内蒙古典型草原凋落物分解对不同草地利用方式的响应
2016-06-28单玉梅温超杨勇陈迪马白永飞陈海军王明玖
单玉梅,温超,杨勇,陈迪马,白永飞,陈海军,王明玖
1. 内蒙古自治区农牧业科学院,内蒙古 呼和浩特 010031;2. 中国科学院内蒙古草业研究中心,内蒙古 呼和浩特 010031;3. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;4. 中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093;5. 内蒙古自治区草原勘察规划院,内蒙古 呼和浩特 010051;6. 内蒙古自治区生物技术研究院,内蒙古 呼和浩特 010010
内蒙古典型草原凋落物分解对不同草地利用方式的响应
单玉梅1, 2, 3, 4,温超1, 2,杨勇5,陈迪马4,白永飞4,陈海军3, 6*,王明玖3*
1. 内蒙古自治区农牧业科学院,内蒙古 呼和浩特 010031;2. 中国科学院内蒙古草业研究中心,内蒙古 呼和浩特 010031;3. 内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;4. 中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093;5. 内蒙古自治区草原勘察规划院,内蒙古 呼和浩特 010051;6. 内蒙古自治区生物技术研究院,内蒙古 呼和浩特 010010
摘要:以2005年设置的中德放牧控制试验为平台,研究了内蒙古典型草原群落凋落物的分解速率、失重率及其碳氮损失率在不同草地利用方式和放牧强度下的变化规律。草地利用方式包括:传统放牧、传统割草和放牧、割草轮换利用(混合利用)。放牧强度包括7个:0、1.5、3.0、4.5、6.0、7.5、9.0 sheep·hm-2。结果表明:传统放牧和混合利用方式下,放牧都不同程度地加速了群落凋落物的分解和失重。在传统放牧平地系统中,放牧强度为7.5 sheep·hm-2小区群落凋落物的分解速率和失重率均显著高于对照区,分别高出45.0%和40.0%;在混合利用平地系统中,放牧强度为6.0 sheep·hm-2小区群落凋落物的分解速率显著高于对照区,高出35.0%;在混合利用坡地系统中,放牧强度为9.0 sheep·hm-2小区群落凋落物的分解速率和失重率均显著高于对照区,分别高出82.4%和62.0%。群落凋落物碳损失率和氮损失率在传统放牧系统中,随放牧强度的增加呈现出相似的变化趋势,而在混合利用系统中,随放牧强度的增加呈现出相反的变化趋势,但是总体表现为,增加放牧强度会加速群落凋落物中碳氮的释放。高放牧强度下,群落凋落物的分解速率、失重率和碳损失率差异显著,表现为:混合利用>传统割草>传统放牧,这一试验结果在一定程度上验证了放牧优化假说。
关键词:草地利用方式;放牧;割草;凋落物;放牧强度;碳和氮
引用格式:单玉梅, 温超, 杨勇, 陈迪马, 白永飞, 陈海军, 王明玖. 内蒙古典型草原凋落物分解对不同草地利用方式的响应[J]. 生态环境学报, 2016, 25(3): 377-384.
SHAN Yumei, WEN Chao, YANG Yong, CHEN Dima, BAI Yongfei, CHEN Haijun, WANG Mingjiu. Impacts of Management Regimes on the Litter Decomposition of Typical Grassland Community in Inner Mongolia [J]. Ecology and Environmental Sciences,2016, 25(3): 377-384.
