内蒙古荒漠草原短花针茅叶片功能性状对不同草地经营方式的响应
2016-06-28刘文亭卫智军吕世杰白玉婷肖嘉圃代景忠孙世贤
刘文亭,卫智军*,吕世杰,白玉婷,肖嘉圃,代景忠,孙世贤
1. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;2. 内蒙古农业大学理学院,内蒙古 呼和浩特 010018;3. 中国农业科学院草原研究所,内蒙古 呼和浩特 010010
内蒙古荒漠草原短花针茅叶片功能性状对不同草地经营方式的响应
刘文亭1,卫智军1*,吕世杰2,白玉婷1,肖嘉圃1,代景忠1,孙世贤3
1. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;2. 内蒙古农业大学理学院,内蒙古 呼和浩特 010018;3. 中国农业科学院草原研究所,内蒙古 呼和浩特 010010
摘要:植物叶片功能性状对不同草地经营方式的适应是一种有效地生态对策。短花针茅(Stipa breviflora)是亚洲中部暖温性荒漠草原的主要建群种,通过调查生长良好的不同龄级短花针茅叶性状,试图揭示荒漠草地利用模式与植物生态响应机制。该研究测量自然叶高(LNH,leaf natural height)、叶长(LL,leaf length)、叶鲜质量(LFW,leaf fresh weight)、叶干质量(LDW,leaf dry weight)、自然叶宽(LW,leaf width)、叶缘距(LED,leaf edge distance)、叶直立度(leaf erection index,LEI)、叶卷曲度(leaf rolling index,LRI)和叶干物质含量(leaf dry matter content,LDMC)等叶性状指标。结果表明,(1)短花针茅的LNH、LL、LFW、LDW、LW、LED等叶性状在灌溉草地几乎都显著大于放牧草地和围封草地。(2)LEI、LFW为生境干扰响应的敏感性状;LW、LDMC为生境干扰响应的惰性性状。(3)Pearson相关分析结果可知,放牧对短花针茅叶性状影响最大。研究认为,短花针茅叶片功能性状对不同草地经营方式的的适应对策,是通过对不同性状的权衡,充分利用环境资源来完成的。
关键词:草地经营方式;权衡;短花针茅;叶片功能性状
引用格式:刘文亭, 卫智军, 吕世杰, 白玉婷, 肖嘉圃, 代景忠, 孙世贤. 内蒙古荒漠草原短花针茅叶片功能性状对不同草地经营方式的响应[J]. 生态环境学报, 2016, 25(3): 385-392.
LIU Wenting, WEI Zhijun, LV Shijie, BAI Yuting, XIAO Jiapu, DAI Jingzhong, SUN Shixian. Response of Grassland Using Modes to Leaf Trait of Stipa breviflora in Desert Steppe of Inner Mongolia [J]. Ecology and Environmental Sciences, 2016, 25(3): 385-392.
叶片是植物获取资源的重要器官之一,对生境变化敏感且具有可塑性(Shipley et al.,2003;Vile et al.,2005)。叶片功能性状不仅解释了植物在不同环境梯度下的分布特征以及对环境的高度适应能力,体现其在复杂生境下的自我调控能力和适应机理,同时还能一定程度地预测生态系统对环境及对生态系统过程的影响(王常顺等,2015)。短花针茅是亚洲中部暖温性荒漠草原的主要建群种,研究短花针茅叶片功能性状对不同草地经营方式的响应,对于揭示荒漠草地资源利用与植物生态响应机制有重要意义。
