氮沉降对杉木和枫香土壤氮磷转化及碳矿化的影响
2016-08-09王方超邹丽群方向民万松泽吴南生王辉民陈伏生
王方超,邹丽群,唐 静,方向民,万松泽,吴南生,*,王辉民,陈伏生
1 江西农业大学林学院, 江西特色林木资源培育与利用2011协同创新中心,南昌 330045 2 赣南师范学院生命与环境科学学院,赣州 341000 3 中国科学院地理科学与资源研究所, 中国科学院千烟洲红壤丘陵综合开发试验站,北京 100101
氮沉降对杉木和枫香土壤氮磷转化及碳矿化的影响
王方超1,邹丽群1,唐静2,方向民1,万松泽1,吴南生1,*,王辉民3,陈伏生1
1 江西农业大学林学院, 江西特色林木资源培育与利用2011协同创新中心,南昌 330045 2 赣南师范学院生命与环境科学学院,赣州 341000 3 中国科学院地理科学与资源研究所, 中国科学院千烟洲红壤丘陵综合开发试验站,北京 100101
摘要:氮沉降是全球变化的重大环境问题,根际是地下生态过程研究的前沿,但目前氮沉降对亚热带地区不同树种土壤氮、磷供应和碳矿化根际过程的影响及其机制尚不清楚。选取典型红壤区15a针叶树杉木(Cunninghamia lanceolata)和阔叶树枫香(Liquidamba formosana)为对象,野外原位开展10 g N m-2a-1氮沉降试验3a,于2014年8月收集杉木和枫香根际土壤和非根际土壤,测定其pH值、有效氮、速效磷、水溶性有机碳及其34 d有机碳矿化动态,并计算根际效应。结果表明:氮沉降显著降低两个树种土壤pH值和杉木根际土壤速效磷(P<0.05);提高枫香非根际土壤-N和杉木非根际土壤水溶性有机碳含量。同时,氮沉降显著提高杉木土壤有机碳矿化速率,根际和非根际的增幅分别为71.2%和41.2%,降低枫香土壤有机碳矿化速率,根际和非根际的降幅分别为10.6%和44.1%。此外,氮沉降显著降低枫香土壤-N和有机碳前期矿化速率的根际效应,增强后期矿化速率的根际效应,而杉木对氮沉降响应不显著。可见,氮沉降可显著改变树木土壤养分供应和有机碳稳定性,且丘陵红壤区针叶树和阔叶树根际过程对氮沉降的响应模式有别。率先报道了亚热带不同树种根际碳、氮、磷耦合过程对氮沉降的响应格局,并较好地揭示了针叶树和阔叶树对氮沉降响应的分异机制。
关键词:根际过程;全球变化;养分供应;有机碳稳定性;丘陵红壤
氮(N)沉降的持续加重及其生态效应受到科学家的广泛关注[1-2]。20世纪以来,由于人类活动的影响,N沉降量不断升高,全球当前N沉降速率已超过10 kg N hm-2a-1,预计到2050年还将增加1倍,部分地区可达50 kg N hm-2a-1[3]。随着社会经济的快速发展,中国已是继欧洲和美国之后的第三大N沉降区[4]。已有研究表明,N沉降影响生态系统的养分循环、碳(C)动态、植被动态,凋落物分解和土壤酸化等各个方面[2,5],但这些影响仍存在很大的不确定性。

根际被认为是森林生物地球化学循环的关键区域,是物质和能量转化最激烈的区域之一,范围一般为距根表几毫米至数厘米的区域[15]。N沉降对养分转化和有机C矿化的影响效应可能优先作用于根际土壤,因为植物根产生分泌物和脱落物可为根际区土壤微生物提供有效C和N源,使根际土壤微生物在数量上显著高于非根际土壤,并随之引起一系列土壤生化过程的变化与连锁效应[16]。此外,由于不同物种对N的需求各异,不同树种根际分泌物对N沉降的响应不同,这必然影响根际土壤N转化和有机C矿化。总体来看,N沉降对树木根际过程及其效应的研究报道较少,对不同树种(如:针叶树和阔叶树)的响应过程及其机制的了解还明显不够[17],不利于确切提出森林应对全球N沉降的有效措施。