凋落物不仅是草原第一生产力的重要组成部分,也是草地生态系统结构的重要组成部分,植物通过光合作用所固定的有机质最终以凋落物形式储存于地表(郭继勋等,1994)。凋落物的产生、积累和分解动态直接或间接地影响着植物的萌发与保护(Molofsky et al.,1992;郭继勋等,2000)、群落结构演替(Facelli et al.,1991),在改善生态环境及土壤理化性状(Naeth et al.,1991)、能量流动和营养物质循环以及生产力维持过程(郭继勋等,1994;高永恒等,2007;陈瑾等,2011;王静等,2013)中起着重要作用。其中凋落物分解既是草地生态系统中有机质残体分解转化的基本过程,也是生态系统养分循环中的重要环节(高永恒等,2007;陈瑾等,2011),生态系统通过凋落物分解将养分释放到土壤中,补充植物生长所需,对调节土壤养分的可利用性和维持草地生产力具有重要作用(Koukoura,1998;汪诗平等,2001;高永恒等,2007;Meyer et al.,2011;陈瑾等,2011)。
长期以来,国内外的研究者对凋落物分解和养分释放的影响因素进行了深入的研究。归纳起来,主要包括植物群落结构(Facelli et al.,1991;王静等,2013)、凋落物自身的特性(或基质质量,Substrate quality)(Aerts et al.,1997;Vitousek et al.,1994;Kalburtji et al.,1999;Hobbie et al.,2000;陈瑾等,2011)、土壤理化性状(Aerts et al.,1997)、生物环境(Setala et al.,1990)、环境因子(Vitousek et al.,1994;Austin et al.,2000)以及草地利用方式(Hamilton et al.,2001;Willms et al.,2002;高永恒等,2007)等方面,其中草地利用方式是诸多因素当中最重要的外因之一(汪诗平等,1999;Hamilton et al.,2001;Willms et al.,2002;高永恒等,2007;薛睿等,2010)。在内蒙古典型草原生态系统中,放牧和打草是两种主要的草地利用方式。已有研究表明,放牧(特别是过度放牧)可以改变群落结构和影响植物繁殖特性,引起凋落物质的降低和量的减少,草地生产力显著下降(汪诗平等,1999),严重影响生态系统的养分循环和服务功能(Wardle et al.,2002;Bai et al.,2004;薛睿等,2010;Shan et al.,2011)。打草是刈割方式的一种,适当的刈割不仅能改善草地生态环境,刺激牧草分蘖,促进牧草的再生和凋落物的积累,在一定程度上也能缓解放牧对草原的持续性压力。但是连续打草不利于牧草的再生,因为连续打草严重影响牧草种子的繁殖,抑制其持续性生长和更新(陈佐忠等,2000)。同时,打草也是放牧的代替性行为,连续打草会导致草地上的群落凋落物减少,生态系统养分循环受阻,养分输入减少,土壤日益贫瘠(Augustine et al.,2006;Ilmarinen et al.,2009)。近年来,内蒙古草原退化严重,草原群落的初级生产力逐渐下降,呈现出诸多的生态环境问题,主要归因于不合理的草地利用方式(王炜等,1996a,1996b;Bai et al.,2007)。因此,合理地协调放牧与打草的关系,使二者有机结合,提高生态系统养分循环速率是维持草地可持续发展的动力之一(薛睿等,2010)。
目前,针对凋落物分解的研究不仅集中在继续探讨分解时的营养动态、分解与凋落物质量、生境、生物以及与气候间的相互关系(郭继勋等,1994;郭继勋等,2000;陈瑾等,2011),也将分解与草原生态环境问题结合起来,并涉及到营养循环等生态系统过程(王蕾等,2013),而对不同草地利用方式下凋落物分解过程的研究少见报道。为此,本研究以内蒙古典型草原为研究对象,基于长期放牧控制实验平台,从群落水平上对不同草地利用方式和放牧强度下凋落物的分解特性及其碳氮损失率进行了野外试验测定和室内分析,以探讨该类型的生态系统中草地利用方式对凋落物分解过程的影响,旨在为内蒙古典型草原生态系统的科学管理提供理论依据,拟回答两个方面的科学问题,(1)不同草地利用方式对内蒙古典型草原凋落物分解有何影响?(2)群落凋落物分解速率以及碳氮损失率随放牧强度(尤其是过度放牧)的变化规律如何?
1 研究地区与研究方法
1.1 研究区自然概况