近年来,植物性状对环境的响应关系逐渐引起了生态学者的关注(Bullock et al.,2001;Bernard-Verdier et al.,2012;李西良等,2015),它反映植物在对环境适应过程中,内部生理与外部表型不同功能之间的权衡,是植物对生境生存策略的重要体现(Mooney et al.,2010;De Kroon et al.,2005)。关于生源要素、气候、草食动物采食等环境因子对植物功能性状的驱动机制,近年研究颇多,但主要集中在研究植物性状对气候环境梯度的适应对策与规律(Van Kleunen et al.,2010;Lindborg et al.,2012)。Díaz et al.(2007)通过整合全球不同地区观测的研究结果,对草地植物的生活史等7个主要性状进行分析,发现相比于多年生、高大、直立等特征的植物,放牧利用对具有一年生、矮小、匍匐等特征的植物作用更强,功能群之间呈现差异化响应特征。但迄今为止,关于草地植物性状可塑性变化的报道仍显不足(Stahlheber et al.,2013),特别是关于不同性状的敏感度差异成为限制解析草原植物生物学过程的重要瓶颈。
此外,叶片随叶龄增长的投资和物质分配亦是植物进化的重要途径之一(黄海侠等,2013)。例如具有较长生活周期的常绿植物叶片,其所处的发育阶段的差异、形成的年份、经历的环境条件改变,使其性状特征受资源分配模式和叶龄变化产生差别(Doust,1980;Reich et al.,1991)。在叶片生长早期,叶片需投入较多的生物量以优化同化器官,进而最大程度地获取光资源,如扩展平均叶面积(mean leaf area,MLA)(Sultan,2000),从而可增大光截获和碳收益方面的竞争优势,以补偿其自身建成的消耗(Williams et al.,1987;Wright et al.,2004)。相对于生长后期的叶片,生长早期的叶片在单位生物量投入下其面积的增大程度较大。相反,叶片干物质含量(leaf dry matter content,LDMC鲜重与干重的比率)在生长前期则较小,反映了发育初期叶片一般含水率较高,代谢活动活跃(Sims et al.,1992)。但随着叶片表面积的增大,植物会经历更多更为复杂的生存威胁,因此需要在抗机械胁迫和结构建造等方面投入更多的生物量,从而维持叶片的存活时间(Witkowski et al.,1991;Wilson et al.,1999;Gratani et al.,2000;Westoby et al.,2002)。具体而言,老叶组织密度和厚度的增大可直接导致叶片光合能力的衰退(Field et al.,1983;He et al.,2009),叶片通过继续增大光合面积所取得的边际收益已甚小,导致叶面积在叶片生长后期的增大空间有限,其大小的变动不大。短花针茅(Stipa breviflora)为多年生丛生型禾草,叶片呈针状并向内卷曲,极少数纵折、扁平,之前的研究结果是否同样适用于短花针茅?
于内蒙古短花针茅荒漠草原选择了3种经营方式的草地进行取样(灌溉草地、放牧草地、围封草地),通过测量3种不同经营方式草地短花针茅的自然叶高(LNH,leaf natural height)、叶长(LL,leaf length)、叶鲜重(LFW,leaf fresh weight)、叶干重(LDW,leaf dry weight)、自然叶宽(LW,leaf width)、叶缘距(LED,leaf edge distance)、叶直立度(leaf erection index,LEI)、叶卷曲度(leaf rolling index,LRI)和叶干物质含量(leaf dry matter content,LDMC),比较不同龄级的短花针茅叶性状,拟探讨以下问题:(1)不同龄级短花针茅叶性状对3种草地经营方式的响应特征;(2)短花针茅叶性状对3种草地经营方式的响应规律。
1 材料与方法
1.1 研究区概况与样地描述
试验区位于内蒙古锡林郭勒盟苏尼特右旗朱日和镇(112°47′E,42°15′N)。地形平坦,有明显的钙积层,分布在10~35 cm土层中;土壤为淡栗钙土,腐殖质层厚5~10 cm。年平均降水量177.2 mm,而60%~80%的降水集中在牧草生长旺盛的7—9月。蒸发量2500 mm,大多数年受到不同程度的干旱威胁。群落种的饱和度平均10种/m2左右,草群高度10~20 cm,总盖度15%~25%。群落结构简单。以短花针茅为建群种,无芒隐子草(Cleistogenes songorica)为亚建群种,伴生狭叶锦鸡儿(Caragana stenophylla)、木地肤(Kochia prostrata)、天门冬(Asparagus gobicus)等。