杉木(Cunninghamialanceolata)是我国南方主要用材树种,为亚热带常绿针叶林的代表性树种[18]。枫香(Liquidambaformosana)为长江中下游次生林的重要冠层树种,是亚热带落叶阔叶树的代表[19]。基于此,结合南方丘陵山区N沉降不断加重的生态问题,研究两种代表性树种根际N、P养分转化和有机C矿化对N沉降的响应过程及其异同机制。研究结果有助于更好的剖析和理解针叶树和阔叶树地下根际N、P和C过程的耦合效应与调控机制,为全球变化背景下亚热带地区森林经营和管理提供科学依据。
1材料与方法
1.1研究区概况
研究地点位于江西省泰和县灌溪镇中国科学院千烟洲试验站(115°04′13″E,26°44′48″N,海拔多在100 m左右,相对高度差20—50 m),该地为典型红壤丘陵地貌,地势起伏、丘顶浑圆,以10°—25°坡度居多,由白垩系红色与紫红色砾岩、砂岩和泥岩组成,除少数丘顶和裸露地外,风化层一般厚30—50 cm。林地土壤为红黄壤,年均气温18℃,年平均降水量约1500 mm,年均相对湿度90%,年日照时数1400 h,全年降水充沛,但年际变化较大,年内分布不均匀,属典型亚热带季风气候[20]。
1.2试验设计
试验样地为典型红壤区15a杉木和枫香混交林(表1),其中杉木和枫香的比例为8∶2,在8块20 m ×20 m混交林地中野外原位开展10 g N m-2a-1N沉降模拟试验,其中N沉降处理和对照各4块,配对设置,施肥自2012年春起,每年4季各1次,施肥方式为NH4NO3拌少量细沙均匀撒施,对照只撒等量细沙。试验处理之前,在每块样地用3 cm土钻随机取表层(0—20 cm)土壤36个点混合,预处理后,测定其基本养分特性(表2)。
表1氮沉降处理前杉木和枫香混交林林分基本背景特征
Table 1General stand properties in mixed forests ofCunninghamialanceolataandLiquidambaformosanabefore nitrogen deposition treatment
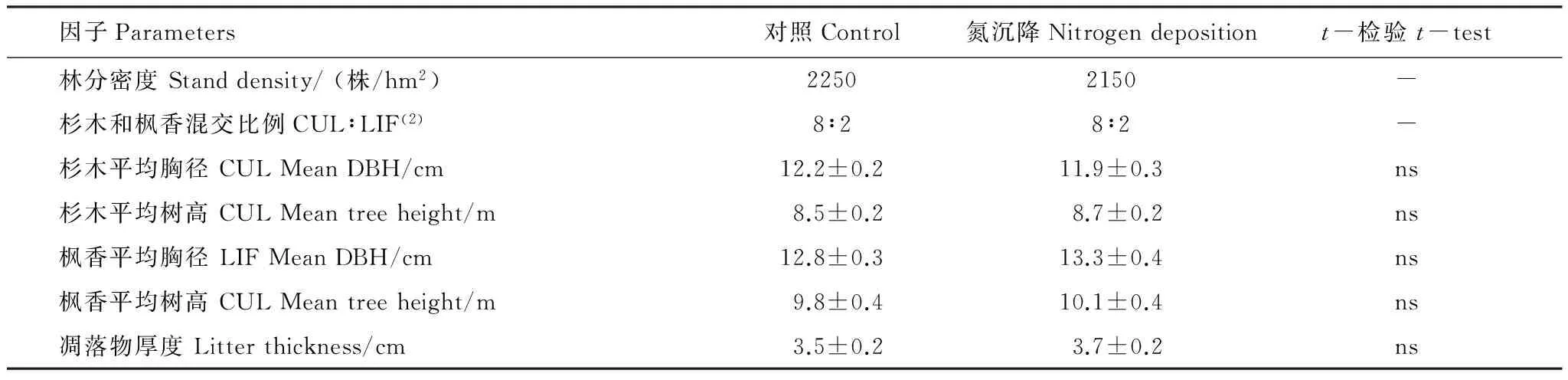
因子Parameters对照Control氮沉降Nitrogendepositiont-检验t-test林分密度Standdensity/(株/hm2)22502150-杉木和枫香混交比例CUL∶LIF(2)8∶28∶2-杉木平均胸径CULMeanDBH/cm12.2±0.211.9±0.3ns杉木平均树高CULMeantreeheight/m8.5±0.28.7±0.2ns枫香平均胸径LIFMeanDBH/cm12.8±0.313.3±0.4ns枫香平均树高CULMeantreeheight/m9.8±0.410.1±0.4ns凋落物厚度Litterthickness/cm3.5±0.23.7±0.2ns
平均值±标准误,ns,差异不显著; CUL:杉木Cunninghamialanceolata;LIF:枫香Liquidambaformosana