本试验在中国科学院内蒙古草原生态系统定位研究站的中德放牧试验样地(43°37′N,116°43′E)进行。该站位于内蒙古锡林河流域,平均海拔在1100 m左右,气候类型为温带半干旱大陆性季风气候,冬季寒冷干燥,夏季温暖湿润。多年平均气温为0.4 ℃,其中最冷月(1月份)均温为-21.4 ℃,最热月(7月份)均温为19.0 ℃。≥10 ℃的年积温约为1600 ℃,多年平均降水量为337 mm,主要集中分布于5─8月份,降水量的年际变异较大。地带性土壤类型为栗钙土(陈佐忠等,2000)。群落类型为典型的羊草草原,常见植物约有45种,以广旱生根茎禾草羊草(Leymus chinensis)占显著优势,其次为大针茅(Stipa grandis)、菭草(Koeleria cristata)和冰草(Agropyron cristatum)等旱生密丛禾草,这些禾草的相对生物量达到60%以上;杂类草中80%为多年生,如黄囊苔草(Carex rostrata kor-shinskyi)、星毛委陵菜(Potentilla acaulis)、野韭(Allium ramosum)等(Bai et al.,2007)。
1.2 研究方法
1.2.1 试验设计
放牧控制实验平台始建于2005年6月,占地面积近300 hm2。实验处理包括两种放牧制度,即传统利用(放牧场和割草场分开,放牧场每年生长季放牧,割草场每年秋季割草)和混合利用(放牧场和割草场每年进行轮换);包括两种草地利用方式,即放牧和割草。每种利用方式设有7个放牧强度(0、1.5、3.0、4.5、6.0、7.5、9.0 sheep·hm-2)和2个区组重复(平地和坡地),实验采用随机区组设计,共计56个实验小区。为了消除空间异质性,每个区组下的主效应为利用方式,对每种利用方式下的7个放牧强度采用裂区设计,即两个相邻小区的放牧强度相同。其中,1.5 sheep·hm-2水平下的放牧小区和对应的割草小区面积为4 hm2(确保每个试验小区至少有6只羊),其它试验小区均为2 hm2。放牧时间为每年的6月初到9月中旬,持续100 d左右,晚上不归牧;打草时间为每年的8月中旬。
1.2.2 样品的采集和处理
于2009年5月,采用尼龙网袋法在每个小区中随机收集植物群落凋落物样品,去除其中杂物,将剩余凋落物用清水洗涤干净装入信封置于65 ℃恒温条件下烘干至恒重后,再将凋落物剪成10 cm左右小段,称取样品10 g,装入已编号的尼龙网袋中,网袋大小为15 cm×25 cm,网孔为1 mm×1 mm(陈瑾等,2011),放回相应的小区内,为使其充分与地面接触采用铁丝固定于地表,每个小区放3袋,作为3个重复。在分装凋落物的同时,取出一部分用于测定其全碳、全氮及全磷含量,即分解前凋落物的碳氮磷含量。生长季结束时,于2009 年10月取回初期装有凋落物的尼龙袋,并去除其中杂物(如小石砾、钻进网袋内的小动物等)后用清水漂洗,使用60目的筛子协助漂洗。漂洗后立即装入信封袋中,再置于65 ℃恒温下烘48 h后称其干重,然后粉碎,测定分解后凋落物中的碳氮磷含量。
1.3 样品测定
粉碎后的凋落物样品,用H2O2/H2SO4消煮,消煮液用于测定全氮(N)、全磷(P)的含量。全N的测定采用半微量凯氏定氮法,全P的测定采用钼蓝比色法。初始凋落物除用于测定全N、全P的含量外,还采用重铬酸钾外加热氧化法测定全碳(C)含量(董鸣等,1996)。
1.4 数据处理与统计分析
为了更加直观地反映凋落物损失情况,采用变化率来表示分解过程中凋落物质量和元素含量的变化,主要测定指标包括分解速率(r)、失重率(RW)以及碳氮损失率(RC、RN)。其计算公式如下:

其中,r为凋落物分解速率;RW为凋落物失重率;t1和t2分别为野外放置凋落物的时间和取回凋落物的时间;W1和W2分别为t1和t2时凋落物的重量;RC和RN分别为凋落物碳氮损失率;C1(N1)和C2(N2)分别为t1和t2时凋落物有机碳(全氮)含量。
所有数据采用SAS(Version 9.0)软件进行统计分析。采用Repeated Measure ANOVA 方法来分析放牧制度(传统利用和混合利用)、利用方式(放牧和割草)、放牧强度(0、1.5、3.0、4.5、6.0、7.5 和9.0 sheep·hm-2)和地形(平地和坡地)及其交互作用对群落凋落物分解速率的影响。采用One-way ANOVA方法来分析传统放牧平地、传统放牧坡地、混合利用平地和混合利用坡地4个系统中,不同放牧强度下群落凋落物的分解速率、失重率和碳氮损失率以及不同利用方式下群落凋落物分解速率、失重率和碳氮损失率的差异并用Duncan′s极差法进行显著性检验(P<0.05表示为差异显著)。
