牛蒡甙元在人类风湿性关节炎成纤维样滑膜细胞生长与凋亡中的影响
2016-08-06陈烁周利武张雷赵云龙杨超赵建宁
陈烁 周利武 张雷 赵云龙 杨超 赵建宁*
1. 南京军区南京总医院骨科, 江苏 南京 210002 2. 南京大学医学院临床学院骨科, 江苏 南京 210000
类风湿性关节炎(RA)是一种以滑膜组织假瘤样生长为主要特征的慢性炎症性疾病[1]。RA滑膜组织的主要特征之一是成纤维样滑膜细胞呈现肿瘤样的生长方式。RAFLS能够抑制凋亡并以非依赖性方式增殖,这种生长方式与细胞在无细胞外基质环境中的生存和凋亡能力相关[2]。RAFLS过度增生导致关节软骨的破坏,虽然RAFLS增生的机制尚未完全明确,但参与此过程的一些抗凋亡分子或信号通路已经得到确认[3]。RAFLS表达的抗凋亡成分包括细胞型Fas相关死亡结构域蛋白样白介素-1β转换酶抑制蛋白(FLIP)[4]、P53基因突变[5], 泛素相关蛋白(sentrin)[6],类泛素修饰因子1(SUMO-1)[6],Bcl-2[7],NF-κB和/或Akt信号通路的激活[8-9]。
牛蒡是一种能够有效减轻风湿病疼痛和发热的中药材。牛蒡甙元(C21H24O6;分子量:372.41)是一种从牛蒡中提取的苯丙素二苄基丁内酯木酚素,具有抗炎、抗氧化、抗癌、抗病毒等多种生物活性[10-15]。作为一种新抗癌剂,牛蒡甙元通过MOS/P38 MAPKs通路诱导非嗜荷尔蒙类型乳癌细胞凋亡[16]。对于膀胱癌T24细胞,牛蒡甙元能诱导细胞周期停滞及细胞凋亡[17]。牛蒡甙元通过下调生存蛋白的表达、抑制信号传感器和STAT3信号通路激活剂,来增强癌细胞对于顺铂的化学敏感性[18-19]。本研究旨在探索牛蒡甙元对RAFLS增殖和凋亡的影响。
1 材料和方法
1.1 细胞分离和处理
22例接受膝关节置换手术的类风湿性关节炎患者,其中男性6名,女性16名,年龄39-63岁,在手术获得的关节滑膜组织中提取RAFLS。全部患者均符合美国风湿病学会关于类风湿性关节炎的诊断标准[20],研究开始前已获得患者的书面同意。将滑膜组织切成2~3 mm的碎片,加入4 mg/ml的I型胶原酶(Worthington, Freehold, NJ, US),在DMEM(Invitrogen Life Tech, Carlsbad, CA, US)中放置,在含5%CO2的37℃保温箱中保存4h。经酶处理后,成纤维样滑膜细胞用尼龙细胞滤网(BD Falcon,Franklin Lakes,NJ, US)滤出,DMEM充分洗涤后浸泡在含10%v/v牛胎儿血清(FBS; Invitrogen)、1%青霉素-链霉素溶液(Invitrogen)、1%谷氨酰胺(Sigma, St. Louis, MO, US)的DMEM中培养。细胞保存于含5%CO2的37℃保温箱中,3天更换1次培养液。当80%~90%细胞汇集以后,贴壁细胞就会被胰蛋白酶化,并以1∶3的比例分裂。本实验使用的FLS在反式显微镜下有结构上的均质性,具有典型的成纤维样结构。FLS的纯度用流式细胞仪进行测定,检测异硫氰酸荧光素(FITC)标记的抗CD3,PE标记的抗CD90和APC标记的抗CD14单克隆抗体含量(由BD Pharmingen公司提供)。RAFLS传代3~8次后,冻存于10%二甲基亚砜/90%胎牛血清并储存在液氮中。
细胞接种于96孔板或60 mm培养皿(Corning Life Sciences, Acton, MA),于无血清DMEM培养基培养24 h,然后分别在无牛蒡甙元和含有10、20、30 μM牛蒡甙元的DMEM全培养基中培养。
1.2 MTT实验
RAFLS在浓度为每孔1×104的96孔细胞培养板中培养,一式三份。三分标本分别培养并观察12、24、48 h,在每个时间点结束前3 h,每孔加入20 μl噻唑蓝溶液(5 mg/ml磷酸盐缓冲液),并且在37℃环境下培养3 h,移除媒介后每孔加入100 μl二甲基亚砜,细胞培养板在定轨摇床中轻柔旋转10 min,使沉淀完全溶解,再用酶标分析仪测量570 nm处吸光度值。
1.3 乳酸脱氢酶检测分析
RAFLS在浓度为每孔1×104的96孔细胞培养板中培养,一式三份。三分标本分别培养并观察48 h,以LDH在媒介中的释放量来评估牛蒡甙元的细胞毒性。
1.4 细胞凋亡实验
RAFLS在浓度为每孔5×104的24孔细胞培养板中培养,一式三份。三分标本分别培养并观察48 h。经PBS洗涤后,用膜联蛋白V-FITC凋亡检测工具进行染色,然后用FACSCaliburTM流式细胞仪检测和CELLQuest软件进行分析,碘化丙啶阴性而膜联蛋白V阳性的为凋亡细胞,二者皆阳性的为坏死细胞。
