菜心Hau CMS雄性不育系的转育研究
2016-06-17陈汉才李桂花黎庭耀
张 艳,陈汉才,李桂花,黎庭耀
(广东省农科院蔬菜研究所/广东省蔬菜新技术研究重点实验室,广东 广州 510640)
菜心Hau CMS雄性不育系的转育研究
张 艳,陈汉才,李桂花,黎庭耀
(广东省农科院蔬菜研究所/广东省蔬菜新技术研究重点实验室,广东 广州 510640)
摘 要:以甘蓝型油菜细胞质雄性不育系Hau CMS为不育胞质供体,菜心为父本,进行种间杂交和连续回交,获得了一个新型菜心Hau CMS不育系,并对不同回交世代材料进行了植物学性状、分子生物学和细胞学的研究。结果表明:F1和各回交世代未出现育性分离现象,不育率和不育度均为100%。F1代植株表型有分离,绝大部分特征偏向于母本,利用菜心连续回交,到BC3群体已基本趋向轮回亲本。用19对SRAP引物对F1和回交世代进行遗传变异的分析,结果显示:随着回交次数的增加,母本特异性条带逐渐消失,缺失条带比率从2.30%(F1)上升至50.42%(BC4),BC4群体母本特异性条带占总带型比例为4.16%。石蜡切片观察发现菜心Hau CMS不育系的花药败育发生于雄蕊原基分化时期,没有孢原细胞的分化,无花粉囊的形成。
关键词:菜心;甘蓝型油菜Hau CMS;种间杂交;SRAP;花药发育
菜心(Brassica campestris L.ssp.chinensis var.utilis Tsenet Lee)属十字花科芸薹属芸薹种白菜亚种的一个变种,是广东省分布最广、栽培面积最大、复种指数最高的蔬菜种类,其产品质地鲜嫩、口感清甜、风味独特。
菜心为典型的异花授粉作物,杂种优势明显。自交不亲和和细胞质雄性不育是杂种优势利用的主要途径,但自交不亲和系亲本繁殖困难,种子生产成本高。20世纪70年代我国开始菜心细胞质雄性不育的研究,彭谦等[1-2]以甘蓝型油菜Pol CMS湘油A为不育源,以菜心为父本通过种间杂交和连续回交,选育出菜心胞质雄性不育系和相应的保持系,进而培育出早优1号、早优2号、中花杂交菜心。陕西省蔬菜花卉研究所同样利用甘蓝型油菜Pol CMS为母本,用大白菜3411-7为父本,进行杂交和连续回交以及定向培育,育成大白菜异源胞质雄性不育系3411-7A,并将其转育到菜心上,育出了秦薹1号菜心[3]。广西柳州市农业科学研究所利用陕西省蔬菜花卉研究所提供的不育系培育出柳杂1号和柳杂2号菜心。但这些杂交种均未能在生产上大面积推广,主要原因有两个:(1)Pol CMS不育率易受温度变化的影响,很难达到100%不育,给制种带来一定风险;(2)其杂种优势主要表现在叶片、叶柄伸长,侧芽增多,商品性状不佳。
Hau CMS是傅廷栋1999年发现的天然芥菜型油菜细胞质雄性不育类型,蜜腺正常、不育性稳定、败育彻底,目前已通过远缘杂交转移至甘蓝型油菜中[4]。本研究以甘蓝型油菜细胞质雄性不育Hau CMS为不育胞质供体,菜心为轮回亲本进行回交转育,成功选育出菜心Hau CMS不育系,并对各回交世代进行遗传学分析,在此基础上探讨通过远缘杂交将不育基因导入菜心的技术和方法,以期为菜心雄性不育系的转育提供依据。
1 材料与方法
1.1 试验材料