2 结果与分析
2.1 不同放牧强度下凋落物分解速率和失重率的变化
多因素方差分析表明,放牧强度对凋落物分解速率的影响极显著(P<0.001),而放牧制度(传统放牧和混合利用)、利用方式(放牧和割草)和地形(平地和坡地)对凋落物分解速率的影响不显著,但放牧制度和利用方式及放牧强度和地形的两因素交互作用对分解速率的影响均显著(P<0.001),且放牧强度与放牧制度、利用方式和地形的三因素交互作用对群落凋落物分解速率的影响均显著(P<0.05),四因素交互作用不显著(表1)。
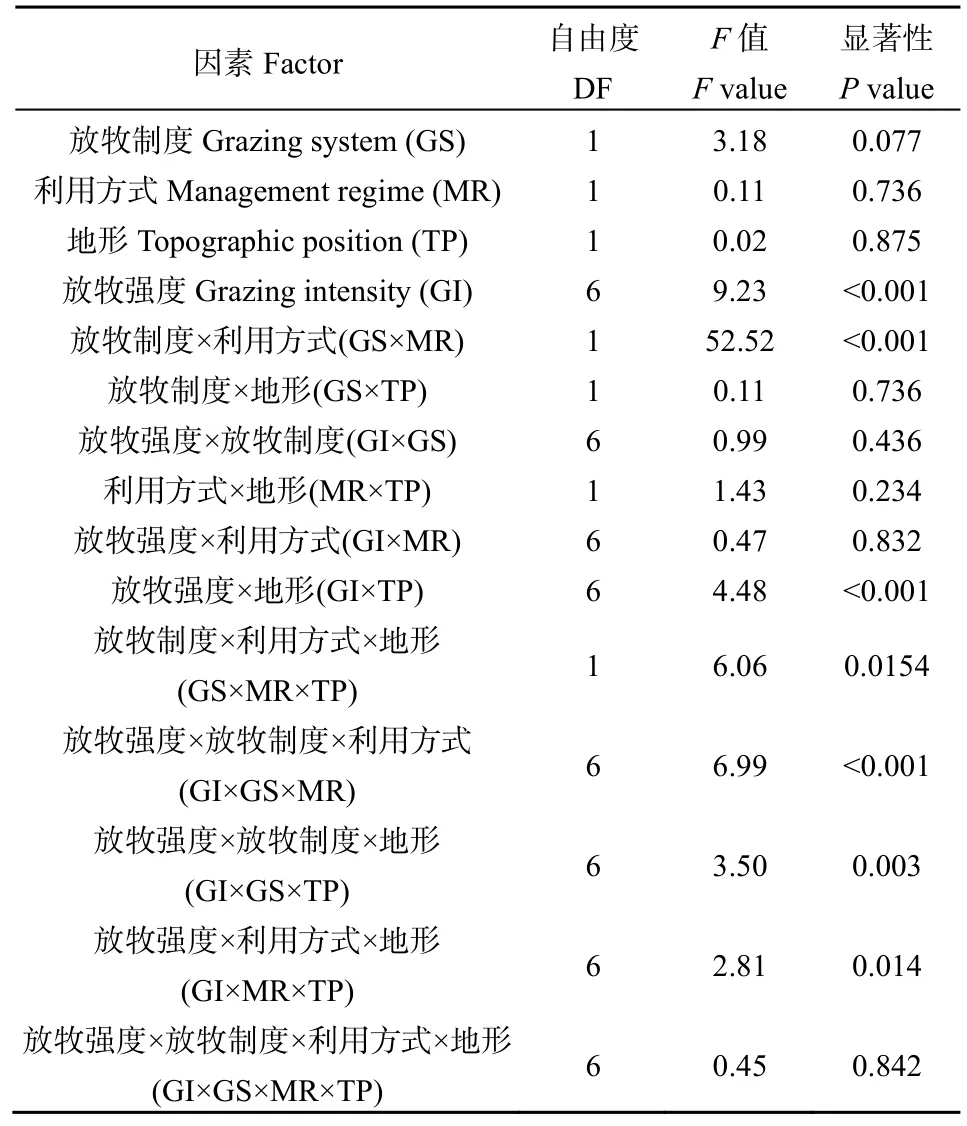
表1 放牧制度、利用方式、地形和放牧强度对群落凋落物分解速率的影响Table 1 Effects of grazing system, management regime, topography and grazing intensity on the decomposition rate of litter in community
在传统放牧平地(TGF)、传统放牧坡地(TGS)、混合利用平地(MGF)和混合利用坡地(MGS)4个系统中,群落凋落物的分解速率和失重率随放牧强度的增加呈现出相同的变化趋势,且混合利用方式下群落凋落物的分解速率和失重率高于对照区(图1A)。在TGF中,放牧强度为7.5 sheep·hm-2小区群落凋落物的分解速率和失重率显著高于对照区(P<0.05),分别高出45.0%和40.0%。但是在TGS中,群落凋落物的分解速率和失重率在各个放牧强度之间差异均不显著(P>0.05)。在MGF中,放牧强度为6.0 sheep·hm-2小区群落凋落物的分解速率显著高于对照区,高出35.0%(P<0.05),而各个放牧强度之间的失重率差异不显著(P>0.05)。在MGS中,放牧强度为9.0 sheep·hm-2小区群落凋落物的分解速率和失重率显著高于对照区(P<0.05),分别高出82.4%和62.0%(图1A)。
2.2 不同放牧强度下凋落物碳氮损失率的变化
在传统放牧系统中,群落凋落物碳损失率和氮损失率随放牧强度的增加呈现出相似的变化趋势,而在混合利用系统中,群落凋落物碳损失率和氮损失率随放牧强度的增加呈现出相反的变化趋势(图1B)。

图1 传统放牧和混合利用系统中,不同放牧强度下群落凋落物分解速率(r)、失重率(RW)(A)以及碳氮损失率(RC、RN)(B)的变化。TGF:传统放牧平地系统;TGS:传统放牧坡地系统;MGF:混合放牧平地系统;MGS:混合放牧坡地系统Fig. 1 Decomposition rate (r), weightless rate (RW) (A) and relative carbon and nitrogen loss (RC and RN) (B) in the litters at different grazing intensities in traditional and mixed grazing systems. TGF, Traditional grazing flat system; TGS, Traditional grazing slope system; MGF, Mixed grazing flat system; MGS,Mixed grazing slope system (n=3)
在TGF中,放牧强度为6.0 sheep·hm-2小区群落凋落物碳氮损失率最小(分别为14.0%和0.7%),放牧强度为7.5 sheep·hm-2小区群落凋落物碳损失率最大(31.4%),而对照区群落凋落物氮损失率最大(1.2%)。在TGS中,群落凋落物碳氮损失率随放牧强度的增加呈现增加的趋势,碳损失率的最大值出现在放牧强度为7.5 sheep·hm-2的小区,高出对照区47.5%,存在显著差异(P<0.05),氮损失率的最大值出现在放牧强度为9.0 sheep·hm-2的小区,显著高于对照区27.0%(P<0.05)。在MGF中,群落凋落物碳损失率随放牧强度的增加呈现逐渐增加的趋势,放牧区显著高于对照区(P<0.05),且放牧强度为9.0 sheep ·hm-2的小区群落凋落物碳损失率为对照区的1.5倍;而群落凋落物氮损失率的最大值出现在放牧强度为6.0 sheep·hm-2的小区。在MGS中,放牧区群落凋落物碳损失率显著高于对照区(P<0.05),最大值出现在放牧强度为3.0 sheep·hm-2的小区,而放牧区群落凋落物氮损失率除放牧强度为1.5 sheep·hm-2的小区外,其他小区均显著低于对照区(P<0.05)(图1B)。