本研究在苏尼特右旗荒漠草原进行,根据当地牧民草地经营方式的不同,分别在以下3种草地取样:灌溉草地、放牧草地、围封草地。围封草地位于朱日和牧区,样地位置远离牧户,历史上一直很少有放牧活动,几乎不受人类活动的干扰,2009年设置围栏,短花针茅在群落中占绝对优势。放牧草地自2000年起开始,草场界限清楚,草地经营水平中等,在当地具有代表性,是成熟的短花针茅草原,主要放牧“苏尼特羊”,载畜率为0.96只羊/(hm2·a-1)。灌溉草地自2012年起围封实施灌溉,围封前与原生样地一样,采用嘉利DPP-325电动平移式大型喷灌设施,灌溉周期为每年的5月上旬—9月下旬,10月份牧户打草以供家畜越冬饲用。
1.2 取样与测量
在自然植物群落中,生境及生长年龄的不确定性,导致短花针茅个体存在较大差异,但短花针茅的独特性也为研究过程提供了另一种思路:采用径级结构代替龄级结构来分析针茅叶性状。尽管径级和龄级不同,但在相同的环境条件下,同一物种的径级和龄级对环境等条件的反应规律基本是一致的。参照乔木相关研究中对林冠层林木径级结构的划分标准,同时结合草地短花针茅个体生长实际情况,将短花针茅基径大小近似划分为龄级Ⅰ(basal diameter<5 cm)、龄级Ⅱ(5 cm
于2014年8月草原生长高峰期,短花针茅停止生物量积累时进行野外取样工作。在3种不同草地经营方式的草地内,分别调查生长良好的大丛短花针茅(基部丛径>10 cm)、中丛短花针茅(基部丛径5~10 cm)、小丛短花针茅(基部丛径<5 cm)各50丛,每丛植株均从不同方向上测量10片叶片的自然叶宽(叶片自然状态的宽度LW)、叶缘距(将叶片展平时的宽度,LED)、自然叶高(自然状态下叶片最高点距离地面的垂直高度LNH)、叶长(将叶片拉直时的长度LL),之后齐地面刈割该丛短花针茅,共采集短花针茅450丛,分别保存在自封袋中,记录编号带回实验室。
于12 h内用去离子水快速冲洗干净,吸干表面水分,称取采集的短花针茅湿重(LFW),待完成后,将新鲜材料置于105 ℃烘箱中杀青10 min,将叶片置于75 ℃干燥箱中烘干至恒重,称量样品干重(LDW)。通过公式(1~3)计算出3种草地内不同基径短花针茅叶片的卷曲度、直立度、干物质含量:

1.3 数据分析
采用方差分析(GLM)对灌溉草地(M)、放牧草地(G)、围封草地(UG)3个龄级的短花针茅叶性状进行差异显著性分析,并进行Duncun多重比较,数据以平均值±标准误差表示。叶性状对草地经营的响应程度用可塑性指数表示,M、G样地中某一叶性状的可塑性指数(PI)为:UG样地数值减去M、G样地数值的绝对值,除以(M和UG)、(G和UG)样地数值中的最大值。用变异系数(CV)表征植物性状的变异程度,计算方法为标准差除以平均值。采用线性回归方程拟合短花针茅叶性状对草地经营模式的可塑性程度(PI)与其变异性(CV)之间的关系,分析植物性状变异性对环境的响应。利用Pearson相关法分析各叶性状之间的相关性,在0.05和0.01水平下进行显著性检验。
2 结果
2.1 异龄叶性状对草地经营模式的响应
叶片是植物体重要的功能器官,从草地经营模式看(图1),短花针茅的LNH、LL、LFW、LDW、LW、LED等叶性状在M几乎都显著大于G和UG (P<0.05),其中叶长与叶宽具有相同的表现型,放牧并没有显著改变其叶性状。从叶龄结构看,龄级Ⅲ的LFW值是龄级Ⅰ的13倍,是龄级Ⅱ的1.4倍,龄级Ⅲ的LDW值是龄级Ⅰ的12倍,是龄级Ⅱ的1.7倍;除此之外,LNH、LL、LW、LED等叶性状数值变化较小。
进一步分析(图2),在LEI方面,不同草地利用的龄级Ⅰ与龄级Ⅱ无显著差异,龄级Ⅲ中UG显著大于G;在LDMC方面,G和UG均显著大于M,其中,龄级Ⅱ与龄级Ⅲ下UG显著大于G,而龄级Ⅰ刚好相反;在LRI方面,G和UG均显著大于M,UG与G无显著性差异。
2.2 叶性状可塑性变化的敏感度
以围封草地为参考系,分析各叶性状的可塑性指数,图3对M与G草地短花针茅9种叶性状的可塑性指数大小进行了排序。从整体分析,M的可塑性指数大于G地,各性状可塑性指数大小排序规律基本一致;在M与G草地中,LEI、LFW的可塑性幅度较大(PI>0.45),为生境干扰响应的敏感性状;LW、LDMC的可塑性幅度较小(PI<0.20),为生境干扰响应的惰性性状。在各种叶性状指标中(图4),LEI、LFW的变异系数幅度较大(CV>0.80),LW、LDMC的变异幅度较小(CV<0.20),即对生境变化敏感的性状具有较大的变异性。短花针茅叶性状对草地方式的响应程度(PI)与其变异性(CV)之间符合线性方程(y=0.460x+0.117)拟合关系(R2=0.810,P<0.001)(图5)。