2010年以国家油菜武汉改良分中心提供的新型甘蓝型油菜Hau CMS雄性不育材料为母本、菜心自交系7-2为父本进行杂交,获得种间杂种,2011—2014连续3年用自交系7-2与其回交,在BC4代即获得稳定的菜心雄性不育系7-2A及其相应的保持系7-2B。
1.2 试验方法
1.2.1 植物学性状观察 在植株营养生长期和抽薹开花期,分别观察植株的叶片形状、大小、叶色、苔色、侧芽、抽薹期、开花期等性状。在开花期,观察植株的花器官形态。
1.2.2 花药发育的石蜡切片 选取菜心不育系和保持系主茎顶端花序,按花蕾的着生顺序及大小分级后,FAA固定液固定,70%乙醇保存。按常规石蜡切片程序制作切片:不同浓度酒精逐级脱水,二甲苯透明,浸蜡,包埋,切片,展片,脱蜡,染色,封藏。花蕾横切,切片厚度4~6 μm,1%甲苯胺兰染色[5]。
1.2.3 F1和回交世代SRAP分析 采集幼嫩的菜心叶片,DNA提取采用CTAB法,参照文献[6]的方法并略有改动。
SRAP引物设计原理参照文献[7],引物序列见表1。PCR体系(15 μL)为:100 ng模板DNA、引物各50 ng、1×反应缓冲液、200 μmol/l dNTPs、1.5 mmol/l MgCl2和1U Taq DNA聚合酶(MBI公司)。PCR程序参照文献[7],PCR产物用6%的聚丙烯酰胺凝胶电泳分离。
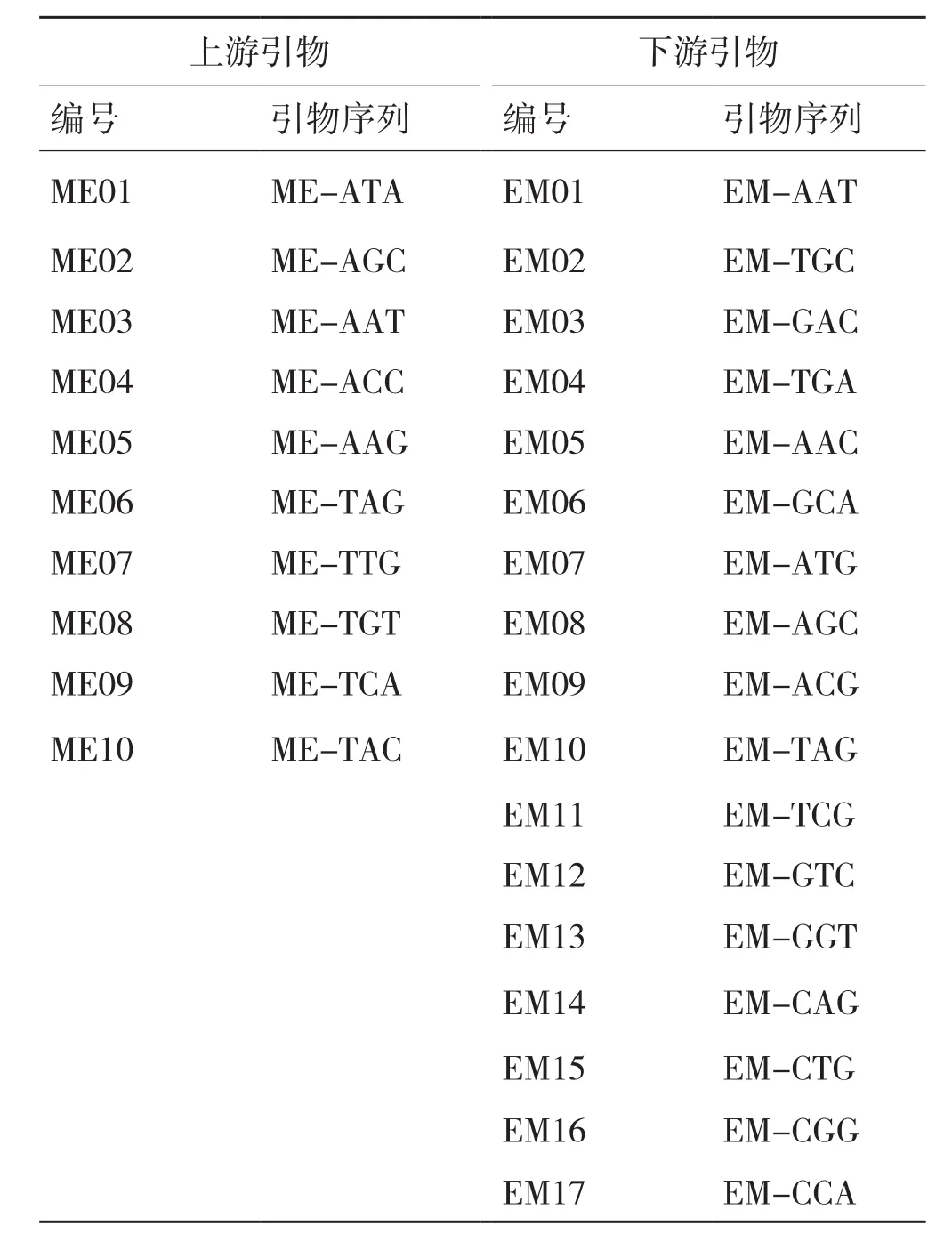
表1 本研究使用的SRAP引物
2 结果与分析
2.1 F1和回交世代的植物学形态
母本为甘蓝型油菜,植株高大,基叶椭圆或长椭圆形,伸长茎叶出现明显的琴状缺刻,薹茎叶披针形、无琴状缺刻、半包茎着生,叶面、薹茎被蜡粉,叶色蓝绿,分枝多,需低温春化,不育度和不育率为100%(图1A,封三)。父本为菜心,早熟,叶片卵圆形,叶色和薹色为油绿色,无蜡粉,侧芽少,无需低温春化(图1B,封三)。通过甘蓝型油菜与菜心杂交获得的F1代和各回交世代未出现育性分离现象,不育率和不育度均为100%。通过田间植物学性状观察发现,F1代有分离,绝大部分单株特征偏向于母本,与甘蓝型油菜相似,但少数单株在叶片形状、大小、颜色、抽薹开花期等方面出现偏向父本的特征,个别单株不需要低温即可抽薹开花。植株苗期不黄化,100%不育且不育性稳定,蜜腺正常,能正常结实(图1C、D,封三)。选择早抽薹的F1单株与菜心回交,得到BC1,BC1代植株表型分离最严重,有的保持母本性状,有的代相似父本,大多数为中间型,植株高矮不均,株型变小,分枝数减少,叶片变小,部分单株叶面和薹茎无蜡粉(图1E、F,封三)。选择无蜡粉,抽薹早的单株与菜心继续回交,BC2代植株表型依然分离严重,但是绝大部分特征偏向于父本,出现和父本极其相似的单株,仅在叶色、薹色和抽薹期方面稍有不同,叶色、薹色哑绿,无光泽,抽薹期较父本推迟7 d左右(图1G,封三)。BC3代植株性状基本趋向轮回亲本,抽薹期略晚,较父本推迟3~5 d(图1H,封三)。选择抽薹期较早,性状与父本最相似的植株与父本回交得到BC4,BC4代植株生长整齐一致,圆叶、柄短、叶色和薹色油绿,侧芽少,不育性稳定、败育彻底,蜜腺正常,结实性好(图1I、J,封三)。
2.2 F1和回交世代SRAP分析