1.3样品采集
2014年8月,在每块样地中选定杉木和枫香标准木各1株,4次重复,共16株。在离树干0.5—1 m的位置挖掘3个10 cm×10 cm×15 cm的土块,将依附在植物根系上4 mm之内土壤轻轻抖落出来,并用镊子将细根及其它杂物移除,将抖落下来的土壤定义为根际土;非根际土为未依附于根系上的土壤[10,21]。收集好足量土壤后,将其混匀分装过2 mm筛,在4℃冰箱中保存,并尽快完成土壤相关指标的测定。
1.4土壤养分特性的测定

1.5土壤碳矿化估测方法
土壤有机C矿化的估测采用碱液密闭吸收法,每个样品各称取3份过2 mm网筛的新鲜土壤30 g于可封闭的广口瓶中,加蒸馏水至饱和持水量的40%左右,量取1 mol/L NaOH溶液10 mL于50 mL烧杯中,在广口瓶内盖上涂抹凡士林直至密封良好,密闭放入25℃的恒温箱中进行培养,采用碱液吸收法测定矿化量,测定时间为第3、6、9、14、19、24天和第34天[23]。为便于描述和分析,本文将0—9 d、9—19 d和19—34 d划分为前期、中期和后期3个矿化阶段。
有机C矿化的动力学模型参数计算公式使用Cm=C0(1-e-kt),Cm和C0分别代表在培养期累积的OC矿化量(%)和矿化潜力(矿化势%),k和t分别代表矿化常数和培养时间。同时,将培养34 d C矿化总量占水溶性有机C量之比定义为可矿化强度;将N沉降处理有机C矿化速率与对照之比定义为对N沉降的响应强度。
1.6根际效应的界定
根际效应是指根际土壤理化与生物学特性不同于非根际土壤的差异性。通常用R/B来表示,其中R代表根际土壤,B代表非根际土壤,根际正效应是指根际土壤中某个指标的量高于非根际土壤,因此,用R/B>1表示为正效应,R/B<1为负效应[10]。
1.7数据处理
采用SPSS 17.0 统计软件和Excel 2007进行统计和数据分析,不同处理间参数的差异采用Duncan法进行多重比较(P<0.05)。
2结果与分析
2.1氮沉降对土壤氮磷供应的影响
表2氮沉降处理前杉木和枫香混交林土壤基本养分特征
Table 2Soil general nutrient properties in the mixed forest ofCunninghamialanceolataandLiquidambaformosanabefore nitrogen deposition treatment
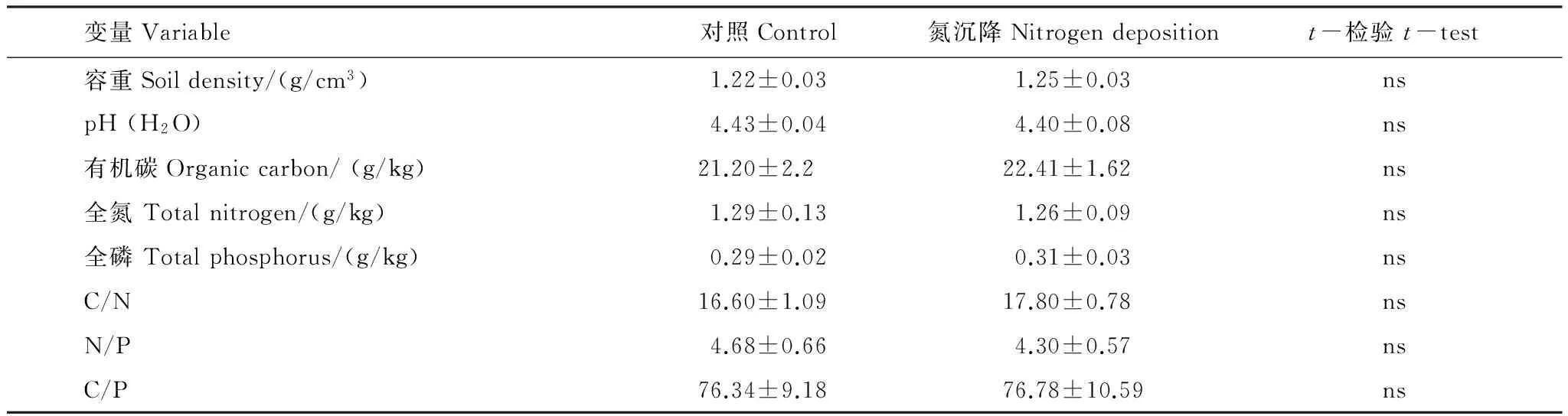
变量Variable对照Control氮沉降Nitrogendepositiont-检验t-test容重Soildensity/(g/cm3)1.22±0.031.25±0.03nspH(H2O)4.43±0.044.40±0.08ns有机碳Organiccarbon/(g/kg)21.20±2.222.41±1.62ns全氮Totalnitrogen/(g/kg)1.29±0.131.26±0.09ns全磷Totalphosphorus/(g/kg)0.29±0.020.31±0.03nsC/N16.60±1.0917.80±0.78nsN/P4.68±0.664.30±0.57nsC/P76.34±9.1876.78±10.59ns
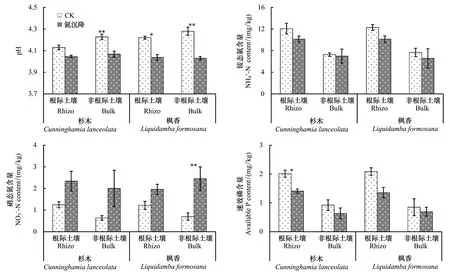