2.3 短花针茅叶性状的关联
由3种不同草地的Pearson相关分析结果(图6)可知,在灌溉草地、放牧草地和围封草地各自的叶性状对中,正关联叶性状对所占百分比分别为60.11%、70.09%、47.01%,负关联叶性状对所占百分比分别为39.89%、29.91%、52.99%。放牧草地正相关叶性状对最多,灌溉草地其次,围封草地最少,而围封草地短花针茅叶性状对的正相关与负相关百分比较接近50%,这表明相对其他草地而言,放牧对短花针茅叶性状影响较大。
相关性达显著或极显著的叶性状对及其关联测定结果(图6),那些关联不显著的叶性状对的联结性质及关联程度也因草地利用的不同而有差异,表明同一地区不同草地利用方式的改变使同一叶性状对的联结性质或关联程度发生变化。LDW 与LRI正相关,且在放牧地呈显著正相关;LED 与LEI几乎都呈显著正相关(P<0.05);LNH与LDMC呈极显著正相关(P<0.01);LNH与LRI正相关;LNH与LW在同龄叶和异龄叶中都呈正相关。
3 讨论
叶片在不同生长阶段的资源分配方式在很大程度上反映了植物的生活史特征(Weiner,2004),短花针茅是多年生丛生型根茎禾草,叶的生活周期长,生活史策略复杂,异龄叶间的关系可通过叶片性状在短花针茅不同叶龄间的关联性体现。LNH与LW在同龄叶和异龄叶中都呈正相关,即LNH增高,LW也同时增宽,而在同龄叶中,仅有LNH与LDMC呈极显著正相关(P<0.01),这可能是因为短花针茅在环境变化过程中分化为敏感性状和惰性性状,而LW是惰性性状,这暗示了短花针茅在生长期将叶片尽可能地拉至最长,可有效提高植物的生存效率。此外,M地叶宽与叶长具有较大值(图1),LDMC值较小(图2),说明M地叶片具有更优的光资源获取能力(Field et al.,1983;Kitajima et al.,1997)和较为活跃的代谢水平(Sims et al.,1992)。相对于G和UG,M将叶片面积展开至最大,以获取最多的光资源和达到最高的碳生产(Williams et al.,1987;Sultan,2000)。而在UG和G,外界环境因子胁迫作用强,叶片光合能力衰退(Field et al.,1983;Kitajima et al.,1997;Bleecker,1998;Ono et al.,2001)。这种情况下,再通过增大叶长与叶宽从而提升光合作用的收益已不显著,这时植物的生存策略可能是投入较多营养在组织密度上,以抵御环境胁迫和病虫害(Reich et al.,1998),如增大丛生基径、长出更多的叶子。
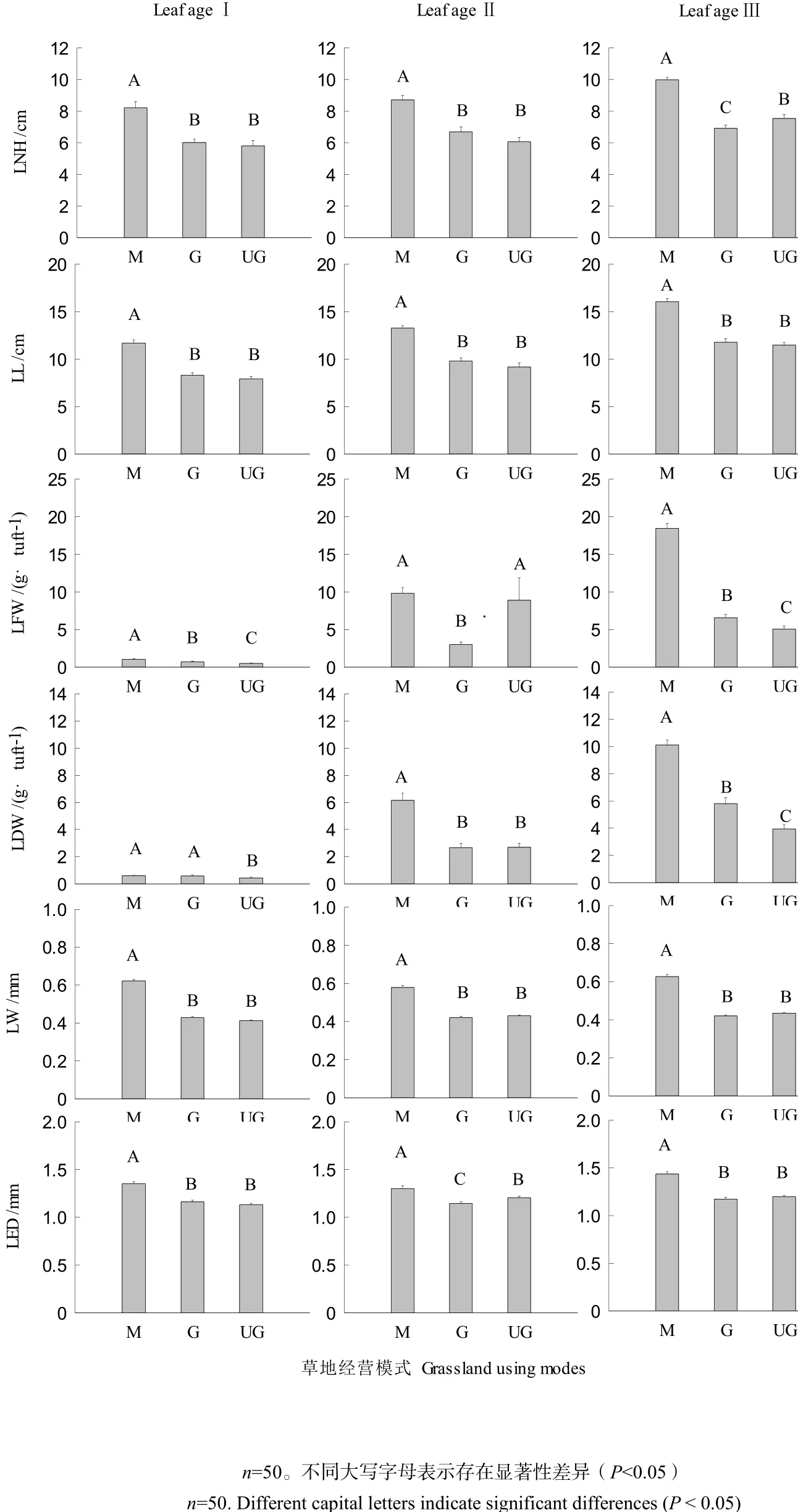
图1 短花针茅自然叶高(LNH,leaf natural height)、叶长(LL,leaf length)、叶鲜重(LFW,leaf fresh weight)、叶干重(LDW,leaf dry weight)、自然叶宽(LW,leaf width)、叶缘距(LED,leaf edge distance)对草地经营方式的响应(平均值±标准误差)Fig. 1 Effects of grassland using modes to leaf trait (LNH, LL, LFW, LDW, LW, LED) of Stipa breviflora (mean ± SE). LNH, leaf natural height; LL, leaf length; LFW, leaf fresh weight; LDW, leaf dry weight; LW, leaf width; LED, leaf edge distance

