几种微藻饵料对缢蛏稚贝脂质营养组成的影响
2016-06-29冉照收于珊珊周海波徐继林严小军
周 率, 冉照收, 于珊珊, 周海波, 徐继林, 严小军
(1. 宁波大学海洋学院 应用海洋生物技术教育部重点实验室, 宁波 315211;2. 福建省宝智水产科技有限公司, 漳州 363503)
几种微藻饵料对缢蛏稚贝脂质营养组成的影响
周 率1, 冉照收1, 于珊珊1, 周海波2, 徐继林1, 严小军1
(1. 宁波大学海洋学院 应用海洋生物技术教育部重点实验室, 宁波 315211;2. 福建省宝智水产科技有限公司, 漳州 363503)
摘 要通过用4种缢蛏人工养殖中常用的饵料微藻(角毛藻、球等鞭金藻、威氏海链藻、假微型海链藻),以单种饵料和混合饵料的形式对缢蛏稚贝进行投喂实验,研究不同的微藻饵料对缢蛏稚贝的脂肪酸和甾醇组成的影响。对喂食饵料后的缢蛏稚贝进行脂肪酸甾醇的GC-MS分析,共检测到16种脂肪酸和7种甾醇,不同的单种饵料及混合饵料的缢蛏投喂组的脂肪酸、甾醇差异很大。C16:2在含有威氏海链藻的单种饵料和混合饵料的投喂组被检测到;金藻混合藻饵料的缢蛏投喂组脂肪酸种类最丰富(13~15种),PUFA百分含量最高,胆甾醇含量不高但富含菜子甾醇。因此,从微藻饵料对缢蛏稚贝的脂质营养化影响上来说,金藻的混合饵料是最佳饵料。
关键词微藻;缢蛏;脂肪酸;甾醇;气象色谱-质谱连用分析
脂肪酸与甾醇都是人类重要的脂质营养来源,同时也是人体脂肪的重要组成部分。不饱和脂肪酸中的ω-3与ω-6系多不饱和脂肪酸尤为重要,它们对婴幼儿的大脑发育有重要的作用[1, 2]。甾醇是人类重要的脂类营养物质,也是真核生物膜的重要组成部分。研究表示,植物甾醇具有降血脂、抑制身体对胆固醇的吸收作用[3, 4]。
缢蛏(Sinonovaculaconstricta, Lamarck),属软体动物门竹蛏科缢蛏属,是中国常见的贝类海产品。贝类富含对人体有重要作用的脂肪酸和甾醇,尤以多不饱和脂肪酸和植物甾醇[5, 6]极为丰富,缢蛏还富含人体生命活动所需的各种必需氨基酸、不饱和脂肪酸等营养物质[7]。国内对缢蛏生物大分子物质的研究也很多,赵艳景等从缢蛏体内分离提纯出抗氧化活性很强的缢蛏多糖[8],林文东等发现缢蛏水提物具有体外抗氧化活性[9],抗氧化作用是抗疲劳作用的基础,能加速疲劳修复[10]。本研究主要研究缢蛏的脂肪酸、甾醇受微藻饵料的影响。
研究显示,贝类几乎不具备自身合成和转化甾醇的能力,所以饵料微藻不仅影响人工养殖贝类的生长情况,而且在很大程度上,其体甾醇来源也受微藻饵料脂质组成的影响[11,12]。不同贝类,养殖所用的适宜微藻不同。马斌[13]等通过研究8种常见饵料微藻对缢蛏稚贝存活率和成长速度的影响,认为角毛藻(Chaetoceroscalcitrans)和球等鞭金藻(Isochrysisgalbana)的单藻饵料效果最好,其优于绿色巴夫藻(Pavlovaviridis)、青岛大扁藻(Platymonashelgolandica)等。