1.5 线粒体膜电位的测定
RAFLS在37℃用浓度1 μg/ml的荧光探针JC-1染色处理10 min,洗涤后用流式细胞仪分析。对JC-1单体,其荧光计被设置为490 nm激发波长和390 nm发射波长。对JC-1总体,荧光计被设置为525 nm激发波长和590 nm发射波长。用FACSCaliburTM流式细胞仪测量荧光效应。在健康细胞,JC-1作为一个单体存在于胞质中,也作为聚合物在线粒体中积累。在凋亡细胞,C-1只在单体形式存在并产生一个绿色的胞质信号。
1.6 Western blot
将2×105细胞接种于60 mm培养皿中经上述处理后收集细胞,分别通过ReadyPrep血清总蛋白提取工具和胞浆、核蛋白提取工具,提取血清总蛋白、胞浆蛋白、核内蛋白。通过染色法确定蛋白质浓度,蛋白通过10%十二烷基硫酸钠聚丙烯酰胺凝胶电泳分离,然后转移到硝酸纤维素膜,在室温下用5%脱脂牛奶在三羟甲基氨基甲烷缓冲盐水和Tween 20 (10 mmol/l Tris-Cl [pH 8.0], 150 mmol/l NaCl, 0.05% Tween 20)中阻断硝酸纤维素膜1 h,然后在4℃下过夜后孵化出主要抗体,如Bcl-2抗体、Bax抗体、cytochrome c抗体、cleaved caspase-9抗体、cleaved caspase-3抗体、p65抗体、IκBα抗体、Akt抗体、pAkt抗体、β-actin抗体,用TBST洗涤后,在室温下与二抗连同辣根过氧化物酶一起孵育1 h。通过增强化学发光检测装备和Hyperfilm-ECL试剂(发射极耦合逻辑)成像,并用Quantity One图像软件(由Bio-Rad公司提供)分析信号强度。
1.7 统计学分析
实验结果用SPSS16.0软件进行分析,数据用平均值±标准差表示,用方差分析统计分析两组以上的平均值。P值<0.05视为具有统计学意义。
2 结果
2.1 牛蒡甙元对RAFLS的影响
为了检测牛蒡甙元对RAFLS活性的影响,RAFLS以不同作用时间和不同浓度与牛蒡甙元反应,用MTT实验检测细胞活性,乳酸脱氢酶检测牛蒡甙元细胞毒性。如图1所示,牛蒡甙元抑制了RAFLS的增殖。10 μM的牛蒡甙元处理后12和24 h,活性细胞数量无明显影响,但处理后48 h细胞数量减少了23%(P<0.05);20 μM牛蒡甙元处理后12 h,细胞生长未受明显影响,但处理后24和48 h细胞死亡率分别为21%和30%(图1)。另外,30 μM牛蒡甙元作用12小时即可抑制细胞增殖(P<0.05)。图1显示,40~160 μM的牛蒡甙元可以进一步抑制RAFLS增殖,及诱导细胞毒性。本实验中,我们选用了10~30 μM牛蒡甙元来检测其对RAFLS的影响,结果印证了牛蒡甙元对RAFLS增殖的抑制作用。

图1 牛蒡甙元对RAFLS细胞活性的影响。RAFLS(1×104)接种于96孔板上。细胞分别在有或无牛蒡甙元(0~160 μM)的环境下培养48 h(图1A、1B),或在有或无牛蒡甙元(0~30 μM)环境下培养0、12、24、48 h后(图1C)用MTT实验检测细胞活性,并用LDH实验检测牛蒡甙元诱导的细胞毒性。数据用3个独立实验的平均值±标准差表示,*P<0.05, **P<0.01。Fig.1 The effects of arctigenin on the cell viability of RAFLSs. RAFLSs (1×104) were seeded into 96-well plates. Cells were cultured in the presence or absence of arctigenin (0-160 μM) for 48 h (A) and (B) or in the presence or absence of arctigenin (0-30 μM) for 0, 12, 24, and 48 h (C). The cell viability was measured by MTT assay. Arctigenin induced cytotoxicity was assessed by LDH assay. Data are shown as mean ±SD (n=3) from three independent experiments.*P<0.05,**P<0.01, relative to control RAFLSs.