图2 短花针茅叶直立度(leaf erection index,LEI)、叶卷曲度(leaf rolling index,LRI)和叶干物质含量(leaf dry matter content,LDMC)对草地经营方式的响应(平均值±标准误差)Fig. 2 Effects of grassland using modes to leaf trait (LEI, LDMC, LRI) of Stipa breviflora (mean ± SE). LEI, leaf erection index; LDMC, leaf dry matter content; LRI, leaf rolling index
在短花针茅与生境协同适应及进化过程中,叶片呈针状并向内卷曲,极少数纵折、扁平。研究表明,植物叶片卷曲现象由叶片细胞膨压降低所引起(Hsiao et al.,1984),是内部水势和渗透调节结果的外部形式的综合表现,通常在干旱情况下,卷叶通过改变其叶片的电导率和有效叶面积来减少其拦截的辐射量及降低蒸腾作用,进而有效防止水分的丧失(Kadioglu et al.,2007;Price et al.,1997),而这一点在本研究中得到了进一步印证,G和UG 的LRI均显著大于M。水分的增加显著地提高了短花针茅的的LNH、LL、LFW、LDW、LW、LED等叶基本性状,而这一系列的改变直接影响了短花针茅叶表现型和对资源的获取利用。说明水分直接影响短花针茅的生物学属性,表明了干旱胁迫是限制荒漠草地植物生长的一个重要因素,植物在异质生境下的性状特征的变异是植物克服环境胁迫的重要途径之一。

图3 灌溉草地、放牧草地的短花针茅不同叶性状可塑性指数(PI)排序Fig. 3 Sorting of Stipa breviflora variation in leaf traits plasticity index (PI) in Manual grassland (M),grazing grassland (G)

图4 灌溉草地、放牧草地、围封草地短花针茅不同叶性状变异系数排序Fig. 4 Sorting of Stipa breviflora leaf trait variable coefficient in Manual grassland (M), grazing grassland (G), ungrazing grassland (UG)

图5 短花针茅不同叶性状变异系数(CV)与可塑性指数(PI)的关系Fig. 5 Regression fitting coefficient variation (CV) and plasticity index (PI) of leaf traits in Stipa breviflora
叶作为植物的基本结构和功能单位,不仅影响植物的基本行为和功能,而且能够反映植物对不同生境的适应能力。然而,有趣的是,本文通过排序构建短花针茅性状变化谱,发现LEI是对生境干扰响应的敏感因子(图3),而这一发现似乎与LEI 在3个样地间无显著性差异(图2)这一结果自相矛盾。其实不然!多数学者认为叶性状的变异是由植物其他性状的协同变异引起,如植物高度(Ackerly et al.,1998;Cornelissen,1999)、干材密度(Pickup et al.,2005)、支撑枝条的大小(Westoby et al.,2003)和单位体积内叶稠密度的大小(Yan et al.,2012)等。叶片发生卷曲是解决叶长与叶挺两者之间矛盾的一种行之有效的方法,因为卷叶具有的最直接效应是对叶片的直化作用,叶片长度的增加往往容易导致植株叶片披垂,但卷叶性状可以提高较长叶片的挺直性,减少披垂现象的发生,卷曲度较大,可支持较长的叶片挺立不披垂。叶片的直立可以有效减少叶片的投影面积、增加群体中上层叶面积的比例、改善群体中后期的受光条件和通风状况及干物质的积累(沈年伟等,2009)。这可能是短花针茅适应干旱环境的的生存策略,即敏感性状并不一定要有十分显著的表现型,它是植物长期进化形成的多性状之间的权衡。那么,短花针茅叶性状对草地经营方式响应的敏感度的分化具有什么样的生态学含义?惰性性状、敏感性状分别在维持短花针茅植株的光合作用、营养吸收、水分利用中的生物学功能如何?由于植物在环境胁迫下具有权衡性状关系的适应策略(Mooney et al.,2010),维持叶片的惰性响应特征对充分发挥植物生产功能具有重要作用。本研究还发现,围封草地与放牧草地的叶长与叶宽无显著性差异,这可能是由于叶宽与叶长的对生境的响应程度不同导致叶片长宽比下降,叶片相对宽度增加,叶片几何形状纤细化程度缩减,可能利于提高植物水分、养分利用效率等生物学功能的发挥(Giese et al.,2011),但需进一步的实验证实。

