F1代转CuZnSOD基因猪的制备与研究
2016-07-11王守栋房国锋曾勇庆李川皓王延东
王守栋,房国锋,曾勇庆*,陈 伟,李川皓,王延东
(1.山东农业大学动物科技学院,泰安 271000; 2.山东省动物生物工程与疾病防治重点实验室,泰安 271000)
F1代转CuZnSOD基因猪的制备与研究
王守栋1,2,房国锋1,2,曾勇庆1,2*,陈伟1,2,李川皓1,2,王延东1,2
(1.山东农业大学动物科技学院,泰安 271000; 2.山东省动物生物工程与疾病防治重点实验室,泰安 271000)
摘要:为进一步研究CuZnSOD基因对肉质表达调控的作用机理,探究通过磁性纳米颗粒介导精子载体法制备转基因猪的可行性。本试验在重组质粒先后与磁性纳米颗粒和精液共孵育的基础上,再通过人工授精获得F1代仔猪。对繁殖得到的F1代个体进行PCR、荧光定量PCR及Western blot检测,对屠宰的4头阳性猪采用绝对定量PCR进行转基因拷贝数鉴定,F1代个体达100 kg左右时抽样屠宰进行肉质及抗氧化性能测定。结果表明,F1代阳性率为30.23%。阳性组CuZnSOD在5种组织中的RNA和蛋白水平上的表达量较阴性组均上调。本试验得到的标准曲线:ΔCt=-3.121 6 lgN+17.281(R2=0.992 1,P<0.001),4头阳性猪中外源基因拷贝数分别为25.89、80.61、106.69、83.03。肉质及抗氧化性能测定结果显示,阳性组肌肉滴水损失显著低于阴性组(P<0.05);阳性组肌肉T-SOD1d、T-SOD2d活性显著高于阴性组,MDA1d、MDA2d含量显著低于阴性组(P<0.05)。研究结果表明:CuZnSOD基因的过表达可以提高肌肉的抗氧化性能,降低滴水损失,进而改善肉质,同时,证明磁性纳米颗粒介导的精子载体法可以用于制备转基因动物。
关键词:CuZnSOD;磁性纳米颗粒;转基因猪;肉质;抗氧化性能
近年来,猪的遗传改良在提高生长速度、增加胴体瘦肉率等方面取得了很大的进步,但肉质特性却停滞不前甚至随之变劣,而消费者对猪肉肉质的要求却在逐渐提高,所以,肉质性状在猪的育种目标中也越来越受到研究者的重视[1]。
CuZnSOD 是一种金属酶,属于超氧化物歧化酶(SODs)家族,能够转移性地清除超氧阴离子自由基,维持氧自由基平衡,帮助机体防御氧化损伤[2]。肉品在货架期酸败的主要原因是脂质氧化,SOD可以阻止脂质氧化反应的进行,改善肉质,延长肉品的货架期[3-4]。杜金芳等[5]利用RACE的方法克隆得到了猪CuZnSOD基因的cDNA全长序列。石元[6]利用精子载体法成功制备了转基因肉兔模型并验证了CuZnSOD对抗氧化性能等肉质性状的重要影响。
精子载体法是利用精子头部可捕获外源DNA的特性,将外源目的基因与精子共孵育,使其在受精过程中导入受精卵,并稳定整合到子代基因组中的一种制备转基因动物的方法[7-8]。磁性纳米基因载体技术是以磁性纳米颗粒作为基因载体,利用磁性纳米颗粒能被细胞有效内吞,实现核酸向细胞内的转运和释放,并在磁场的作用下实现靶向性基因治疗[9-11]。庄乾元等[12]用磁性纳米颗粒作为基因载体携带质粒pEGFP-C1,转染人的细胞,结果表明磁性纳米颗粒转染后的细胞增殖活性及功能略优于脂质体对照组。同时S.W.Gersting等[13]将质粒与磁性纳米颗粒形成复合物转染猪的细胞,发现磁性纳米颗粒的转化效率比dendrimer、PEI和Lipofectamine介导的转染效率都高。本研究通过磁性纳米颗粒介导精子载体法成功制备了CuZnSOD转基因猪并对外源基因拷贝数进行了鉴定,旨在研究CuZnSOD基因对抗氧化性能等肉质特性的影响机理,对培育具有高抗氧化性的猪种资源具有重要意义。
