW,DjalilvandH.IncreasingNandPresorptionefficiencyandproficiencyinnortherndeciduoushardwoodswithdecreasingfoliarNandPconcentrations.AnnalsofForestScience, 2002, 59(3): 275- 281.
[13]KutbayHG,OkT.FoliarNandPresorptionandnutrientlevelsalonganelevationalgradientinJuniperus oxycedrusL.subsp. macrocarpa (Sibth. &Sm.)Ball.AnnalsofForestScience, 2003, 60(5): 449- 454.
[14]EnokiT,KawaguchiH.NitrogenresorptionfromneedlesofPinus thunbergiiParl.growingalongatopographicgradientofsoilnutrientavailability.EcologicalResearch, 1999, 14(1): 1- 8.
[15]PugnaireFI,ChapinIIIFS.Controlsovernutrientresorptionfromleavesofevergreenmediterraneanspecies.Ecology, 1993, 74(1): 124- 129.
[16]RatnamJ,SankaranM,HananNP,GrantRC,ZambatisN.Nutrientresorptionpatternsofplantfunctionalgroupsinatropicalsavanna:variationandfunctionalsignificance.Oecologia, 2008, 157(1): 141- 151.
[17]DruryCF,ZhangTQ,KayBD.Thenon-limitingandleastlimitingwaterrangesforsoilnitrogenmineralization.SoilScienceSocietyofAmericanJournal, 2003, 67(5): 1388- 1404.
[18]PaulKI,PolglasePJ,O′ConnellAM,CarlyleJC,SmethurstPJ,KhannaPK.Definingtherelationbetweensoilwatercontentandnetnitrogenmineralization.EuropeanJournalofSoilScience, 2003, 54(1): 39- 48.
[19]BurkeIC,LauenrothWK,PartonWJ.Regionalandtemporalvariationinnetprimaryproductionandnitrogenmineralizationingrasslands.Ecology, 1997, 78(5): 1330- 1340.
[20]AustinAT,SalaOE.CarbonandnitrogendynamicsacrossanaturalprecipitationgradientinPatagonia,Argentina.JournalofVegetationScience, 2002, 13(3): 351- 360.
[21]ZhanXY,YuGR,HeNP.Effectsofplantfunctionaltypes,climateandsoilnitrogenonleafnitrogenalongthenorth-southtransectofeasternChina.JournalofResourcesandEcology, 2013, 4(2): 125- 131.
[22]丁小慧, 罗淑政, 刘金巍, 李魁, 刘国华. 呼伦贝尔草地植物群落与土壤化学计量学特征沿经度梯度变化. 生态学报, 2012, 32(11): 3467- 3476.
[23]MeierIC,LeuschnerC.NutrientdynamicsalongaprecipitationgradientinEuropeanbeechforests.Biogeochemistry, 2014, 120(1/3): 51- 69.
[24]ReedSC,TownsendAR,DavidsonEA,ClevelandCC.Stoichiometricpatternsinfoliarnutrientresorptionacrossmultiplescales.NewPhytologist, 2012, 196(1): 173- 180.
[25]OyarzabalM,ParueloJM,delPinoF,OesterheldM,LauenrothWK.TraitdifferencesbetweengrassspeciesalongaclimaticgradientinSouthandNorthAmerica.JournalofVegetationScience, 2008, 19(2): 183- 192.
[26]BarrettJE,McCulleyRL,LaneDR,BurkeIC,LauenrothWK.InfluenceofclimatevariabilityonplantproductionandN-mineralizationinCentralUSgrasslands.JournalofVegetationScience, 2002, 13(3): 383- 394.
[27]BarrettJE,BurkeIC.Potentialnitrogenimmobilizationingrasslandsoilsacrossasoilorganicmattergradient.SoilBiologyandBiochemistry, 2000, 32(11- 12): 1707- 1716.
[28]BarrettJE,BurkeIC.Nitrogenretentioninsemiaridecosystemsacrossasoilorganic-mattergradient.EcologicalApplications, 2002, 12(3): 878- 890.
[29]VintonMA,BurkeIC.Interactionsbetweenindividualplantspeciesandsoilnutrientstatusinshortgrasssteppe.Ecology, 1995, 76(4): 1116- 1133.