2.2 牛蒡甙元诱导RAFLS凋亡
本实验也同时明确了牛蒡甙元所导致的RAFLS数量减少,除了抑制细胞增殖外,是否与促进细胞凋亡有关。实验用膜细胞凋亡的标志——磷脂酰丝氨酸外部化,来检测细胞凋亡[21]。如表图2所示,牛蒡甙元显著诱导RAFLS凋亡,但10 μM牛蒡甙元诱导作用较弱,早期凋亡比例只有8.61%,而20、30 μM牛蒡甙元诱导作用较强,早期凋亡比例分别为15.16%和22.21%。

图2 牛蒡甙元诱导RAFLS细胞凋亡。RAFLS用相应浓度牛蒡甙元(0~30 μM) 处理48h后用膜联蛋白V(annexin V)和碘化丙啶(PI)双套染,并用流式细胞仪分析凋亡细胞。凋亡细胞是annexin V-阳性而PI-阴性细胞(右下象限)。(图A)RAFLS用相应浓度牛蒡甙元(0-30μM) 处理48h后流式细胞仪结果。(图B)显示RAFLS 的凋亡比例(%)。数据用3个独立实验的平均值±标准差表示,*P<0.05, ** P<0.01。Fig.2 Arctigenin induces RAFLS apoptosis. RAFLSs were treated with various concentrations of arctigenin (0-30 μM) for 48 h and examined by annexin V/ propidium iodide (PI) double staining. Apoptosis was analyzed using flow cytometry. Apoptotic cells were annexin V-positive and PI-negative cells (lower right quadrant). (A) Representative flow cytometry results for RAFLSs exposed to arctigenin (0-30 μM) for 48 h. (B) The histogram shows the apoptosis ratio (%) of RAFLSs. The data represent three independent experiments.*P<0.05,**P<0.01, relative to control RAFLSs.
2.3 牛蒡甙元介导的RAFLS凋亡途径——线粒体途径
为明确牛蒡甙元所激活的凋亡途径,本实验用JC-1检测了RAFLS与牛蒡甙元作用时的线粒体膜电位的变化。图3A显示牛蒡甙元在RAFLS凋亡途径中线粒体膜电位的去极化。在分别与10、20、30 μM牛蒡甙元作用后,RAFLS中低线粒体膜电位的细胞百分比为9.56±1.59%、16.85±1.46%和23.28±2.36%(图3A)。同样,Western blot检测也显示牛蒡甙元能上调促凋亡蛋白Bax,下调抗凋亡蛋白Bcl-2,增加细胞色素C释放进入胞质,并导致胱解酶-9,胱解酶-3和PARP的裂解(图3B-D)
2.4 牛蒡甙元抑制NF-κB和Akt信号通路的活性
既往研究表明NF-κB和Akt信号通路的激活在RAFLS凋亡中起重要作用[8-9],因此本实验用Western blot检测牛蒡甙元对上述信号通路激活的影响(图4)。结果显示牛蒡甙元能抑制p65的核转运及IκBα的降解,提示牛蒡甙元降低NF-κB通路的活性(图4A)。同样,牛蒡甙元可以使Akt的磷酸化减弱,但对总Akt的表达没有影响(图4B)。
3 讨论
本研究提示牛蒡甙元能抑制RAFLS的细胞活性并导致细胞的凋亡。实验中牛蒡甙元处理的RAFLS线粒体膜电位降低,证明牛蒡甙元通过线粒体途径导致RAFLS凋亡。与此同时,牛蒡甙元能上调促凋亡蛋白Bax,下调抗凋亡蛋白Bcl-2,促进细胞色素C释放进入胞质,并加强胱解酶-9,胱解酶-3和PARP的裂解。此外,牛蒡甙元在RAFLS中减弱NF-κB和Akt信号通路的活性。提示牛蒡甙元通过线粒体途径,以及抑制NF-κB和Akt信号通路,从而导致RAFLS的凋亡。