因此,在周海波的实验结果基础上,本研究选择了饵料效果最佳的角毛藻和金藻,并增加了近期在养殖生产中应用较好,较为感兴趣的假微型海链藻(Thalassiosirapseudonana)和威氏海链藻(Thalassiosiraweissflogii)2种饵料微藻,希望通过研究这4种在贝类养殖上饵料效果良好的微藻及其混合藻对缢蛏稚贝脂质组成的影响,进而从缢蛏的肉质脂质营养优化角度,为缢蛏稚贝的养殖饵料筛选提供更有价值的数据。
1材料和方法
1.1微藻饵料的培养
本研究的饵料微藻(角毛藻、球等鞭金藻、假微型海链藻和威氏海链藻)藻种均由宁波大学海洋生物实验室藻种实验室提供,采用“NML3号”培养液进行培养。藻种于2.5 L锥形瓶中培养1周(20℃±2℃)后转入50 L白色塑料桶中,扩大培养约1周,培养温度23℃~27℃,均在自然光下进行培养。每天用CASY-TT颗粒粒度计数分析仪(德国CASY公司)记录藻类密度,在生长指数后期进行缢蛏投喂。
1.2缢蛏稚贝的培养
缢蛏苗由福建省紫泥甘文育苗场提供。缢蛏浮游幼虫附着后在连续充空气的水泥池中进行培养。每天投喂足量的初始混合藻饵料(角毛藻、球等鞭金藻、青岛大扁藻、云微藻、假微型海链藻、威氏海链藻等多种藻等比例混合),藻类细胞密度保持为80~100 cell/μL。水泥池底铺高温灭菌、经200目(直径75 μm)尼龙筛绢过滤的干净海泥(厚度大于1 mm),自然海水(盐度10)经沙滤后引入水泥池。稚贝在18℃~26℃下培养12 d后,取出少量于干净海水中饥饿处理24 h后用蒸馏水洗净,冷藏为空胃对照组(即实验前样品);剩余缢蛏饥饿处理12 h(排空胃中食物),进行饵料投喂实验[13, 20]。
1.3缢蛏稚贝饵料投喂实验
将饥饿处理12 h的缢蛏分为10组,每组3平行。每平行称取湿重1 g的缢蛏稚贝均匀撒入28 cm×15 cm×10 cm (长×宽×高)的白色塑料盆中,盆中海泥和海水条件与上述水泥池相同。每实验盆按实验要求投入足量单一微藻饵料或两两等比例混合后的混合微藻饵料。实验盆微藻细胞密度均为100~120 cell/μL,每4~5 d更新海水和海泥,在自然温度(18℃~26℃)下培养12 d后于干净海水中饥饿处理24 h,蒸馏水洗净缢蛏稚贝后收集冷藏样品。
1.4缢蛏脂质的提取和GC-MS检测
1.4.1脂肪酸和甾醇的提取
用冷冻干燥机(美国LABCONCO公司)干燥缢蛏样品去除水分,称取约30 mg干样研磨成粉末,参考改进的Bligh-Dyer法提取总脂[14,15]。提取总脂后加入2 mL 5%~6%氢氧化钾甲醇水(体积比为4∶1),充氮气1 min,密封后60 ℃水浴皂化2 h,冷却后用氯仿和正己烷(体积比为1∶4)6 mL分3次进行提取,往提取液中加入3 g无水硫酸钠,静置吸水12 h,将液体用旋转蒸发仪真空干燥,加入100 μL过量BSTFA,密封后60℃水浴1 h。氮气吹干后用色谱纯正己烷(美国TEDIA公司)复溶解样品,进样进行GC-MS分析。实验中其他试剂均为国产分析纯试剂。