[30]MurphyKL,BurkeIC,VintonMA,LauenrothWK,AguiarMR,WedinDA,VirginiaRA,LowePN.RegionalanalysisoflitterqualityinthecentralgrasslandregionofNorthAmerica.JournalofVegetationScience, 2002, 13(3): 395- 402.
[31]李明森. 藏北高原草地资源合理利用. 自然资源学报, 2000, 15(4): 335- 339.
[32]李明森. 羌塘高原土壤特点及其利用. 资源科学, 1980(4): 60- 69.
[33]毛飞, 卢志光, 郑凌云, 张佳华. 近40年那曲地区日照时数和风速变化特征. 气象, 2006, 32(9): 77- 83.
[34]杨富裕, 张蕴薇, 苗彦军, 魏学红. 藏北高寒退化草地植被恢复过程的障碍因子初探. 水土保持通报, 2003, 23(4): 17- 20.
[35]PartonWJ,SchimelDS,ColeCV,OjimaDS.AnalysisoffactorscontrollingsoilorganicmatterlevelsinGreatPlainsGrasslands.SoilScienceSocietyofAmericaJournal, 1987, 51(5): 1173- 1179.
[36]HanWX,FangJY,GuoDL,ZhangY.Leafnitrogenandphosphorusstoichiometryacross753terrestrialplantspeciesinChina.NewPhytologist, 2005, 168(2): 377- 385.
[37]ReichPB,OleksynJ.GlobalpatternsofplantleafNandPinrelationtotemperatureandlatitude.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2004, 101(30): 11001- 11006.
[38]AertsR,CornelissenJHC,vanLogtestijnRSP,CallaghanTV.Climatechangehasonlyaminorimpactonnutrientresorptionparametersinahigh-latitudepeatland.Oecologia, 2007, 151(1): 132- 139.
[39]SeligmanNG,SinclairTR.GlobalenvironmentchangeandsimulatedforagequalityofwheatII.Waterandnitrogenstress.FieldCropsResearch, 1995, 40(1): 29- 37.
[40]OsmondCB,AustinMP,BerryJA,BillingsWD,BoyerJS,DaceyJWH,NobelPS,SmithSD,WinnerWE.StressPhysiologyandtheDistributionofPlants.BioScience, 1987, 37(1): 38- 48.
[41]YuanZY,LiLH.Soilwaterstatusinfluencesplantnitrogenuse:acasestudy.PlantandSoil, 2007, 301(1/2): 303- 313.
[42]李元恒, 韩国栋, 王珍, 王正文, 赵萌莉, 王萨仁娜. 增温和氮素添加降低荒漠草原多年生植物氮素回收效率. 生态学报, 2015, 35(18): 5948- 5956.
[43]李玉霖, 崔夺, 陈静, 毛伟, 赵学勇. 科尔沁沙质草地优势多年生植物氮素回收效率的分异特征. 中国沙漠, 2013, 33(3): 688- 695.
[44]FreschetGT,CornelissenJHC,vanLogtestijnRSP,AertsR.Substantialnutrientresorptionfromleaves,stemsandrootsinasubarcticflora:whatisthelinkwithotherresourceeconomicstraits?NewPhytologist, 2010, 186(4): 879- 889.
[45]MarschnerH.MineralNutritionofHigherPlants.London:AcademicPress, 1995.
[46]GuhaMM,MitchellRL.Thetraceandmajorelementcompositionoftheleavesofsomedeciduoustrees.PlantandSoil, 1966, 24(1): 90- 112.
[47]EcksteinRL,KarlssonPS,WeihM.Leaflifespanandnutrientresorptionasdeterminantsofplantnutrientconservationintemperate-arcticregions.NewPhytologist, 1999, 143(1): 177- 189.
[48]YuanZY,LiLH,HanXG,HuangJH,JiangGM,WanSQ,ZhangWH,ChenQS.Nitrogenresorptionfromsenescingleavesin28plantspeciesinasemi-aridregionofnorthernChina.JournalofAridEnvironments, 2005, 63(1): 191- 202.