RAFLS是RA的发生和病理过程的主要效应细胞之一,其主要特点为快速增殖和凋亡缺陷[22]。本实验研究了牛蒡甙元对RAFLS的细胞活性及凋亡的影响,研究发现牛蒡甙元可以降低RAFLS的细胞活性并诱导凋亡。研究结果提示牛蒡甙元诱导的细胞毒性可能是中药牛蒡能缓解类风湿性关节炎的机制之一。而且,牛蒡甙元已证实可以抑制人类卵巢癌OVCAR3和SKOV3细胞的增殖[23],提高人类肺癌细胞H460对顺铂的化疗敏感性[24],减低膀胱癌细胞T24的细胞活性,诱导肺癌细胞A549、肝癌细胞HepG2和胃癌细KATO III的细胞毒作用[18]。总之,上述结果显示牛蒡甙元对多种肿瘤或肿瘤样细胞的增殖有抑制作用。
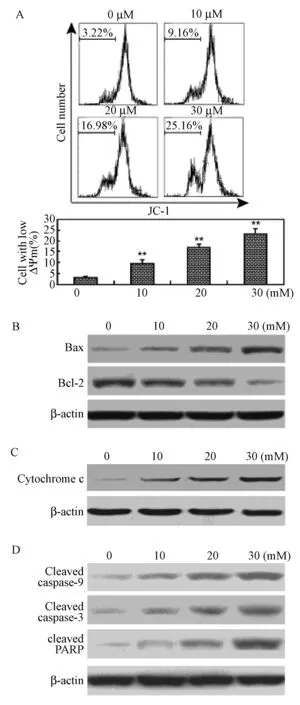
图3 牛蒡甙元诱导RAFLS线粒体凋亡。(图A)RAFLS用相应浓度牛蒡甙元(0~30 μM) 处理48 h后进行然后用JC-1染色。并通过FACS分析JC-1平均荧光密度。上图1分别是3个实验结果。下图1为定量FACS分析。(图B、C、D)RAFLS用相应浓度牛蒡甙元(0~30 μM) 处理48 h后进行Western blot分析。 数据用3个独立实验的平均值±标准差表示(n=3),*P<0.05, **P<0.01。Fig.3 Arctigenin induces mitochondrial apoptosis of RAFLSs. (A) Cells were treated with indicated concentrations of arctigenin (0-30 μM) for 48 h and stained with JC-1. The mean JC-1 fluorescence intensity was detected by FACS analysis. Images are representative of three independent experiments (upper panel). Quantification of FACS analysis is shown in the lower panel. (B)-(D) Cells were treated with indicated concentrations of arctigenin (0-30 μM) for 48 h and subjected to Western blot analysis. Data are shown as mean±SD (n=3) from three independent experiments. *P<0.05,**P<0.01, relative to control RAFLSs.
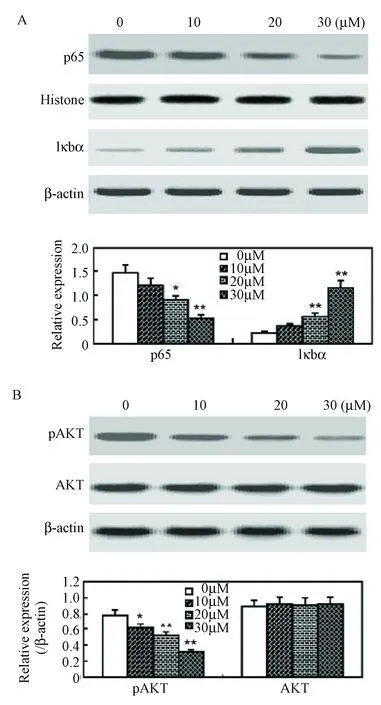
图4 牛蒡甙元抑制RAFLS中的NF-κB和Akt信号通路。RAFLS用相应浓度牛蒡甙元(0~30 μM)处理48 h后进行Western blot分析。上图A显示p65核转位和IκBα的降解,上图B显示Akt的磷酸化。下图显示定量Western blot分析。 数据用3个独立实验的平均值±标准差表示 (n=3),*P<0.05,**P<0.01。Fig.4 Arctigenin inhibits NF-κB and Akt signaling pathways in RAFLSs. RAFLSs were treated with indicated concentrations of arctigenin (0-30 μM) for 48 h and subjected to Western blot analysis. The nuclear translocation of p65 and the degradation of IκBα (A) and the phosphorylation of Akt (B) are shown in the upper panel. Quantification of Western blot analysis is shown in the lower panel. Data are shown as mean±SD (n=3) from three independent experiments. *P<0.05,**P<0.01, relative to control RAFLSs.