2.2.1 亲本和杂种F1分析 用170对SRAP引物对2个亲本进行筛选,随机选取19对扩增条带清晰、多态性好的SRAP引物对2个亲本、杂种F1和回交后代进行分析。亲本总共扩增出145条带,其中甘蓝型油菜特异性带32条,菜心特异性带12条,甘蓝型油菜和菜心共有的带型为101条。
F1总共扩增出522条带,其中甘蓝型油菜特异性条带181条、占总带型的34.67%,母本特异带缺失12条、占总带型的2.30%,菜心特异性带41条、占总带型的7.85%,甘蓝型油菜和菜心共有的带型为264条、占总带型的50.57%,双亲均没有的新增带型36条、占总带型的6.90%。
2.2.2 回交一代(BC1)分析 BC1总共扩增出488条带,其中甘蓝型油菜特异性带132条、占总带型的27.05%,菜心特异性带43条、占总带型的8.81%,甘蓝型油菜和菜心共有的带型274条、占总带型的56.15%,双亲均没有的新增带39条、占总带型的7.99%,母本特异带缺失62条、占总带型的12.70%。
2.2.3 回交二代(BC2)分析 BC2总共扩增出431条带,其中甘蓝型油菜特异性带68条、占总带型的15.78%,菜心特异性带68条、占总带型的15.78%,甘蓝型油菜和菜心共有的带型为278条、占总带型的64.50%,双亲均没有的新增带型17条、占总带型的3.94%,母本特异带缺失108条、占总带型的25.06%。
2.2.4 回交三代(BC3)分析 BC3总共扩增出384条带,其中甘蓝型油菜特异性带34条、占总带型的8.85%,菜心特异性带62条、占总带型的16.15%,甘蓝型油菜和菜心共有的带型为265条、占总带型的69.01%,双亲均没有的新增带型23条、占总带型的5.99%,母本特异性带缺失159条、占总带型的41.41%。
2.2.5 回交四代(BC4)分析 BC4总共扩增出361条带,其中甘蓝型油菜特异性带15条、占总带型的4.16%,菜心特异性带76条、占总带型的21.05%,甘蓝型油菜和菜心共有的带型为260条、占总带型的72.02%,双亲均没有的新增带型10条、占总带型的2.11%。母本特异性带缺失182条、占总带型的50.42%。
2.3 菜心不育系花药败育特征
2.3.1 菜心保持系花药的发育 显微观察显示,花药形态从最早的花药原基分化至花药各层细胞出现,花粉母细胞被四层体细胞包围:外表皮、内表皮、中层和绒毡层(图3A,封三),花药呈蝴蝶状,4个药隔出现。花粉母细胞经过减数分裂形成四分体,随着包裹四分体的胼胝质降解,小孢子被释放出来形成了单核小孢子(图3B,封三)。随着绒毡层和中层细胞的降解,单核小孢子经过两轮有丝分裂形成一个包含两个精子和1个营养细胞的的成熟花药,随后花药开裂,成熟花粉粒从药隔中释放出来(图3C,封三)。
2.3.2 菜心不育系花药的发育 菜心不育系7-2A花药发育受阻于孢原细胞分化期,孢原细胞不分裂形成造孢细胞和壁细胞(图3D,封三),后期花药向边缘生长并最终发育成由表皮、基本组织和维管束组成的无花粉囊型花药(图3E,封三)。
3 结论与讨论