1.4.2GC-MS检测条件
采用QP2010气相色谱-质谱分析仪,带AOC-20自动进样器(日本SHIMADZU公司),30 m×0.25 mm×0.25 btm SPB-50色谱柱(美国SUPELCO公司);标准品购自美国SIGMA-ALDRICH公司。GC采用分流进样模式,进样口温度250 ℃,载气为高纯氦,柱流速0.81 mL/min,柱前压73.0 kPa,柱起始温度150 ℃,保持3.5 min,以20 ℃/min升至200 ℃,保持5 min,以5 ℃/min升至280 ℃后,保持30 min。MS用电子轰击源(electron impact, EI)分析,电子能量70 eV,离子源温度200 ℃,接口温度250 ℃,选择全程离子碎片扫描(SCAN)模式,质量扫描范围为50~600 m/z。
1.5数据的处理与分析
根据GC-EIMS-TIC中各组分的离子碎片质量图谱,通过对NIST库和WILEY库检索并参考脂肪酸和甾醇标准谱图进行鉴定[16]。用面积归一法计算脂肪酸和甾醇组分的百分含量。不同饵料微藻投喂下脂肪酸和甾醇组成的数据用SPSS 12.0进行差异显著性分析,显著性水平P<0.05。
2结果与分析
2.1不同饵料对缢蛏脂肪酸的影响
实验共检测到16种脂肪酸,结果见表1。可以看出,不同微藻的单种饵料和混合饵料所投喂的缢蛏稚贝在脂肪酸种类数量上存在差异;作为脂肪酸组分的单不饱和脂肪酸(MUFA)、多不饱和脂肪酸(PUFA)、饱和脂肪酸在含量上也存在明显差异。实验组的不饱和脂肪酸(UFA)百分含量均明显高于饱和脂肪酸(SFA),空胃对照组的UFA百分含量略低于SFA。脂肪含量最丰富的应属畜肉类,但畜肉脂肪多为SFA,而SFA含量过高则会导致动脉粥样硬化[17],贝类食品的脂质与陆生动植物的脂质相比,具有组成复杂、PUFA含量较高的优势[18,19]。
通过徐继林[20]、杨帆[21]等人的研究,引用其对微藻脂肪酸的研究结果(如表3),本实验所用4种微藻饵料在脂肪酸种类和含量上均有差异,其中以金藻所含UFA含量最高(75.8%),角毛藻次之(75.2%)。此外,投喂不同饵料12 d后,发现缢蛏稚贝的存活率存在差异,角毛藻和金藻投喂组的空壳较少。对缢蛏稚贝进行脂质分析,发现针对脂肪酸种类丰富度,空胃组检测出10种脂肪酸;饵料投喂的实验组脂肪酸种类均多于空胃组,按表1顺序检测到的种类分别是12种、13种、10种、11种及15种、10种、11种、13种、14种、11种,其中,缢蛏脂肪酸种类丰富度最高的饵料分别是:角毛藻+金藻(15种)、金藻+威氏海链藻(14种)、金藻+假微型海链藻(13种)、金藻(13种)等。其次,针对UFA和SFA百分含量高低,空胃组与实验组呈现了明显差异,仅空胃组SFA(51.40%)含量高于UFA(48.60%),其余微藻饵料投喂组则是UFA含量均高于SFA。