图1 两种处理条件下杉木和枫香根际和非根际土壤pH值和有效养分的变异格局Fig.1 Rhizosphere and bulk soil pH and available nutrients of two trees with and without N deposition平均值±标准误;* P<0.05,** P<0.01,** P<0.001
2.2氮沉降对不同树种土壤有机碳矿化的影响
方差分析结果表明,N沉降对根际和非根际土壤矿化势影响显著,对非根际水溶性有机C、可矿化强度和前期矿化速率影响显著;对中期、后期矿化速率和平均矿化速率影响均不显著;除矿化势以外,不同树种以上土壤化学指标在根际土壤中均无显著差异。N沉降处理和不同树种对根际和非根际土壤可矿化强度、前期、中期和平均矿化速率的交互作用显著,仅对非根际土壤后期矿化速率和矿化势交互作用显著。此外,N沉降仅对前期矿化速率的根际效效应影响显著,N沉降处理和不同树种的交互作用仅对后期矿化速率具显著影响(图2)。
进一步对比分析发现,N沉降仅显著提高了杉木非根际土壤水溶性有机C含量,增幅为23.6%,对杉木根际土壤、枫香根际和非根际土壤水溶性有机C含量影响不显著。N沉降显著降低枫香非根际土壤有机C可矿化强度,降幅达44.2%,但对杉木根际和非根际、枫香根际土壤有机C可矿化强度影响不显著(图2)。
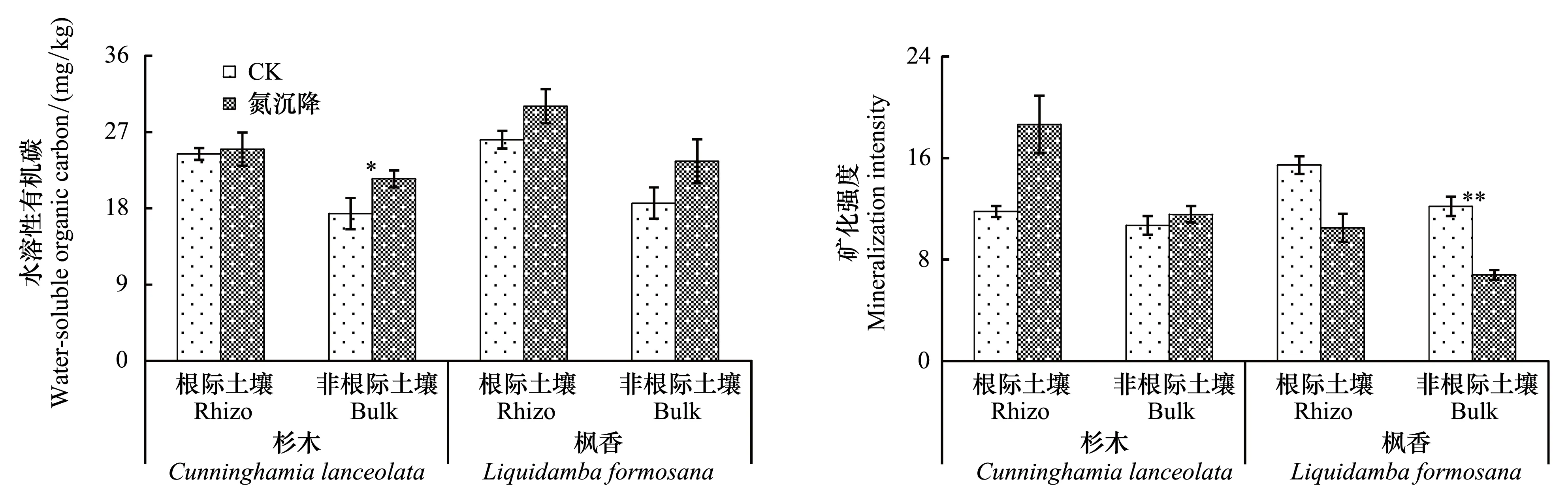
图2 不同处理杉木和枫香根际和非根际土壤水溶性有机碳和可矿化强度(可矿化碳/水溶性有机碳)的变异格局Fig.2 Rhizosphere and bulk soil water soluble organic carbon and mineralization intensity of two trees with and without N deposition
杉木根际和非根际土壤的34 d有机C矿化量均表现为N沉降处理高于对照,增幅分别为71.2%和41.2%;枫香根际和非根际土壤的34 d矿化量均表现为对照高于N沉降处理,降幅分别为10.6%和44.1%;其中对照处理枫香非根际土壤矿化量最高((67.13±5.09)mg CO2/g OC),对照处理杉木根际土壤矿化量最低((33.49±3.01)mg CO2/g OC)(图3)。同时,杉木土壤有机C各个时期的矿化速率对N沉降的响应强度均表现为正效应,而枫香则均为负效应(图4)。
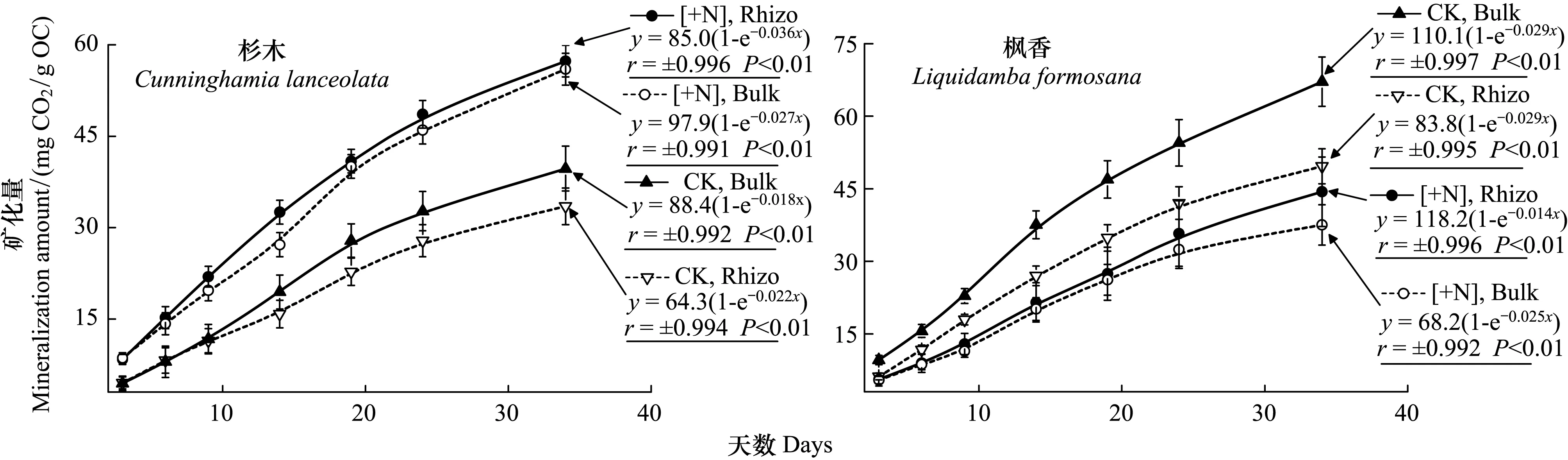
图3 不同处理杉木和枫香根际和非根际土壤有机碳矿化动态及其模型Fig.3 The dynamics and their models of rhizosphere and bulk soil organic carbon mineralization under two tree species with and without nitrogen depositionCK, Rhizo表示对照处理根际土壤;CK, Bulk表示对照处理非根际土壤;[+N], Rhizo表示N沉降处理根际土壤;[+N], Bulk表示N沉降处理非根际土壤
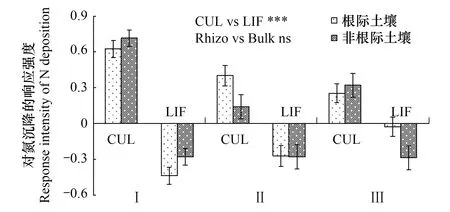