2.3 草地利用方式对凋落物分解速率和失重率的影响
在放牧强度为9.0 sheep·hm-2(代表过度放牧利用)的小区,平地系统中群落凋落物分解速率和失重率呈现出相同的变化趋势,传统割草>混合利用>传统放牧(图2A),各处理间差异不显著(P>0.05);坡地系统中群落凋落物分解速率和失重率的变化趋势相一致,即混合利用>传统割草>传统放牧(图2B),且混合利用系统群落凋落物分解速率和失重率显著高于传统放牧系统(P<0.05)。
2.4 草地利用方式对凋落物分解过程中碳氮损失率的影响
在放牧强度为9.0 sheep·hm-2的小区,平地系统中,群落凋落物碳损失率大小依次为混合利用>传统割草>传统放牧,且混合利用系统显著高于传统割草系统和传统放牧系统(P<0.05);群落凋落物氮损失率则为传统割草>传统放牧>混合利用,传统割草系统凋落物氮损失率显著高于传统放牧系统,传统放牧系统又显著高于混合利用系统(P<0.05)(图2A)。在坡地系统中,凋落物碳损失率在各处理间差异不显著(P>0.05);凋落物氮损失率的变化趋势和平地系统中相同(P<0.05)(图2B)。可见,从群落凋落物分解碳氮损失角度而言,今后在制定草地管理措施时不仅要考虑放牧强度的大小,还应考虑地形因素的影响。
3 讨论
3.1 放牧强度对凋落物分解的影响
凋落物分解是草地生态系统中有机质残体分解转化的基本过程,也是系统养分循环的重要环节,对调节土壤养分的可利用性和维持草地生产力具有重要作用(Koukoura,1998;汪诗平等,2001)。本研究中,不同的草地利用方式下,凋落物的分解速率和失重率随着放牧强度的增加都呈现出增加的趋势,放牧区的碳氮损失率也显著高于对照区,一方面放牧家畜的践踏活动促使凋落物与地面的紧密接触,有助于凋落物的分解(Manley et al.,1995;高永恒等,2007);另一方面,家畜粪便中的养分增强了土壤微生物的活性(Shan et al.,2011),从而加速了土壤中有机质的分解速率。放牧不仅促进了凋落物中碳氮的释放,也有利于群落凋落物中的碳氮参与草地生态系统的碳氮循环,对维持草地生态系统中碳氮的平衡具有重要意义(Shariff et al.,1994)。但重度放牧尽管加速凋落物的分解,但同时会大幅度地减少群落盖度和地上现存生物量(薛睿等,2010),这也不利于草地生态系统的健康发展。因此,适度放牧有利于调节草地生态系统的碳氮循环(Mcnaughton,1979;Schuman et al.,1999)。
3.2 草地利用方式对凋落物分解的影响
放牧与割草轮换利用的草地利用方式可以减缓持续放牧带来的压力,对草场起到较好的保护作用。尤其是在过度放牧的情况下,混合利用方式是最佳的利用方式之一(薛睿等,2010)。本研究中,在放牧强度为9.0 sheep·hm-2时,混合利用方式下的群落凋落物的分解速率和失重率最大,且在MGF中的群落凋落物碳损失率也最大,这说明混合利用方式有利于草地生态系统的养分循环。传统放牧系统一直面临持续性的压力,而传统割草系统会因割草量过多而使地上生物量减少,导致土壤养分流失(Ilmarinen et al.,2009)。但是,本研究中混合利用系统并不利于群落凋落物氮的释放,导致这一结果的原因可能是,放牧可以通过改变土壤的微气候,如土壤温度和湿度,增加土壤氮的有效性和土壤微生物的活性(Shariff et al.,1994;Kielland et al.,1998),已有数据表明,传统割草系统土壤湿度比混合利用系统高出30.3%,传统放牧系统土壤温度比混合利用系统高出20.0%,因此,混合利用系统中群落凋落物氮释放较低。但是,不同利用方式下的各个物种凋落物的分解速率及碳氮释放特性还有待进一步的研究。
4 结论