1材料与方法
1.1试验材料和试验动物
磁性纳米颗粒polyMAG购自Chemicell;PCR引物由上海生工合成;Anti-SOD1购自Santa Cruz、Ⅱ抗HRP购自EarthOX;Anti-GAPDH、Ⅱ抗HRP购自碧云天;促排卵素3号购自宁波第二激素厂;SOD、MDA试剂盒购自南京建成;其他试剂为国产或进口分析纯。
选取3头繁殖性能良好、胎次相同并且同期发情的大白母猪和1头精液质量良好的大白公猪作为试验动物,经相应处理并配种。F1代仔猪出生后采集耳组织并迅速投入70%乙醇中,用于PCR检测。仔猪在同样的饲养管理条件下饲喂至100 kg左右时抽样空腹屠宰,其中阳性猪4头,阴性猪2头。屠宰后取背最长肌肌肉、肝、背膘、肾、肺组织样并迅速投入液氮,用于荧光定量PCR和Western blot检测。取背最长肌放入带有冰袋的泡沫箱中带回实验室用于肉质测定。
1.2试验方法1.2.1无内毒素质粒的提取及线性化处理用实验室前期构建的质粒[6]转化大肠杆菌感受态DH5α菌株,并接种于卡那霉素抗性(50 μg·mL-1)的琼脂糖平板上,挑取阳性菌落经扩大培养后,利用QIAGEN的无内毒素质粒大提试剂盒提取质粒。用限制性内切酶VspⅠ酶切提取的重组质粒,酶切反应液经1%琼脂糖凝胶电泳后进行回收纯化。
1.2.2CuZnSOD转基因猪的制备将400 μg按1∶3比例混合的线性质粒和环状质粒与500 μL 磁性纳米颗粒polyMAG混匀,室温孵育10 min后,将质粒-纳米混合物逐滴加入到稀释好的精液中,上下翻转混匀,室温孵育30 min后进行人工授精,同时每头母猪注射2 mL促排卵素3号;第2天重复以上操作步骤。
1.2.3F1代仔猪PCR检测与测序本研究中转入的重组质粒pIRES2-AcGFP1-CuZnSOD上的CuZnSOD序列是猪本身CuZnSOD基因的CDS区,因此,在设计引物对该序列检测时采用跨越内含子的方法进行设计。根据重组质粒的序列,用Primer5设计针对3个序列的引物来检测F1代猪的阳性率,引物信息如表1。用基因组提取试剂盒提取试验所需的基因组,用3对特异性引物分别进行PCR扩增,将产物进行1%的琼脂糖凝胶电泳检测,对目的条带进行回收测序。
表1特异性引物信息
Table 1The information of specific primers
1.2.4荧光定量PCR检测不同组织CuZnSOD基因的表达用Trizol试剂法提取5种组织的总RNA,然后用TaKaRa反转录试剂盒将RNA反转录成cDNA,-20 ℃保存。荧光定量采用SYBR Green染料法,在Stratagene Mx3000P荧光定量PCR仪上进行反应。以GAPDH基因作为内参[14],用Primer5.0设计针对CuZnSOD基因的1对特异性引物,引物名称为F1、R1和FGAPDH、RGAPDH,引物序列信息见表2。
表2定量PCR引物序列
Table 2Primers sequence of qPCR
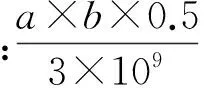
用Primer5.0设计2对特异性引物,以ACTB作为内参标化基因组DNA的量。用CMV-F和CMV-R扩增转基因片段,用ACTB-F和ACTB-R扩增持家基因ACTB的片段,引物序列信息见表2。
反应体系为20 μL,反应条件:95 ℃预变性30 s;95 ℃变性5 s,58 ℃复性10 s,72 ℃延伸15 s,40个循环;最后再72 ℃延伸5 min。每个待测样品设置3个生物学重复。将外源基因片段检测引物扩增CtCMV减去相应的ACTB基因的扩增CtACTB,得到ΔCt,再对样品的拷贝数的对数值lgN作图得到绝对定量的标准曲线。待反应结束后根据相应的Ct值及标准曲线计算外源基因的拷贝数。