图4 杉木和枫香根际和非根际土壤有机碳矿化前期(Ⅰ)、中期(Ⅱ)、后期(Ⅲ)对N沉降的响应强度(N沉降/对照)Fig.4 Response intensity to nitrogen deposition at early (Ⅰ), middle (Ⅱ), late (Ⅲ) organic carbon mineralization rates in rhizosphere and bulk soil under two tree speciesⅠ,前期Early stage;Ⅱ,中期Middle stage;Ⅲ,后期Late stage;CUL,杉木 Cunninghamia lanceolata; LIF,枫香 Liquidamba formosana
2.3两个树种各变量根际效应对氮沉降的响应格局
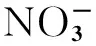
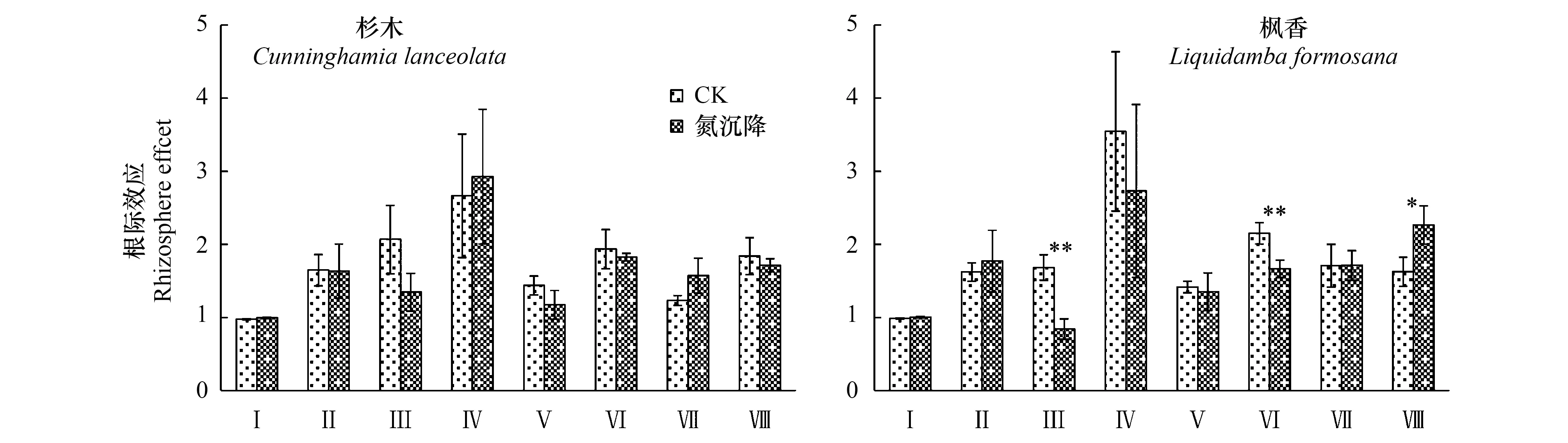
图5 不同处理条件下杉木和枫香土壤氮磷供应和有机碳稳定性的根际效应Fig.5 The rhizosphere effects of soil nitrogen and phosphorus supply, and organic carbon stability under two tree species with and without nitrogen depositionI表示pH值的根际效应;II、III和IV分别表示-N和速效磷的根际效应根际效;V表示水溶性有机碳的根际效应;VI、VII和VIII分别表示有机碳前期矿化速率、中期矿化速率和后期矿化速率的根际效应
3讨论
氮沉降对土壤生态过程的影响已成为近年来森林生态学研究的热点[2,4,7]。N沉降影响效应的多面性表现为,在缺N的某些地区,适量N沉降对生态系统将产生正效应,如增加土壤N供应,从而提高生态系统生产力;但在大多数地区,过量N沉降给养分循环带来负面效应,如过量N导致土壤C∶N∶P化学计量比改变,影响土壤呼吸,降低土壤pH,加快养分流失,形成N饱和症状等[14, 24-25]。根际是陆地生态系统物质和能量转化最激烈的区域[10, 15],N沉降对于地下生态过程的影响可能优先作用于根际土壤[17]。
碳和N的耦合是森林生态过程研究关注的焦点之一[2]。氮沉降改变森林土壤N循环,必将影响土壤C过程。本研究发现,N沉降提高杉木非根际土壤水溶性有机C含量,增强杉木根际和非根际土壤有机C矿化速率,而降低枫香非根际土壤有机C可矿化强度,抑制枫香根际和非根际土壤有机C矿化速率。袁颖红等[31]于杉木林中开展模拟N沉降试验,发现随着N沉降量增加,土壤可溶性有机C增加。Deforest等[32]在美国密歇根开展的N沉降试验结果也表明,土壤可溶性有机C含量因N沉降可导致可溶性酚类物质积累而显著提高。与此同时,胡正华等[33]在北亚热带落叶阔叶林开展了模拟N沉降试验,发现N沉降显著抑制土壤呼吸。刘博奇等[14]在北方针叶林内开展模拟N沉降试验,发现N沉降提高了土壤呼吸速率。李凯等[34]开展模拟N沉降对石栎(Lithocarpusglaber)和苦槠(Castanopsissclerophylla)幼苗土壤呼吸速率影响的研究,发现土壤呼吸对N沉降的响应因施N水平和幼苗种类不同而各异,但两树种的土壤呼吸在中、高N处理下均表现为抑制。