以内蒙古典型草原为研究对象,基于长期放牧控制实验平台(已具有连续11年放牧的历史),对草原群落凋落物的分解速率、失重率及其碳氮损失率在不同草地利用方式下的变化规律与响应过程进行了研究,结果表明:在传统放牧和混合利用方式下,放牧活动均不同程度地加速了群落凋落物的分解和失重。在传统放牧平地系统中,放牧强度为7.5 sheep·hm-2的小区群落凋落物的分解速率和失重率分别显著高于对照区;在混合利用平地系统中,放牧强度为6.0 sheep·hm-2的小区群落凋落物的分解速率高出对照区35.0%,存在显著差异;在混合利用坡地系统中,放牧强度为9.0 sheep·hm-2的小区群落凋落物的分解速率和失重率分别显著高于对照区。此外,群落凋落物碳氮损失率在传统放牧系统中,随放牧强度的增加呈现出相似的变化趋势,而在混合利用系统中,随放牧强度的增加呈现出相反的变化趋势,总体表现为,放牧强度的增加会加速群落凋落物中碳氮的释放。在高放牧强度下,群落凋落物的分解速率、失重率和碳损失率差异显著,均表现为:混合利用>传统割草>传统放牧,该试验结果在一定程度上验证了放牧优化假说。
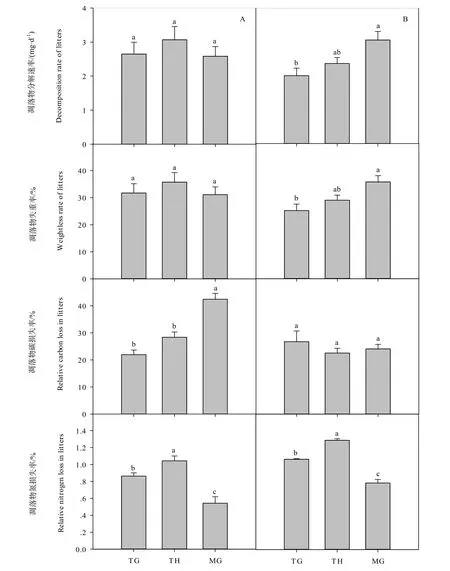
图2 凋落物分解速率、失重率以及碳氮损失率在传统放牧、传统割草和混合利用的平地(A)和坡地(B)系统中的变化。不同小写字母表示各个指标在不同草地利用方式下具有显著差异(P<0.05)。TG:传统放牧系统;TH:传统割草系统;MG:混合放牧系统Fig. 2 The variation of decomposition rate, weightless rate and relative carbon and nitrogen loss of community litters in traditional grazing, traditional haymaking and mixed grazing systems in flat (A) and slope (B) blocks. Different lowercase letters denote significant difference among different grassland management types at P < 0.05. TG, Traditional grazing system; TH, Traditional haymaking system; MG, Mixed grazing system (n=3)
致谢:感谢中国科学院内蒙古草原生态系统定位研究站的工作人员和师弟师妹们在野外工作和室内样品分析中给予的大力帮助。
参考文献:
AERTS R. 1997. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship [J]. Oikos, 79(3): 439-449.
AERTS R, HANNIE D C. 1997. Nutritional and plant mediated controls on leaf litter decomposition of Carex species [J]. Ecology, 78(1): 244-260.
AUGUSTINE D J, MCNAUGHTON S J. 2006. Interactive effects of ungulate herbivores, soil fertility, and variable rainfall on ecosystem processes in a semiarid savanna [J]. Ecosystems, 9(8): 1242-1256.
AUSTIN A T, VITOUSEK P M. 2000. Precipitation, decomposition and decomposability of Metrosideros polymorpha in native forests on Hawai'i [J]. The Journal of Ecology, 88(1): 129-138.
BAI Y F, HAN X G, WU J G, et al. 2004. Ecosystem stability and compensatory effects in the Inner Mongolia grassland [J]. Nature,431(7005): 181-184.
BAI Y F, WU J G, PAN Q M, et al. 2007. Positive linear relationship between productivity and diversity: evidence from the Eurasian Steppe [J]. Journal of Applied Ecology, 44(5): 1023-1034.