1.2.7F1代试验猪肉质与抗氧化性能测定[16-18]肉色及大理石花纹采用美国标准5级评分图对照评定;pH采用PHB-4型便携式pH计进行测定;滴水损失、烹饪损失、失水率分别用吊挂处理法、蒸煮法、压力法进行测定;肌肉嫩度采用C-LM3型肌肉嫩度计进行测定。肉样0~4 ℃保存5 d内的抗氧化性能指标T-SOD及MDA按照南京建成试剂盒说明书进行测定。
1.2.8统计分析本试验数据采用SPSS 13.0软件进行单因素方差分析,所得的数值均以“平均值±标准误”表示,P<0.05为差异显著,有统计学意义。
2结果
2.1F1代转基因猪PCR检测及阳性率统计
3对特异性引物都能扩增出目的条带的为阳性猪,部分检测结果如图1。3头试验母猪均顺利受孕并产仔43头,其中阳性猪13头,阴性猪30头,阳性率为30.23%。
2.2F1代猪不同组织CuZnSOD基因的表达分析
F1代猪阳性组与阴性组5种组织CuZnSOD基因mRNA相对表达量见图2。结果表明,阳性组5种组织中CuZnSOD基因mRNA表达量高于阴性组,在肌肉和肺组织中达到差异显著水平(P<0.05)。
2.3F1代猪不同组织CuZnSOD蛋白的表达分析
F1代猪阳性组与阴性组5种组织CuZnSOD蛋白相对表达量见图3。结果表明,阳性组5种组织中CuZnSOD蛋白的表达量都高于阴性组,在肌肉和肝组织中达到差异显著水平(P<0.05)。
2.4转CuZnSOD基因阳性猪外源基因拷贝数的鉴定
本试验精确度高,同一标准品的3次重复Ct值变化很小,得到的绝对定量标准曲线(图4),决定系数R2=0.992 1,计算公式为:ΔCt=-3.121 6 lgN+17.281(R2=0.992 1,P<0.001)。CMV和ACTB基因扩增产物的熔解曲线见图5。每个样品进行3次独立重复试验,各样品关于CMV和ACTB基因的3次定量检测的Ct值标准误小,重复性好,外源基因拷贝数鉴定结果如表3。
2.5试验猪肉质与抗氧化性能测定结果
对F1代阳性猪与阴性猪进行常规肉质测定,结果表明,阳性组肌肉滴水损失显著小于阴性组(P<0.05),其他指标较阴性组均有所改善但差异不显著(表4)。
对屠宰后0~4 ℃保存5 d内的肌肉组织的抗氧化性能指标T-SOD及MDA进行测定,结果表明,阳性组T-SOD活性始终高于阴性组,并且阳性组T-SOD1d、T-SOD2d活性显著高于阴性组(P<0.05)。阳性组MDA含量始终低于阴性组,并且阳性组MDA1d、MDA2d含量显著低于阴性组(P<0.05,图6)。
表3转基因阳性猪中外源基因拷贝数的估算
Table 3Estimate of the exogenous gene copy number in transgenic positive pigs
表4F1代阳性组与阴性组的肉质测定结果
Table 4Results of meat quality in positive group and negative groups of F1generation
同一行标注不同字母的数值间差异显著(P<0.05)
Values with the different letters within the same row differ significantly(P<0.05)
3讨论