其中,将表1所有微藻饵料投喂组的UFA含量从高到低排列:角毛藻+金藻(70.49%)、金藻+假微型海链藻(69.75%)、角毛藻+威氏海链藻(66.85%)、金藻+威氏海链藻(65.31%)等。UFA分为MUFA和PUFA。空胃组MUFA百分含量为19.39%,角毛藻+威氏海链藻混合饵料的缢蛏投喂组MUFA百分含量最高且高达42.32%,明显高于其它微藻饵料投喂组(19.39%~36.59%)。角毛藻+威氏海链藻混合饵料投喂组的MUFA中,油酸(C18:1)的含量明显高于其它实验组微藻饵料(20.64%)。PUFA根据双键的位置和功能分为ω-3与ω-6系列。ω-3系列中最常见的是廿碳五烯酸(EPA)和廿二碳六烯酸(DHA),ω-3系列的PUFAs具有一定抑制肿瘤生长和转移的功能[22, 23];ω-6系列以花生四烯酸(AA)最常见,也有研究发现,AA产生的自由基最多,相比于EPA和DHA,AA具有更高的抗肿瘤活性[24]。本实验结果显示,AA的含量以角毛藻的单种饵料和含角毛藻的混合饵料投喂组最高(4.60%~6.69%)。空胃组只检测到EPA(16.18%)的存在,将饵料投喂组按EPA+DHA的含量从高到低顺序排列,分别是:假微型海链藻(28.9%)、假微型海链藻+威氏海链藻(26.9%)、威氏海链藻(26.67%)、金藻+假微型海链藻(24.85%)等。假微型海链藻与威氏海链藻的缢蛏投喂组EPA+DHA百分含量较高,这很可能与这两种海链藻自身EPA和DHA含量均高于角毛藻和金藻有关[21]。总而言之,将所有饵料投喂组的PUFA百分含量从高到低排序,分别是:金藻+假微型海链藻(42.64%)、角毛藻+金藻(39.84%)、金藻+威氏海链藻(33.81%)及假微型海链藻(32.45%)等。
根据实验结果,我们认为:金藻的单种饵料和含金藻的混合饵料,在提高养殖缢蛏脂肪酸种类丰富度和缢蛏UFA含量上,效果优于其它微藻饵料实验组,其次为角毛藻和含角毛藻的混合饵料。这可能与金藻、角毛藻本身的脂肪酸组成和缢蛏稚贝对外源脂肪酸的摄入与转化机制相关;此外,我们还认为金藻的混合饵料在提高缢蛏稚贝PUFA含量上,效果优于其它微藻饵料实验组。金藻在提高缢蛏稚贝脂肪酸营养组成上饵料效果更优,这与马斌[13]、杨帆[21]等人对影响缢蛏生长速度及存活率的饵料微藻的研究结果是具有相似性的。
本研究还发现C20:2和C20:3仅在金藻的单种饵料和混合饵料的投喂组被检测到;C21:0仅在金藻的单种饵料和混合饵料的投喂组被检测到(角毛藻和金藻除外)。除了角毛藻单种饵料与角+金混合饵料的投喂组以外,C16:2仅在威氏海链藻的单种饵料和混合饵料的缢蛏投喂组被检测到。这现象在一定程度上与饵料微藻的脂质组成相关[20,21]。关于C16:2、C20:2、C20:3和C21:0的作用与意义,我们不在此讨论。
2.2不同饵料对缢蛏甾醇的影响