我们的研究结果也表明,针叶树杉木和阔叶树枫香的土壤C过程对N沉降的响应各异,潜在原因可能是:杉木为喜肥性的速生树种,且根系无内外生菌根[18],N输入可促进根系的生长发育,提高根系的活性,导致根系分泌物增加,提高可溶性C含量及其矿化速率;而枫香为具内外菌根菌的树种[19],高N输入不利于菌根菌的生存,抑制根系发育及碳水化合物的分泌,导致土壤有机C矿化强度和矿化速率下降。为此,也不难解释杉木矿化速率对N沉降的响应表现为正效应,即根际有机C矿化速率优先加快;而枫香表现为负效应,即其根际有机C矿化速率优先受到抑制。
综上所述,许多他人的研究和本研究都表明N沉降效应因树种不同而表现各异,本研究还发现根际和非根际土壤养分转化和C稳定性对N沉降的响应也存在显著差异。无菌根的喜肥针叶树种杉木根际和非根际土壤过程对N沉降响应的同步性显著强于具有菌根的阔叶树种枫香。鉴于N沉降效应的复杂性,今后的研究应更多地关注土壤-植物-微生物的耦合机制[15],并力求将地下和地上过程联系起来[16],为森林应对全球N沉降提供有益参考。
参考文献(References):
[1]Wei C, Yu Q, Bai E, Lü X, Li Q, Xia J, Kardol P, Liang W, Wang Z, Han X. Nitrogen deposition weakens plant-microbe interactions in grassland ecosystems. Global Change Biology, 2013, 19(12): 3688- 3697.
[2]Chen F S, Duncan D S, Hu X F, Liang C. Exogenous nutrient manipulations alter endogenous extractability of carbohydrates in decomposing foliar litters under a typical mixed forest of subtropics. Geoderma, 2014, 214- 215: 19- 24.
[3]Galloway J N, Dentener F J, Capone D G, Boyer E W, Howarth R W, Seitzinger S P, Asner G P, Cleveland C C, Green P A, Holland E A, Karl D M, Michaels A F, Porter J H, Townsend A R, Vöosmarty C J. Nitrogen cycles: past, present, and future. Biogeochemistry, 2004, 70(2): 153- 226.
[4]王晶苑, 张心昱, 温学发, 王绍强, 王辉民. 氮沉降对森林土壤有机质和凋落物分解的影响及其微生物学机制. 生态学报, 2013, 33(5): 1337- 1346.
[5]陈浩, 莫江明, 张炜, 鲁显楷, 黄娟. 氮沉降对森林生态系统碳吸存的影响. 生态学报, 2012, 32(21): 6864- 6879.
[6]詹书侠, 陈伏生, 胡小飞, 甘露, 朱友林. 中亚热带丘陵红壤区森林演替典型阶段土壤氮磷有效性. 生态学报, 2009, 29(9): 4673- 4680.