目前传统的转基因动物制备方法有显微注射法[19]、胚胎干细胞法[20]和逆转录病毒感染法[21]等。精子载体法的优点主要是利用生殖细胞受精的自然过程,操作简单,提高了转基因效率。M.Lavitrano等[7]用小鼠精子作为载体,体外受精获得转基因小鼠,证实了精子载体法生产转基因动物的可行性;J.H.Kim等[22]利用精子载体法获得了转基因小鼠和猪。徐正刚等[23]通过精子介导法制备了RNA干扰PID1转基因肉兔模型。纳米颗粒作为基因载体具有装载容量大,浓缩并保护外源DNA分子,提高转染效率等优点。2011年,V.F.Campos等[24]将纳米聚合物和HCNs应用于精子载体法中,结果表明,纳米载体结合外源DNA再与精子共孵育后,能够获得更高的基因结合内化效率。这为磁性纳米颗粒介导精子载体法制备转基因动物提供了参考。
需氧生物在代谢过程中会产生大量的自由基,过多的自由基若不能及时被清除,会攻击各种生物大分子,引起DNA损伤、酶失活、脂质过氧化等一系列氧化损伤,进而引起生物体各种生理病变[25]。肌肉在货架期的氧化以脂质氧化为主,自由基对肌肉中的PUFA具有很高的亲和力,可引发脂质氧化,形成不稳定的ROOH,并迅速降解成MDA等物质[3]。SOD能将超氧阴离子自由基快速歧化为H2O2和O2,然后H2O2在CAT和GPX的作用下被转化为H2O和O2,从而解除超氧阴离子自由基所造成的危害[26]。李华等[17]研究结果表明,SOD活性越高、MDA含量越低的肌肉,其系水力越高、肉色越鲜艳,并且肉质越细嫩。
本研究获得转基因阳性率为30.23%,阳性率检测参照R.Drews等[27]在试验中采用的设计2条引物来检测体系的可靠性的方法,针对3段不同基因的序列设计3对特异性引物来进行PCR检测,3对引物都跑出目的条带的个体为阳性个体,以此来提高阳性率检测的准确性。本研究从RNA和蛋白质水平上检测了阳性组与阴性组的相对表达量,试验结果表明,阳性组CuZnSOD基因在RNA和蛋白质水平上的表达量均上调,这些结果进一步说明我们通过磁性纳米颗粒介导精子载体法成功制备了转CuZnSOD猪。尽管阳性组CuZnSOD基因在RNA和蛋白质水平上的表达量均上调,但是不同组织阳性组与阴性组在RNA或蛋白质水平上却存在着表达差异,这可能与转入质粒上的CMV启动子有关,相关研究表明,CMV启动子可以在不同组织中引起不同水平的表达[28-29]。转基因的整合位点与拷贝数是影响外源基因表达的主要因素,鉴定转基因阳性猪外源基因拷贝数是后续表型研究的基础。绝对定量PCR技术与传统的定量技术如Southern blot相比,具有高特异性、高安全性、高效率、高通量、低成本等优点[30]。本研究建立了实时定量PCR方法检测转基因猪中外源基因拷贝数的方法。由于染料 SYBR GreenⅠ对所有DNA双链具有高选择性,为保证扩增片段的特异性,引物的选择至关重要[31]。本试验所设计的两对检测引物的扩增片段大小相近,Tm值基本相等,扩增效率基本一致,并且PCR扩增产物电泳和熔解曲线很好的验证了引物扩增的特异性。本试验肌肉抗氧化性能及肉质测定结果表明,阳性组的肉质较阴性组有所改善,0~4 ℃保存5 d内的肌肉组织中,阳性组T-SOD活性始终高于阴性组,MDA含量始终低于阴性组,这与李华等[17]研究结果基本一致。滴水损失、烹饪损失、压力损失等是能够反映猪肉持水能力(WHC)的指标[32]。本试验阴性组的滴水损失显著高于阳性组,可能是由于阴性猪SOD表达量低于阳性猪,阴性猪屠宰后,机体氧化还原系统失衡,产生过量的ROS。这些自由基未能被及时清除,攻击生物膜中的不饱和脂肪酸,使之发生过氧化反应,细胞膜的结构和功能遭到破坏,细胞内液释出,导致滴水损失增多,持水能力减弱[33]。因此,本研究通过磁性纳米颗粒介导精子载体法制备转CuZnSOD猪证明是可行的,并由此进一步验证了CuZnSOD基因对肉质表达调控的作用机理。
参考文献(References):
[1]曾勇庆,王根林,王林云,等.猪肉质性状基因及其遗传标记的研究进展[J].畜牧与兽医,2003,35(8):37-40.