图6 不同草地经营方式下短花针茅异龄叶性状的半矩阵Pearson相关分析。其中1、10、19,自然高度;2、11、20,叶长;3、12、21,叶鲜重;4、13、22,叶干重;5、14、23,叶宽;6、15、24,叶缘距;7、16、25,叶直立度;8、17、26,干物质含量;9、18、27,叶卷曲度。▲,极显著正相关 ;△,显著正相关;▼,极显著负相关;▽,显著负相关;+,正相关;-,负相关Fig. 6 The semi-matrix of Pearson's correlation for different age of Stipa breviflora foliar traits in Manual grassland (M), grazing grassland (G), ungrazing grassland (UG). 1, 10, 19, LNH; 2, 11, 20, LL; 3, 12, 21, LFW; 4, 13, 22, LDW; 5, 14, 23, LW; 6, 15, 24, LED; 7, 16, 25, LEI; 8, 17, 26, LDMC; 9, 18, 27, LRI.▲, Positive correlation at P = 0.01; △, Positive correlation at P = 0.05; ▼, Negative correlation at P = 0.01; ▽, Negative correlation P = 0.05; +, Non positive correlation; -, Non negative correlation
4 结论
本研究从不同荒漠草地经营方式入手,分析不同龄级短花针茅叶片自然叶高、叶长、叶鲜重、叶干重、自然叶宽、叶缘距、叶直立度、叶卷曲度和叶干物质含量。研究认为,灌溉条件下草地短花针茅叶片自然叶高、叶长、叶鲜重、叶干重、自然叶宽、叶缘距基本显著大于放牧草地和围封草地,水分条件导致短花针茅叶性状差异显著;短花针茅的叶直立度、叶鲜重为生境干扰响应的敏感性状,自然叶宽、叶干物质含量为生境干扰响应的惰性性状,即对生境变化敏感的性状具有较大的变异性;放牧对短花针茅叶性状影响最大。短花针茅叶性状对不同草地经营方式的的适应对策,体现了其在不同生境下的最优生活对策,在生态系统亚稳态下,充分利用环境资源完成其生活史。
参考文献:
ACKERLY D D, DONOGHUE M J. 1998. Leaf size, sapling allometry, and Corner's rules: phylogeny and correlated evolution in maples (Acer) [J]. The American Naturalist, 152(6): 767-791.
BERNARD-VERDIER M, NAVAS M L, VELLEND M, et al. 2012. Community assembly along a soil depth gradient: contrasting patterns of plant trait convergence and divergence in a Mediterranean rangeland [J]. Journal of Ecology, 100(6): 1422-1433.
BLEECKER A B. 1998. The evolutionary basis of leaf senescence: method to the madness? [J] Current Opinion in Plant Biology, 1(1): 73-78.
BULLOCK J M, FRANKLIN J, STEVENSON M J, et al. 2001. A plant trait analysis of responses to grazing in a long-term experiment [J]. Journal of Applied Ecology, 38(2): 253-267.