甾醇为真核生物细胞膜基本组成成分,控制生物膜的流动性和渗透性,在某些植物中,甾醇还具有细胞信号传递、细胞酶活性调节等功能[25]。本研究共检测出了7种甾醇(表2),分别为胆甾-5-烯-3β-醇(胆甾醇)C27:1(Δ5)、24-甲基-胆甾-5, 22-二烯醇(菜子甾醇)C28:2(Δ5,22/24Me)、24-甲基-胆甾-5-烯醇(菜油甾醇)C28:1(Δ5/24Me)、4, 4-二甲基-胆甾-7, 24-二烯醇C29:2(Δ7, 24/4, 4diMe)、24-乙基-胆甾-5, 22-二烯醇(豆甾醇)C29:2(Δ5, 22/24Ethy)、24-乙基-胆甾醇(谷甾醇)C29:1(Δ5/24Ethyl)和24-乙基-胆甾-5, 24(28)E-二烯醇(岩藻甾醇)C29:2(Δ5, 24(28)E/24Ethy)。
表2显示,空胃组、角毛藻的单种饵料和含角毛藻的混合饵料投喂组、假微型海链藻的单种饵料和含假微型海链藻的混合饵料投喂组、威氏海链藻的单种饵料和含威氏海链藻的混合饵料投喂组中,均以C27:1(Δ5)百分含量最高(42.20%~91.35%)。金藻的单种饵料和含金藻的混合饵料投喂组(角毛藻和金藻混合饵料除外)未呈现此现象,且均以C28:2(Δ5,22/24Me)为含量最高的甾醇(52.93%~79.64%),此现象可能与金藻中甾醇含量的特殊性相关[16]。研究表明,菜子甾醇可以在胆甾醇缺乏时为双壳类生物提供良好的甾醇类营养物质[26]。金藻的单种饵料和含金藻的混合饵料投喂的缢蛏稚贝胆甾醇含量不高且富含菜子甾醇,从缢蛏甾醇的营养价值考虑,我们认为金藻的单种饵料和混合饵料(角毛藻和金藻混合饵料除外)的饵料效果最佳。
3结论
就脂肪酸而言,金藻的混合藻饵料的缢蛏投喂组脂肪酸种类最丰富(13~15种);金藻的混合藻饵料投喂组比其它微藻饵料投喂组富含对人体有益的UFA,尤其是PUFA含量最高。就甾醇而言,金藻的单种饵料及其混合饵料投喂组的缢蛏,胆甾醇含量不高且富含菜子甾醇,在甾醇整体营养价值更优。综上,本研究认为金藻的单种饵料及其混合饵料在对缢蛏稚贝的脂质影响上更具有营养价值,在改善缢蛏稚贝脂质组成的养殖上更有利用价值。
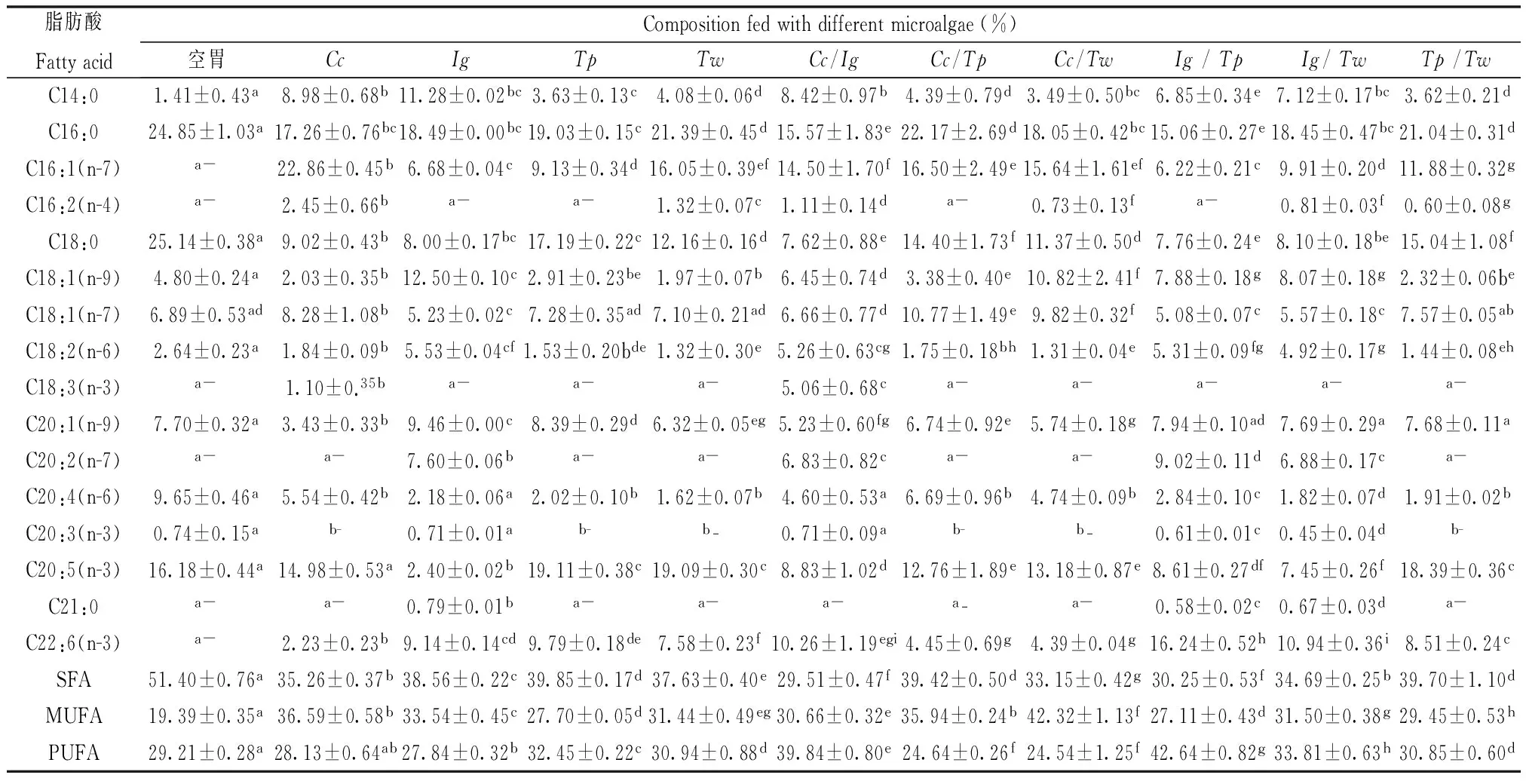
表1 4种单藻饵料及其混合饵料投喂的缢蛏稚贝脂肪酸组成
表中数据均以平均值±标准偏差表示;“-”表示该物质未检测到;同一行数据共有相同的字母上标,表示差异性不显著(P>0.05)。SFA:饱和脂肪酸;MUFA:单不饱和脂肪酸;PUFA:多不饱和脂肪酸

表2 4种单藻饵料及其混合饵料投喂的缢蛏稚贝甾醇组成
表中数据均以平均值±标准偏差表示;“-”表示该物质未检测到;同一行数据共有相同的字母上标,表示差异性不显著(P>0.05)

表3 4种微藻饵料的脂肪酸组成[20, 21]
SFA:饱和脂肪酸;MUFA:单不饱和脂肪酸;PUFA:多不饱和脂肪酸
参考文献:
[1]HAGGARTY P. Fatty acid supply to the human fetus[J].Annual Review of Nutrition, 2010, 30(1), 237-255.