内在的凋亡通路包括非受体介导的细胞内信号,通过降低线粒体膜电位导致细胞凋亡[25]。研究表明线粒体途径对RAFLS凋亡有重要作用。为了研究牛蒡甙元诱导RAFLS凋亡的机制,本实验检测牛蒡甙元处理后RAFLS线粒体膜电位,发现牛蒡甙元增加了RAFLS中线粒体低电位细胞的比例,故导致线粒体膜电位的减低。此外已有研究证实牛蒡甙元通过线粒体途径引起膀胱癌细胞T24的凋亡[17]。与此同时,Bcl-2蛋白家族是线粒体凋亡途径的重要调节蛋白,家族成员包括促凋亡蛋白和抗凋亡蛋白[26]。RAFLS中抗凋亡蛋白Bcl-2的表达对线粒体稳态和细胞活性十分重要[7]。牛蒡甙元处理后RAFLS中Bcl-2表达减少,Bax是Bcl-2家族的一种促凋亡蛋白,可以同其他促凋亡因子组成同源或异源二聚体,比如Bak,以破坏线粒体膜的完整性,并导致细胞色素酶C等凋亡因子释放[27]。本实验结果显示牛蒡甙元能上调Bax的表达。牛蒡甙元处理的RAFLS中Bcl-2的下调和Bax的上调表明Bcl-2和Bax可能参与牛蒡甙元介导的线粒体途径。此外本实验观察到细胞色素C释放到胞质,同样其细胞色素C的释放也被显著增强。一旦细胞色素C释放,凋亡蛋白酶—血小板激活因子1(Apaf1)、胱解酶-9前体和ATP将受到抑制,并诱导胱解酶-9裂解[28]。裂解和激活的胱解酶-9诱导胱解酶-3的激活,导致细胞骨架、核骨架、基因修复和细胞周期蛋白的裂解[29]。牛蒡甙元处理后RAFLS中胱解酶-9、胱解酶-3和PARP的裂解增强,揭示了胱解酶-9和胱解酶-3参与了牛蒡甙元介导的凋亡。综上所述,牛蒡甙元处理后RAFLS通过线粒体途径诱导凋亡。
RA滑膜细胞中NF-κB的表达和激活已被既往研究所广泛证实,NF-κB激活后可通过促进RAFLS增殖及抑制凋亡而使滑膜细胞增生[30]。研究证实牛蒡甙元抑制p65的核转位以及IκBα的降解,从而抑制NF-κB的活性,这也显示抑制NF-κB的活性是牛蒡甙元诱导凋亡的途径之一。 Akt在RAFLS活性中也有重要作用[9,31],故本实验也研究了牛蒡甙元对Akt磷酸化的影响。研究发现牛蒡甙元减低RAFLS中Akt的磷酸化。所以,牛蒡甙元诱导的凋亡也可能是通过对NF-κB和Akt通路的抑制实现的。
总之,本实验结果显示牛蒡甙元通过线粒体途径的激活和NF-κB及Akt信号通路的抑制这两条途径,实现其对RAFLS的细胞毒性及凋亡的影响。体外实验表明牛蒡甙元可能是治疗类风湿性关节炎的新药剂,然而在进入临床试验之前,仍需进行大量检验牛蒡甙元抗增殖和抗炎效应的类风湿关节炎动物实验。另外,对牛蒡甙元同其他经典药物在治疗类风湿性关节炎上的协同、增效或拮抗作用也需要进一步研究。