ZENG Y Q,WANG G L,WANG L Y,et al.The research progress of the gene and its genetic markers related to meat quality in pigs[J].AnimalHusbandryandVeterinaryMedicine,2003,35(8):37-40.(in Chinese)
[2]ZYRACKA E,ZADRAG R,KOZIOS,et al.Yeast as a biosensor for antioxidants:simple growth tests employing a Saccharomyces cerevisiae mutant defective in superoxide dismutase[J].ActaBiochimPol,2005,52(3):679-684.[3]FERNNDEZ J,PÉREZ-LVAREZ J A,FERNNDEZ-LPEZ J A.Thiobarbituric acid test for monitoring lipid oxidation in meat[J].FoodChem,1997,59(3):345-353.
[4]DU J F,ZENG Y Q,WANG H,et al.CuZnSODgene expression and its relationship with anti-oxidative capacity and pork quality[J].SAfrJAnimSci,2010,40(3):265-272.
[5]杜金芳,曾勇庆,陈伟,等.猪CuZnSOD基因的克隆、表达及功能分析[J].遗传,2010,32(10):1037-1042.
DU J F,ZENG Y Q,CHEN W,et al.Cloning,expression and functional analysis ofCuZnSODgene in swine[J].Hereditas,2010,32(10):1037-1042.(in Chinese)
[6]石元.猪CuZnSOD基因核心启动子鉴定及精子介导法制备CuZnSOD转基因肉兔的研究[D].泰安:山东农业大学,2013.
SHI Y.Identification of core promoter of pigCuZnSODgene and the study ofCuZnSODgene transgenic rabbit by sperm-mediated gene transfer[D].Tai’an:Shandong Agricultural University,2013.(in Chinese)
[7]LAVITRANO M,CAMAIONI A,FAZIO V M,et al.Sperm cells as vectors for introducing foreign DNA into eggs:genetic transformation of mice[J].Cell,1989,57(5):717-723.
[8]杜伟,崔海信,王琰,等.精子载体法制备转基因动物的研究进展[J].生物技术通报,2012,(12):13-18.
DU W,CUI H X,WANG Y,et al.Research progress on spermatozoa-mediated gene transfer to product transgenic animals[J].BiotechnologyBulletin,2012,(12):13-18.(in Chinese)
[9]BERTRAM J.MATra-Magnet assisted transfection:combining nanotechnology and magnetic forces to improve intracellular delivery of nucleic acids[J].CurrPharmBiotechnol,2006,7(4):277-285.
[10]PANYAM J,ZHOU W Z,PRABHA S,et al.Rapid endo-lysosomal escape of poly(DL-lactide-co-glycolide) nanoparticles:implications for drug and gene delivery[J].FASEBJ,2002,16(10):1217-1226.
[11]DOBSON J.Gene therapy progress and prospects:magnetic nanoparticle-based gene delivery[J].GeneTher,2006,13(4):283-287.
[12]庄乾元,孙璇,周四维,等.葡聚糖磁性纳米颗粒在基因转染中的应用[J].武汉大学学报(医学版),2007,28(3):270-273,286.
ZHUANG Q Y,SUN X,ZHOU S W,et al.Application of dextran coated magnetic iron oxide nanoparticles in delivering target gene into cellsinvitro[J].MedicalJournalofWuhanUniversity,2007,28(3):270-273,286.(in Chinese)
[13]GERSTING S W,SCHILLINGER U,LAUSIER J,et al.Gene delivery to respiratory epithelial cells by magnetofection[J].JGeneMed,2004,6(8):913-922.