[2]RYAN A S, ASYWOOD J D, GAUTIER S, et al. Effects of long-chain polyunsaturated fatty acid supplementation on neurodevelopment in childhood: a review of human studies[J].Prostaglandins, Leukotrienes and Essential Fatty Acids, 2010, 82(4-6):305-314.
[3]EADY S, WALLACE A, WILLIS J, et al. Consumption of a plant sterol-based spread derived from rice bran oil is effective at reducing plasma lipid levels in mildly hypercholesterolaemic individuals[J].British Journal of Nutrition, 2011, 105(12):1808-1818.
[4]WEINGARTNER O, ULRICH C, ISMAL K, et al. Differential effects on inhibition of cholesterol absorption by plant stanol and plant sterol esters in apoE2/2 mice[J].Cardiovascular Research, 2011, 90(3):484-492.
[5]LE GRAND F, KRAFFE E, MARTY Y, et al. Membrane phospholipid composition of hemocytes in the Pacific oysterCrassostreagigas and the Manila clamRuditapesphilippinarum[J].Comparative Biochemist and Physiology Part A: Molecular and Integrative Physiology, 2011, 159(4):383-391.
[6]COPEMAN L A, PARRISH C C. Lipid classes, fatty acids, and sterols in seafood from Gilbert Bay, southern Labrador[J]. Journal of Agriculture and Food Chemistry, 2004, 52(15):4872-4881.
[7]林叶, 苏秀榕, 孙蓓, 等.不同种群缢蛏氨基酸及脂肪酸比较研究[J]. 食品科学, 2006, 27(12):675-678.
[8]赵艳景, 胡虹, 王颖.缢蛏多糖的提取及抗氧化作用研究[J]. 水产科学, 2010, 29(2): 91-94.
[9]林文东, 缢蛏水提物的体外抗氧化活性研究[J]. 亚太传统医药, 2011, 1(1): 25-26.
[10]张永娟, 吕学军, 张大勇.缢蛏水解物的抗疲劳作用[J]. 中国生化药物杂志, 2011, 32(2): 133-136.
[11]SOUDANT P, SANIES M V, QUERE C, et al. The use of lipid emulsions for sterol supplementation of spat of the Pacific oyster,Crassostreagigas[J].Aquaculture, 2000, 184(99):315-326.
[12]NAPOLITANO G E, ACKMAN R G, SILVA- SERRA M A. Incorporation of dietary sterols by the sea scallopPlacopectenmagellanictrsGmelin fed on microalgae[J].Marine Biology, 1993, 117(4): 647-654.
[13]马斌, 周海波, 徐继林, 等.缢蛏稚贝优质微藻饵料的筛选[J]. 宁波大学学报, 2011, 24(4):5-9.
[14]BLIGH E G, DYER W J. A rapid method lipid extraction and purification[J].Canadian Journal of Biochemistry and Physiology, 1959, 37(8):911-917.
[15]李海英, 徐继林, 严小军.16种贝类脂肪酸和甾醇组成分析[J].宁波大学学报, 2009, 22(1):48-54.
[16]徐继林, 严小军, 周成旭, 等.甾醇在海洋微藻中的分布研究[J].海洋学报, 2007, 29(6):93-100.
[17]葛可佑.中国营养科学全书[M].北京:人民卫生出版社, 2004:56-66.