FACELLI J, PICKETT S T A. 1991. Plant litter: Its dynamics and effects on plant community structure [J]. Botanical Review, 57(1): 1-32.
HAMILTON E W, FRANK D A. 2001. Can plants stimulate soil microbes and their own nutrient supply? Evidence from a grazing tolerant grass [J]. Ecology, 82(9): 2397-2402.
HOBBIE S E, VITOUSEK P M. 2000. Nutrient limitation of decomposition in Hawaiian forests [J]. Ecology, 81(7): 1867-1877.
ILMARINEN K, MIKOLA J, NISSINEN K, et al. 2009. Role of soil organisms in the maintenance of species-rich seminatural grasslands through mowing [J]. Restoration Ecology, 17(1): 78-88.
KALBURTJI K L, MOSJIDIS J A, MAMOLOS A P. 1999. Litter dynamics of low and high tannin serica lepedeza plants under field conditions [J]. Plant and Soil, 208(2): 271-281.
KIELLAND K, BRYANT J P. 1998. Moose herbivores in taiga: effects on biogeochemistry and vegetation dynamics in primary succession [J]. Oikos, 82(2): 377-383.
KOUKOURA Z. 1998. Decomposition and nutrient release from C3 and C4 plant litters in a natural grassland [J]. Acta Oceologica, 19(2): 115-123.
MANLEY J T, SCHUMAN G E, REEDER J D, et al. 1995. Rangeland soil carbon and nitrogen responses to grazing [J]. Journal of Soil and Water Conservation, 50(3): 294-298.
MCNAUGHTON S J. 1979. Grazing as an optimization process:grassungulate relationships in the Serengeti [J]. The American Naturalist,113(5): 691-703.
MEYER III WM, OSTERTAG R, COWIE R H. 2011. Macro-inverte-brates accelerate litter decomposition and nutrient releasein a Hawaiian rainforest [J]. Soil Biology and Biochemistry , 43(1): 206-211.
MOLOFSKY J, AUGSPURGER C K. 1992. The effect of leaf litter in early seeding establishment in a tropical forest [J]. Ecology, 73(1): 68-77.
NAETH M A, BAILEY A W, CHANASYK D S. 1991. Water holding capacity of litter and soil organic matter in mixed prairie and fescue grassland ecosystems of Alberta [J]. Journal of Range Management,44(1): 13-17.
SCHUMAN G E, REEDER J D, MANLEY J T, et al. 1999. Impact of grazing management on the carbon and nitrogen balance of mixed-grass rangeland [J]. Ecological Applications, 9(1): 65-71.
SETALA H, HUHTA H. 1990. Evaluation of the soil faunal impact on decomposition in a simulated coniferous forest soil [J]. Biology and Fertility of Soils, 10(3): 163-169.
SHAN Y M, CHEN D M, GUAN X X, et al. 2011. Seasonally dependent impacts of grazing on soil nitrogen mineralization and linkages to ecosystem functioning in Inner Mongolia grassland [J]. Soil Biology and Biochemistry, 43(9): 1943-1954.
SHARIFF A R, BIONDINI M E, GRYGIEL C E. 1994. Grazing intensity effects on litter decomposition and soil nitrogen mineralization [J]. Journal of Range Management, 47(6): 444-449.
VITOUSEK P, TURNER D, PARTON W, et al. 1994. Litter decomposition on the Mauna Loa environmental matrix, Hawaii: patterns,mechanisms, and models [J]. Ecology, 75(2): 418-429.
WARDLE D A, BONNER K I, BARKER G M. 2002. Linkages between plant litter decomposition, litter quality, and vegetation responses to herbivores [J]. Functional Ecology, 16(5): 585-595.
WILLMS W D, DORMAAR J F, ADAMS B W. 2002. Response of the mixed prairie to protection from grazing [J]. Journal of Range Management, 55(3): 210-216.
陈瑾, 李扬, 黄建辉. 2011. 内蒙古典型草原4种优势植物凋落物的混合分解研究[J]. 植物生态学报, 35(1): 9-16.
陈佐忠, 汪诗平. 2000. 中国典型草原生态系统[M]. 北京: 科学出版社.
董鸣, 王义凤, 孔繁志, 等. 1996. 陆地生物群落调查观测与分析[M].北京: 中国标准出版社: 152-154.
高永恒, 陈槐, 罗鹏, 等. 2007. 放牧强度对川西北高山草甸两个优势物种凋落物分解的影响[J]. 生态科学, 26(3): 193-198.
郭继勋, 孙刚, 姜世成. 2000. 松嫩平原羊草草原凋落物层群落学作用的研究[J]. 植物生态学报, 24(4): 473-476.