[14]BARBER R D,HARMER D W,COLEMAN R A,et al.GAPDHas a housekeeping gene:analysis ofGAPDHmRNA expression in a panel of 72 human tissues[J].PhysiolGenomics,2005,21(3):389-395.
[15]YUAN J S,WANG D,STEWART C NJR.Statistical methods for efficiency adjusted real-time PCR quantification[J].BiotechnolJ,2008,3(1):112-123.
[16]HONIKEL K O.Reference methods for the assessment of physical characteristics of meat[J].MeatSci,1998,49(4):447-457.
[17]李华,曾勇庆,魏述东,等.猪宰后肌肉SOD与MDA的变化及其对肉质特性的影响[J].畜牧兽医学报,2010,41(3):257-261.
LI H,ZENG Y Q,WEI S D,et al.Changes of superoxide dismutase activity and malondialdehyde level in postmortem muscle and their association with meat quality in pigs[J].ActaVeterinariaetZootechnicaSinica,2010,41(3):257-261.(in Chinese)
[18]张天阳,徐海鹏,曾勇庆,等.饲喂不同剂量乳酸菌液对肥育猪肉质特性及抗氧化性的影响[J].养猪,2014(1):41-43.
ZHANG T Y,XU H P,ZENG Y Q,et al.Effect of feeding different doses of lactic acid bacteria on meat quality and antioxidant properties of finishing pigs[J].SwineProduction,2014(1):41-43.(in Chinese)
[19]GORDON J W,SCANGOS G A,PLOTKIN D J,et al.Genetic transformation of mouse embryos by microinjection of purified DNA[J].ProcNatlAcadSci,1980,77(12):7380-7384.
[20]GOSSLER A,DOETSCHMAN T,KORN R,et al.Transgenesis by means of blastocyst-derived embryonic stem cell lines[J].ProcNatlAcadSci,1986,83(23):9065-9069.
[21]SALTER D,BALANDER R,CRITTENDEN L.Evaluation of Japanese quail as a model system for avian transgenesis using avian leukosis viruses[J].PoultSci,1999,78(2):230-234.
[22]KIM J H,JUNG-HA H S,LEE H T,et al.Development of a positive method for male stem cell-mediated gene transfer in mouse and pig[J].MolReprodDev,1997,46(4):515-526.
[23]徐正刚,杨伦,曾勇庆,等.RNA 干扰沉默PID1 基因肉兔模型的构建[J].畜牧兽医学报,2014,45(5):750-756.
XU Z G,YANG L,ZENG Y Q,et al.Construction of transgenic rabbit model by RNA interferencingPID1 gene[J].ActaVeterinariaetZootechnicaSinica,2014,45(5):750-756.(in Chinese)
[24]CAMPOS V F,DE LEON P M,KOMNINOU E R,et al.NanoSMGT:transgene transmission into bovine embryos using halloysite clay nanotubes or nanopolymer to improve transfection efficiency[J].Theriogenology,2011,76(8):1552-1560.
[25]JIN L H,BAHN J H,EUM W S,et al.Transduction of human catalase mediated by an HIV-1 TAT protein basic domain and arginine-rich peptides into mammalian cells[J].FreeRadicBioMed,2001,31(11):1509-1519.
[26]MICHIELS C,RAES M,TOUSSAINT O,et al.Importance of Se-glutathione peroxidase,catalase,and Cu/Zn-SOD for cell survival against oxidative stress[J].FreeRadicBioMed,1994,17(3):235-248.
[27]DREWS R,DROHAN W N,LUBON H.Transgene detection in mouse tail digests[J].Biotechniques,1994,17(5):866-867.
[28]KONG Q,WU M,HUAN Y,et al.Transgene expression is associated with copy number and cytomegalovirus promoter methylation in transgenic pigs[J].PLoSOne,2009,4(8):e6679.
[29]ZHANG Y,XI Q,DING J,et al.Production of transgenic pigs mediated by pseudotyped lentivirus and sperm[J].PLoSOne,2012,7(4):e35335.
[30]王晓建,杨旭,宋晓东,等.实时荧光定量PCR法检测转基因小鼠拷贝数[J].中国实验动物学报,2007,15(3):170-174.