[7]Gundersen P, Emmett B A, Kjønaas O J, Koopmans C J, Tietema A. Impact of nitrogen deposition on nitrogen cycling in forests: a synthesis of NITREX data. Forest Ecology and Management, 1998, 101(1/3): 37- 55.
[8]樊后保, 刘文飞, 李燕燕, 廖迎春, 袁颖红, 徐雷. 亚热带杉木(Cunninghamialanceolata)人工林生长与土壤养分对氮沉降的响应. 生态学报, 2007, 27(11): 4630- 4642.
[9]庞丽, 张一, 周志春, 丰忠平, 储德裕. 模拟氮沉降对低磷胁迫下马尾松不同家系根系分泌和磷效率的影响. 植物生态学报, 2014, 38(1): 27- 35.
[10]刘煜, 胡小飞, 陈伏生, 袁平成. 马尾松和苦槠林根际土壤矿化和根系分解CO2释放的温度敏感性. 应用生态学报, 2013, 24(6): 1501- 1508.
[11]Lorenz K, Lal R. Biogeochemical C and N cycles in urban soils. Environment International, 2009, 35(1): 1- 8.
[12]张炜, 莫江明, 方运霆, 鲁显楷, 王晖. 氮沉降对森林土壤主要温室气体通量的影响. 生态学报, 2008, 28(5): 2309- 2319.
[13]Izauralde R C, Megill W B, Rosenberg N J. Carbon cost of applying nitrogen fertilizer. Science, 2000, 288(5467): 811- 812.
[14]刘博奇, 牟长城, 邢亚娟, 王庆贵. 模拟氮沉降对云冷杉红松林土壤呼吸的影响. 林业科学研究, 2012, 25(6): 767- 772.
[15]Philippot L, Raaijmakers J M, Lemanceau P, van der Putten W H. Going back to the roots: the microbial ecology of the rhizosphere. Nature Reviews Microbiology, 2013, 11(11): 789- 799.
[16]Terzano R, Cesco S, Mimmo T. Dynamics, thermodynamics and kinetics of exudates: crucial issues in understanding rhizosphere processes. Plant and Soil, 2015, 386(1/2): 399- 406.
[17]Phillips R P, Fahey T J. The influence of soil fertility on rhizosphere effects in northern hardwood forest soils. Soil Science Society of America Journal, 2008, 72(2): 453- 461.
[18]盛炜彤, 童书振, 段爱国. 杉木丰产栽培实用技术. 北京: 中国林业出版社, 2011.
[19]苏琍英, 程爱兴, 喻爱林, 傅卫庆, 郑平谣. 天目山自然保护区林木菌根调查. 浙江林学院学报, 1992, 9(3): 263- 276.
[20]陈永瑞, 林耀明, 李家永, 刘允芬, 杨汝荣. 江西千烟洲试区杉木人工林降雨过程及养分动态研究. 中国生态农业学报, 2004, 12(1): 74- 76.
[21]Phillips R P, Fahey T J. Tree species and mycorrhizal associations influence the magnitude of rhizosphere effects. Ecology, 2006, 87(5): 1302- 1313.
[22]鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000: 156- 183.
[23]陈伏生. 城乡梯度森林生态过程研究. 北京: 中国林业出版社, 2013: 72- 93.
[24]Turner M M, Henry H A L. Interactive effects of warming and increased nitrogen deposition on15N tracer retention in a temperate old field: seasonal trends. Global Change Biology, 2009, 15(12): 2885- 2893.
[25]沈芳芳, 袁颖红, 樊后保, 刘文飞, 刘苑秋. 氮沉降对杉木人工林土壤有机碳矿化和土壤酶活性的影响. 生态学报, 2012, 32(2): 517- 527.
[26]Bergkvist B, Folkeson L. Soil acidification and element fluxes of aFagussylvaticaforest as influenced by simulated nitrogen deposition. Water, Air, and Soil Pollution, 1992, 65(1/2): 111- 133.
[27]袁颖红, 樊后保, 王强, 裘秀群, 陈秋凤, 李燕燕, 黄玉梓, 廖迎春. 模拟氮沉降对杉木人工林土壤有效养分的影响. 浙江林学院学报, 2007, 24(4): 437- 444.
[28]孟范平, 李桂芳. 酸雨对土壤元素化学行为的影响. 中南林学院学报, 1998, 18(1): 27- 34.
[29]鲁如坤, 时正元, 钱承梁. 磷在土壤中有效性的衰减. 土壤学报, 2000, 37(3): 323- 329.
[30]陈希, 陈伏生, 叶素琼, 喻苏琴, 方向民, 胡小飞. 丘陵红壤茶园根际氮磷转化对不同强度酸雨胁迫的响应. 应用生态学报, 2015, 26(1): 1- 8.
[31]袁颖红, 樊后保, 刘文飞, 张子文, 孟庆银, 胡锋, 李辉信. 模拟氮沉降对杉木人工林土壤可溶性有机碳和微生物量碳的影响. 水土保持学报, 2012, 26(2): 138- 143.
[32]DeForesta J L, Zaka D R, Pregitzerc K S, Burtonc A J. Atmospheric nitrate deposition and the microbial degradation of cellobiose and vanillin in a northern hardwood forest. Soil Biology and Biochemistry, 2004, 36(6): 965- 971.