CORNELISSEN J H C. 1999. A triangular relationship between leaf size and seed size among woody species: allometry, ontogeny, ecology and taxonomy [J]. Oecologia, 118(2): 248-255.
DE KROON H, HUBER H, STUEFER J F, et al. 2005. A modular concept of phenotypic plasticity in plants [J]. The New Phytologist, 166(1): 73-82.
DÍAZ S, LAVOREL S, MCINTYRE S, et al. 2007. Plant trait responses to grazing-A global synthesis [J]. Global Change Biology, 13(2): 313-341.
DOUST J L. 1980. A comparative study of life history and resource allocation in selected Umbelliferae [J]. Biological Journal of the Linnean Society, 13(2): 139-154.
FIELD C, MOONEY H A. 1983. Leaf age and seasonal effects on light,water, and nitrogen use efficiency in a California shrub [J]. Oecologia,56(2-3): 348-355.
GIESE M, GAO Y Z, LIN S, et al. 2011. Nitrogen availability in a grazed semi-arid grassland is dominated by seasonal rainfall [J]. Plant and Soil,340(1-2): 157-167.
GRATANI L, BOMBELLI A. 2000. Correlation between leaf age and other leaf traits in three Mediterranean maquis shrub species: Quercus ilex,Phillyrea latifolia and Cistus incanus [J]. Environmental and Experimental Botany, 43(2): 141-153.
HE J S, WANG X P, FLYNN D F B, et al. 2009. Taxonomic, phylogenetic,and environmental trade-offs between leaf productivity and persistence [J]. Ecology, 90(10): 2779-2791.
HSIAO T C, O'TOOLE J C, YAMBAO E B, et al. 1984. Influence of osmotic adjustment on leaf rolling and tissue death in rice. Plant physiology [J]. 75(2): 328-341.
KITAJIMA K, MULKEY S S, WRIGHT S J. 1997. Decline of photosynthetic capacity with leaf age in relation to leaf longevities for five tropical canopy tree species [J]. American Journal of Botany, 84(5): 702-708.
KADIOGLU A, TERZI R. 2007. A dehydration avoidance mechanism: leaf rolling[J]. Botanical Review, 73(4): 290-302.
LINDBORG R, HELM A, BOMMARCO R, et al. 2012. Effect of habitat area and isolation on plant trait distribution in European forests and grasslands [J]. Ecography, 35(4): 356-363.
MOONEY K A, HALITSCHKE R, KESSLER A, et al. 2010. Evolutionary trade-offs in plants mediate the strength of trophic cascades [J]. Science, 327(5973): 1642-1644.
ONO K, NISHI Y, WATANABLE A, et al. 2001. Possible mechanisms ofadaptive leaf senescence [J]. Plant Biology, 3(3): 234-243.
PICKUP M, WESTOBY M, BASDEN A. 2005. Dry mass costs of deploying leaf area in relation to leaf size [J]. Functional Ecology,19(1): 88-97.
PRICE A H, YOUNG E M, TOMOS A D. 1997. Quantitative trait loci associated with stomatal conductance leaf rolling and heading date mapped in upland rice [J]. New phytologist, 137(1): 83-91.
REICH P B, WALTERS M B, ELLSWORTH D S, et al. 1998. Relationships of leaf dark respiration to leaf nitrogen, specific leaf area and leaf life-span: a test across biomes and functional groups [J]. Oecologia,114(4): 471-482.
REICH P B, WALTERS M B, ELLSWORTH D S. 1991. Leaf age and season influence the relationships between leaf nitrogen, leaf mass per area and photosynthesis in maple and oak trees [J]. Plant, Cell and Environment, 14(3): 251-259.
SHIPLEY B, ALMEIDA-CORTEZ J. 2003. Interspecific consistency and intraspecific variability of specific leaf area with respect to irradiance and nutrient availability [J]. Ecoscience, 10(1), 74-79.
SIMS D A, PEARCY R W. 1992. Response of leaf anatomy and photosynthetic, capacity in Alocasia macrorrhize (Araceae) to a transfer from low to high light [J]. American Journal of Botany, 79(4): 449-455.
STAHLHEBER K A, D'ANTONIO C M. 2013. Using livestock to manage plant composition: A meta-analysis of grazing in California Mediterranean grasslands [J]. Biological Conservation, 157(2): 300-308.
SULTAN S E. 2000. Phenotypic plasticity for plant development,function and life history [J]. Trends in Plant Science, 5(12): 537-542.
VAN KLEUNEN M, WEBER E, FISCHER M. 2010. A meta-analysis of trait differences between invasive and non-invasive plant species [J]. Ecology Letters, 13(2): 235-245.