WANG X J,YANG X,SONG X D,et al.Absolute quantitative real-time PCR assay for determining transgene copy number in transgenic mice[J].ActaLaboratoriumAnimalisScientiaSinica,2007,15(3):170-174.(in Chinese)
[31]INGHAM D J,BEER S,MONEY S,et al.Quantitative real-time PCR assay for determining transgene copy number in transformed plants[J].Biotechniques,2001,31(1):132-134.
[32]FRANCO D,VAZQUEZ J A,LORENZO J M.Growth performance,carcass and meat quality of the Celta pig crossbred with Duroc and Landrance genotypes[J].MeatSci,2014,96(1):195-202.
[33]陈代文,张克英,胡祖禹.猪肉品质特征的形成原理[J].四川农业大学学报,2002,20(1):60-66.
CHEN D W,ZHANG K Y,HU Z Y.The mechanism for developing pork quality traits[J].JournalofSichuanAgriculturalUniversity,2002,20(1):60-66.(in Chinese)
(编辑郭云雁)
Preparation and Study ofCuZnSODGene Transgenic Pigs in F1Generation
WANG Shou-dong1,2,FANG Guo-feng1,2,ZENG Yong-qing1,2*,CHEN Wei1,2,LI Chuan-hao1,2,WANG Yan-dong1,2
(1.CollegeofAnimalScienceandVeterinaryMedicine,ShandongAgriculturalUniversity,Taian271000,China;2.ShandongProvincialKeyLaboratoryofAnimalBiotechnologyandDiseaseControlandPrevention,Taian271000,China)
Key words:CuZnSOD;magnetic nanoparticles;transgenic pigs;meat quality;antioxidant properties
Abstract:The aim of this study was to further research the mechanism of howCuZnSODgene effect the meat quality by expression and regulation,and explore the possibility to generate transgenic pigs by sperm carrier method mediated by magnetic nanoparticles.F1generation individuals were obtained by artificial insemination on the basis of the recombinant vector incubated with magnetic nanoparticles and semen,respectively.F1individuals were detected by PCR,qRT-PCR and Western blot.Exogenous gene copy number in the 4 slaughtered transgenic positive pigs were estimated by absolute qPCR.Individuals from positive and negative groups were selected at the weight of about 100 kg and slaughtered to determine the meat quality and the antioxidant properties.The results showed that:The positive rate was 30.23% in F1individuals.Compared with the negative group,theCuZnSODexpression at RNA and protein levels in 5 tissues were up-regulated in positive group.The standard curve in this study was:ΔCt=-3.121 6 lgN+17.281(R2=0.992 1,P<0.001).The exogenous gene copy number in the 4 positive pigs was 25.89,80.61,106.69 and 83.03,respectively.The results also showed that:Compared with the negative group,the positive group was significantly increased in muscle T-SOD1dand T-SOD2dactivity(P<0.05),but decreased in muscle drip loss,the content of MDA1dand MDA2d(P<0.05),respectively.The results indicated that the overexpression ofCuZnSODgene could elevate the antioxidant properties,reduce the drip loss and further improve the meat quality.Meanwhile,our results proved that transgenic animals could be generated by sperm carrier method mediated by magnetic nanoparticles.
doi:10.11843/j.issn.0366-6964.2016.01.003
收稿日期:2015-01-12
基金项目:国家转基因重大专项(2013ZX08006-002;2011ZX08006-002);山东省现代农业(生猪)产业技术体系建设专项(SDAIT-06-022-03);山东省农业良种工程重大课题(2013LZ02-015)
作者简介:王守栋(1989-),男,山东新泰人,硕士生,主要从事动物遗传育种学研究,E-mail:wangshoudong.8@163.com *通信作者:曾勇庆,教授,博士生导师,主要从事动物遗传育种学研究,E-mail:yqzeng@sdau.edu.cn
中图分类号:S828;S813.3
文献标志码:A
文章编号:0366-6964(2016)01-0016-09