[33]胡正华, 李涵茂, 杨燕萍, 陈书涛, 李岑子, 申双和. 模拟氮沉降对北亚热带落叶阔叶林土壤呼吸的影响. 环境科学, 2010, 31(8): 1726- 1732.
[34]李凯, 江洪, 由美娜, 曾波. 模拟氮沉降对石栎和苦槠幼苗土壤呼吸的影响. 生态学报, 2011, 31(1): 82- 89.
基金项目:国家自然科学基金资助项目(31260199, 31360179);江西省研究生创新资金资助项目(YC2014-S190);江西省青年科学家培养对象资助项目(20122BCB23005);江西省自然科学基金资助项目(2012BAB204005)
收稿日期:2015- 06- 17;
修订日期:2015- 10- 22
*通讯作者
Corresponding author.E-mail: rensh111@126.com
DOI:10.5846/stxb201506171222
Influence of nitrogen deposition on soil nutrient supply and organic carbon mineralization inCunninghamialanceolataandLiquidambarformosanaplantations
WANG Fangchao1, ZOU Liqun1, TANG Jing2, FANG Xiangmin1, WAN Songze1, WU Nansheng1,*, WANG Huimin3, CHEN Fusheng1
1CollegeofForestry,JiangxiAgriculturalUniversity,Nanchang330045,China2CollegeofLifeandEnvironmentSciences,GannanNormalUniversity,Ganzhou341000,China3InstituteofGeographicScienceandNaturalResearch,ChineseAcademyofSciences,Beijing100101,China
Abstract:Nitrogen (N) deposition is a major environmental issue that affects global climate change. Study of the rhizosphere has become a research frontier in underground ecology. However, the effects of N deposition on N and phosphorus (P) supplies, and organic carbon (OC) mineralization in the rhizosphere of trees in subtropical China remains unclear, and whether the underlying mechanisms differ between tree species need to be determined. In the present study, a coniferous tree (Cunninghamia lanceolata) and a broadleaf tree (Liquidambar formosana) each 15 years of age were selected at the Qianyanzhou Experimental Station of the Chinese Academy of Sciences to evaluate in situ N deposition. Trees were treated with 10 g N m-2a-1and compared to the control. After 3 years, rhizosphere and bulk soils for both tree species were collected to assess pH value, mineral N, available P, water-soluble organic carbon (WSOC), and the dynamic of OC mineralization following 34 days incubation. Rhizosphere effects of all measured variables were assessed using ratios of rhizosphere to bulk soil, and OC mineralization parameters at early (0—9 d), middle (9—19 d), and late stages (19—34 d) were obtained using the traditional dynamics model. N deposition significantly decreased rhizosphere and bulk soil pH, available P in rhizosphere soil of C. lanceolata, and OC mineralization intensity in bulk soil of L. formosana. N deposition also increased -N in bulk soil of L. formosana and WSOC in bulk soil of C. lanceolata (P < 0.05). In contrast, -N, mineral N, and the ratio of mineral N to available P in rhizosphere and bulk soils did not differ significantly between both species. N deposition significantly increased OC mineralization rates in C. lanceolata rhizosphere and bulk soil by 71.2% and 41.2%, respectively; but decreased OC mineralization rates in L. formosana rhizosphere and bulk soil by 10.6% and 44.1%, respectively. N deposition significantly decreased both the rhizosphere effects of -N and the early OC mineralization rate in L. formosana, but increased its late OC mineralization rate; whereas the rhizosphere effects of all measured variables in C. lanceolata showed no significant differences between control (CK) and N treatments. N deposition significantly altered soil nutrient supply and OC stability, and the synchronicity of responses to N deposition between rhizosphere and bulk soil was stronger for C. lanceolata than for L. formosana. We concluded that response of rhizosphere processes to N deposition differs between conifer and broadleaf tree species. The response of rhizosphere OC mineralization to N deposition showed a positive effect for C. lanceolata, and a negative effect for L. formosana, as their rates preferentially increased and decreased, respectively with N deposition. This difference could be attributed to their nutrient requirements and root traits, because C. lanceolata prefers fertile soil and are not associated with mycorrhizal fungi, whereas L. formosana can tolerate infertile soil with roots that are associated with mycorrhizal fungi. To our knowledge, the present study is the first to report on the effects of N deposition on the interactions between supplies of N and P in soil, and the stability of OC in different tree species of subtropical China, and reports divergent mechanisms in coniferous versus broadleaf tree species.
Key Words:rhizosphere process; global climate change; nutrient supply; organic carbon stability; hilly red soil
王方超,邹丽群,唐静,方向民,万松泽,吴南生,王辉民,陈伏生.氮沉降对杉木和枫香土壤氮磷转化及碳矿化的影响.生态学报,2016,36(11):3226- 3234.
Wang F C, Zou L Q, Tang J, Fang X M, Wan S Z, Wu N S, Wang H M, Chen F S.Influence of nitrogen deposition on soil nutrient supply and organic carbon mineralization inCunninghamialanceolataandLiquidambarformosanaplantations.Acta Ecologica Sinica,2016,36(11):3226- 3234.