VILE D, GARNIER E, SHIPLEY B, et al. 2005. Specific Leaf Area and dry Matter Content Estimate thickness in Laminar Leaves [J]. Annals of Botany, 96(6):1129-36.
WEINER J. 2004. Allocation, plasticity and allometry in plants [J]. Perspectives in Plant Ecology Evolution and Systematics, 6(4): 207-215.
WESTOBY M, FALSTER D S, MOLES A T, et al. 2002. Plant ecological strategies: some leading dimensions of variation between species [J]. Annual Review of Ecology and Systematics, 33(1): 125-159.
WESTOBY M, WRIGHT I J. 2003. The leaf size-twig size spectrum and its relationship to other important spectra of variation among species [J]. Oecologia, 135(4): 621-628.
WILLIAMS K, PERCIVAL F, MERINO J, et al. 1987. Estimation of tissue construction cost from heat of combustion and organic nitrogen content [J]. Plant, Cell & Environment, 10(9): 725-734.
WILSON P J, THOMPSON K, HODGSON J G. 1999. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies [J]. New Phytologist, 143(143): 155-162.
WITKOWSKI E T F, LAMONT B B. 1991. Leaf specific mass confounds leaf density and thickness [J]. Oecologia, 88(4): 486-493.
WRIGHT I J, REICH P B, WESTOBY M, et al. 2004. The worldwide leaf economics spectrum [J]. Nature, 428(6985): 821-827.
YAN E R, MILLA R, AARSSEN L W, et al. 2012. Functional relationships of leafing intensity to plant height, growth form and leaf habit [J]. Acta Oecologica, 41(4): 20-29.
黄海侠, 杨晓东, 孙宝伟, 等. 2013. 浙江天童常绿植物当年生与往年生叶片性状的变异与关联[J]. 植物生态学报, 37(10): 912-921.
李西良, 刘志英, 侯向阳, 等. 2015. 放牧对草原植物功能性状及其权衡关系的调控[J]. 植物学报, 50(2): 159-170.
沈年伟, 钱前, 张光恒. 2009. 水稻卷叶性状的研究进展及在育种中的应用[J]. 分子植物育种,7(5): 852-860.
王常顺, 汪诗平. 2015. 植物叶片性状对气候变化的响应研究进展[J].植物生态学报, 39(2): 206-216.
Response of Grassland Using Modes to Leaf Trait of Stipa breviflora in Desert Steppe of Inner Mongolia
LIU Wenting1, WEI Zhijun1*, LV Shijie2, BAI Yuting1, XIAO Jiapu1, DAI Jingzhong1, SUN Shixian3
1. College of Ecology and Environmental Science, Inner Mongolia Agricultural University, Huhhot 010019, China;2. College of Science, Inner Mongolia Agricultural University, Huhhot 010018, China;3. Institute of Grassland Research, Chinese Academy of Agriculture Sciences, Hohhot 010010, China
Abstract:The trade-off of plant function traits to adapt the different habitats is a kind of effective ecological strategy. Stipa breviflora is the main edificator in warm-temperate desert grassland. The response of S. breviflora leaf traits to grassland using modes may reveal the modern desert grassland resource utilization and plant ecological response mechanism. Therefore, we investigated S. breviflora by measuring its leaf traits in different age classes in the manual irrigation grassland, grazied grassland and ungrazed grassland. The results showed S. breviflora's leaf natural height, leaf length, leaf fresh weight, leaf dry weight, leaf width and leaf edge distance in the manual irrigation grassland were larger than the leaf traits in the grazed grassland and in the ungrazed grassland. Leaf erection index and leaf fresh weight were sensitive characters to habitat disturbance. Leaf width and leaf dry matter content were inert characters. The degree of response to grassland using modes and its variation fit the linear regression relationship (y=ax+b) on S. breviflora leaf traits. By pearson correlation analysis, there was a greater influence on the leaf traits to grazing. Our study suggested grassland using modes led to the leaf traits differentiation of the desert grassland edificator in Mongolian Plateau, S. breviflora plasticity trade-off through a variety of leaf traits in response to the habitats to complete its life cycle under the metastable ecological system.
Key words:grassland using modes; trade-off; S. breviflora; leaf traits; age classes
DOI:10.16258/j.cnki.1674-5906.2016.03.004
中图分类号:Q948; X17
文献标志码:A
文章编号:1674-5906(2016)03-0385-08
基金项目:国家自然科学基金项目(31460126);国家重点基础研究发展计划(2014CB138805);国家科技支撑计划(2012BAD13B00)
作者简介:刘文亭(1990年生),男,博士研究生。E-mail: nmgliuwenting@163.com
*通信作者。E-mail: nmndwzj@163.com
收稿日期:2015-10-25