郭继勋, 祝廷成. 1994. 羊草草原枯枝落叶积累的研究—自然状态下枯枝落叶的积累及放牧、割草对积累量的影响[J]. 生态学报, 14(3): 255-259.
汪诗平, 李永宏. 1999. 内蒙古典型草原退化机理的研究[J]. 应用生态学报, 10(4): 437-441.
汪诗平, 李永宏, 王艳芬, 等. 2001. 不同放牧率对内蒙古冷蒿草原植物多样性的影响[J]. 植物学报, 43(1): 89-96.
王静, 赵萌莉, WILLMS W, 等. 2013. 内蒙古典型草原不同功能群生产力对凋落物去除的响应[J]. 草业学报, 22(6): 31-38.
王蕾, 张宇, 许冬梅, 等. 2013. 围封对草地凋落物分解速率和N、P、K含量的影响[J]. 草业科学, 30(10): 1502-1508.
王炜, 刘钟龄, 郝敦元, 等. 1996a. 内蒙古草原退化群落恢复演替的研究. I. 退化草原的基本特征与恢复演替动力[J]. 植物生态学报,20(5): 449-459.
王炜, 刘钟龄, 郝敦元, 等. 1996b. 内蒙古草原退化群落恢复演替的研究. II. 恢复演替时间进程的分析[J]. 植物生态学报, 20(5): 460-471.
薛睿, 郑淑霞, 白永飞. 2010. 不同利用方式和载畜率对内蒙古典型草原群落初级生产力和植物补偿性生长的影响[J]. 生物多样性,18(3): 300-311.
Impacts of Management Regimes on the Litter Decomposition of Typical Grassland Community in Inner Mongolia
SHAN Yumei1, 2, 3, 4, WEN Chao1, 2, YANG Yong5, CHEN Dima4, BAI Yongfei4,CHEN Haijun3, 6*, WANG Mingjiu3*
1. Inner Mongolia Academy of Agriculture & Animal Husbandry Science, Hohhot 010031, China;2. Inner Mongolia Research Center for Prataculture, Chinese Academy of Science, Hohhot 010031, China;3. College of Ecology and Environmental Science, Inner Mongolia Agricultural University, Hohhot 010019, China;4. State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China;5. Inner Mongolia Institute of Grassland Survey and Planning, Huhhot 010051, China; 6. Inner Mongolia Institute of Biotechnology, Huhhot 010010, China
Abstract:The study was carried out in the Sino-German grazing experiment site, which was established in June 2005, and located in the typical steppe region of Inner Mongolia grassland In order to explore two Our objective, 1) how litter decomposition vary with grazing intensity and haymaking in typical steppe of Inner Mongolia grassland, and 2) how the litter decomposition rate, weightless rate, relative carbon and nitrogen loss rates respond to grazing intensity. The experimental treatments included traditional grazing,traditional haymaking and alternate mixed utilization of grazing & haymaking, and seven levels of grazing intensity (i.e., 0, 1.5, 3.0,4.5, 6.0, 7.5, 9.0 sheep·hm-2). Our results showed that grazing accelerated decomposition and weightless of litters in both traditional and mixed grazing plots after one growing season. In flat area of traditional grazing plots, the decomposition and weightless rates of litters in 7.5 sheep·hm-2plots were significantly higher than that in control plots by 45.0% and 40.0%, respectively. In flat area of mixed grazing plots, the decomposition rate of litters in 6.0 sheep·hm-2plots was significantly higher than control plot by 35.0%. While in slop area of mixed grazing plots, the decomposition and weightless rates of litters in 9.0 sheep·hm-2plots were significantly higher than that in control plots by 82.4% and 62.0%, respectively. With the increases of grazing intensity, the variations of carbon and nitrogen loss in litters were similar in traditional system, whereas they showed divergent trends in mixed system. But as a whole,grazing accelerated carbon and nitrogen releasing during litter decomposition. At high grazing intensity, the decomposition,weightless and relative carbon loss rates of litters in mixed grazing plots were significantly higher than that in the traditional grazing plots. Our results were partially in agreement with the grazing optimization hypothesis.
Key words:management regimes; grazing; haymaking; litter; grazing intensity; carbon and nitrogen
DOI:10.16258/j.cnki.1674-5906.2016.03.003
中图分类号:Q948; X17
文献标志码:A
文章编号:1674-5906(2016)03-0377-08
基金项目:国家自然科学基金项目(31200351);内蒙古科技创新引导奖励基金计划项目(20140701);内蒙古自治区自然科学基金项目(2014MS0318);内蒙古自治区自然科学基金项目(2015MS3084)
作者简介:单玉梅(1980年生),女,博士,助理研究员,主要从事草地放牧生态系统结构与功能方面的研究。E-mail: shanym2008@163.com*通信作者,E-mail: wangmj_0540@163.com; chenhaijun2004@163.com
收稿日期:2016-01-13