[18]王炜, 张伟敏.单不饱和脂肪酸的功能特性[J].中国食物与营养, 2005(4):44-46.
[19]张永普, 贾守菊, 应雪萍.不同种群泥蚶肉营养成分的比较研究[J], 海洋湖沼通报, 2002(2):33-38.
[20]XU J L, ZHOU H B, YAN X J, et al. Effect of unialgal diets on the composition of fatty acids and sterols in Juvenile Ark ShellTegillarcagranosaLinnaeus[J].Journal of Agriculture and Food Chemistry, 2012, 60(15):3973-3980.
[21]YANG F, CHEN S, MIAO Z, et al. The effect of several microalgae isolated from East China Sea on growth and survival rate of postset juveniles of razor clam,Sinonovaculaconstricta(Lamarck, 1818)[OL]. Aquaculture Nutrition, 2015, dio:10.1111.
[22]尹勇, 詹文华, 彭俊生.ω-3多不饱和脂肪酸抗肿瘤机制的研究进展[J].国际外科学杂志, 2004, 31(4):237-240.
[23]ROSE D P, CONNOLLY J M. Effects of fatty acids and inhibitors of eicosanoid synthesis on the growth of human breast cancer cell line in culture[J].Cancer Research, 1990, 50(22):7139-7144.
[24]BEGIN M E, ELLS G, DAS U N, et al. Differential kill of human carcinoma cells supplemented with n-3 and n-6 polyunsaturated fatty acids[J].Journal of the National Cancer Institute, 1986, 77(5):1053-1062.
[25]PIIRONEN V, LINDSAY D G, MIETTINEN T A, et al. Plant sterols: biosynthesis, biological function and their importance to human nutrition[J].Journal of the Science of Food and Agriculture, 2000, 80 (7): 939- 966.
[26]SOUDANT P, MARTY Y, MOAL J, et al. Effect of food fatty acid and sterol quality onPectenmaximusgonad composition and reproduction process[J].Aquaculture, 1996, 143(3):361-378.
Effect of several diets on the composition of fatty acids and sterols in juvenile Sinonovacula constricta
ZHOU Lv1, RAN Zhao-shou1, YU Shan-shan1, ZHOU Hai-bo2, XU Ji-lin1, YAN Xiao-jun1
(1. Key Laboratory of Applied Marine Biotechnology, School of Marine Science, Ningbo University, Ningbo 315211;2. Zhibao Fishery Science Co., Ltd. in Fujian Province, Zhangzhou 363503, China)
AbstractIt is investigated that effects of the four microalgae (Chaetoceros calcitrans, Isochrysis galbana, Thalassiosira pseudonana and Thalassiosira weissflogii) and their mixture, which commonly used as S. constricta rearing diets, on the composition of fatty acids and sterols in juvenile S. constricta. The fatty acid and sterol composition of juveniles were analyzed by GC-MS. The results showed that different diets have effect on fatty acid and sterol compositions in juveniles, and 16 fatty acids and 7 sterols have been detected. C16:2 detected in T. weissflogii and its mix. Juveniles fed with I. galbana mix was abundant in the species of fatty acids(13-15) and C28:1(Δ5/24Me) (Brassicasterol). Besides, the PUFA percentage content of Juveniles fed with I. galbana mix was higher than that of others. Above all, we choose I. galbana mix as optimal diet because they made lipids of S. constricta juveniles more nitrification.
Key wordsmicroalgae; S. constricta; fatty acid; sterol; GC-MS
收稿日期:2015-12-10;修回日期:2016-01-04
基金项目:国家自然科学基金(31172448);教育部博士点基金优先发展领域项目(20133305130001);浙江省公益性项目(2014C32081);浙江自然科学基金项目(LY15C190004);宁波市科技攻关项目(2013C10014,2014C10005)
作者简介:周 率,硕士研究生,从事海洋生物化学研究,E-mail: zhoulv9597@126.com; 通信作者:徐继林,研究员,博士生导师,研究方向为海洋生物的环境生态和养殖生态学研究,E-mail: xujilin@nbu.edu.cn。
中图分类号Q959.9
文献标识码A
文章编号2095-1736(2016)03-0052-05
doi∶10.3969/j.issn.2095-1736.2016.03.052