[49]DemarsBG,BoernerREJ.FoliarnutrientdynamicsandresorptioninnaturalizedLonicera maackii (Caprifoliaceae)populationsinOhio,USA.AmericanJournalofBotany, 1997, 84(1): 112- 117.
基金项目:国家自然科学基金资助项目(41271067); 中国科学院战略性先导专项课题资助项目(XDA05060700)
收稿日期:2015- 06- 10;
修订日期:2015- 11- 12
*通讯作者
Corresponding author.E-mail: shipl@igsnrr.ac.cn
DOI:10.5846/stxb201506101173
LeafnitrogenresorptionefficiencyofStipa purpureaanditsdeterminantsalongaprecipitationgradientontheChangtangPlateau
ZHAOGuangshuai1,2,XIONGDingpeng1,2,SHIPeili1,*,FENGYunfei1,2,WUJianshuang1,2,ZHANGXianzhou1,ZENGZhaoxu1,2
1 Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China 2 University of Chinese Academy of Sciences, Beijing 100049, China
Abstract:Nitrogen (N) resorption from senescent leaves has long been recognized as an important mechanism by which plants can effectively use soil nutrients and adapt to their environments, particularly under arid and infertile conditions. Many studies have shown that N resorption capacity is controlled by N contents in the soil or vegetation, which is influenced by precipitation. There exists a remarkable precipitation gradient (ranging from less than 100 mm to 700 mm) from the west to east on the Changtang Plateau, where the climate is highly cold and dry and soil nutrients are poor. To better understand the N use characteristics along a precipitation gradient in the alpine nutrient-poor environment, we investigated leaf N resorption strategies of a dominant plant species, Stipa purpurea and their correlations with environmental factors on the Changtang Plateau in Tibet. Green and senescent leaves of S. purpurea and soil samples were collected along the precipitation gradient in July and October 2014. Nitrogen resorption efficiency (NRE, relative reduction in N between green and senescent leaves) and N resorption proficiency (NRP, absolute reduction in N in senescent leaves) were calculated, and the relationships between NRE, NRP, N contents, and climate were determined. Stipa purpurea had higher leaf N concentration and NRE along the precipitation gradient. The average green leaf N concentration of S. purpurea was (23.87 ± 3.92) g/kg in the growing season, which was higher than the mean levels across China (20.9 g/kg) and the world (20.1 g/kg). The leaf N concentration, which was higher in the west than in the east of the plateau, was closely correlated with mean annual precipitation (MAP). The average NRP was (6.76 ± 1.42) g/kg. The mean NRE was (71.25 ± 6.46)%, which was considerably higher than the counterpart both in the Chinese temperate grasslands and in grasslands worldwide (46.9%—58.5%). The N levels of senescent leaves were among the key determinants of NRE. High levels of N resorption from senescent leaves were the internal basis for high NRE. NRE was significantly correlated with MAP, soil total N (TN), and soil inorganic N; NRP was only related with soil inorganic N, but not with TN and MAP. The NRP and NRE were not related to leaf N concentrations, although both were negatively correlated. The influence of mean annual temperature and altitude on NRE and NRP was not significant. Therefore, high NRE and low NRP of S. purpurea might be indicators of N conservation in the arid and nutrient-poor environment. Internal N cycling through N resorption might have reduced N loss. Our findings suggest that soil N availability is a key controlling factor for the N resorption capacity of S. purpurea along the precipitation gradient on the Changtang Plateau, and that precipitation has an indirect effect on NRE by influencing soil N availability and green leaf N concentrations.
Key Words:Changtang Plateau; precipitation gradient; nutrient resorption efficiency; soil nitrogen; leaf nitrogen concentration
赵广帅,熊定鹏,石培礼,冯云飞,武建双,张宪洲,曾朝旭.羌塘高原降水梯度带紫花针茅叶片氮回收特征及影响因素.生态学报,2016,36(11):3419- 3428.
ZhaoGS,XiongDP,ShiPL,FengYF,WuJS,ZhangXZ,ZengZX.LeafnitrogenresorptionefficiencyofStipa purpureaanditsdeterminantsalongaprecipitationgradientontheChangtangPlateau.ActaEcologicaSinica,2016,36(11):3419- 3428.