甘蓝型油菜与菜心远缘杂交,其亲和性良好,人工授粉后不需经过特殊处理即可在田间得到少量种子。可能是因为菜心 (AA=20)与甘蓝型油菜(AACC=38)有一组相同的染色体组AA,这与前人研究的芥菜型油菜(AABB)、甘蓝型油菜(AACC)和白菜(AA)杂交容易得到杂种的结论是一致的。甘蓝型油菜(AACC)是源于芸薹(AA)和甘蓝(CC)的自然复合种。A、C基因组间高度同源,已知功能的编码蛋白质的基因在本质上完全共线性,大约95%的序列是一致的[8-9]。在甘蓝型油菜中,A和C基因组均保留了其完整性,并与它们的二倍体亲本(白菜和甘蓝)非常接近。在进行白菜与甘蓝型油菜的比较作图时,发现两个物种中的多数A基因组上的位点都是保守的[10]。SRAP标记对F1和回交世代遗传变异的分析结果也证实了这一点,杂种和回交世代父母本共有的条带比率占50%以上,BC4群体父母本共有的条带比率为72.02%。F1和回交世代父母本都没有的新增条带比率维持在5%左右,说明在甘蓝型油菜(AACC)和菜心(CC)远缘杂交和回交过程中发生了染色体交换重组。BC3母本特异性条带占总带型的8.85%,材料的田间表型较整齐一致,基本与轮回亲本的性状相同,这表明AACC染色体组物种与AA染色体物种远缘杂交经少数几代回交即可迅速获得后代性状相对稳定的材料。
关于芸薹属作物细胞质雄性不育的花药发育,前人做了很多研究,Pol CMS的花药发育受阻于孢原细胞分化期,没有花粉囊形成;Ogu CMS的败育发生在四分体至单核花粉时期,小孢子难以从四分体中释放出来[11];白菜Hau CMS不育系6w-9605A的花药败育有两种方式,2/3的花药发育受阻于孢原分化时期,不形成花粉囊,1/3的败育花药可形成花粉囊,小孢子受阻于单核靠边期[11];本研究菜心Hau CMS不育系7-2 A的败育时期为孢原细胞分化期,没有造孢细胞和壁细胞的分化,属于无花粉囊型,这与芥菜Hau CMS不育系0912A的败育方式类似[12]。而芥菜型油菜不育系00-6-102A的花药败育发生在雄蕊原基分化期,典型特征是雄蕊原基偏离正常的分化轨道,形成花瓣原基,无孢原细胞及花粉囊的形成,6枚雄蕊发育成小花瓣[13]。说明Hau CMS不育胞质在不同的核背景下,其花药败育方式会发生改变,这与王涛涛等[14]的研究结果一致,原因可能是不同的核基因对Hau CMS的调控作用不同。
参考文献:
[1] 彭谦,李汝松,吕英华,等.菜心雄性不育研究初报[J].中国蔬菜,1989(1):1-3.
[2] 刘自珠,张华,刘艳辉,等.菜心胞质雄性不育系的选育及利用[J].广东农业科学,1996(5):13-15.
[3] 赵利民,柯桂兰,宋胭脂.胞质雄性不育系配制的菜薹新品种“秦薹1号”[J].园艺学报,2002,29 (4):399.
[4] Wan Z J,Jing B,Tu J X,et al.Genetic characterization of a new cytoplasmic male sterility system(hau)in Brassica juncea and its transfer to B.napus.[J].Theor Appl Genet,2008,116:355-362.
[5] 王灶安.植物显微技术[M].北京:中国农业出版社,1992.
[6] Doyle J J,Doyle J L.Isolation of plant DNA from fresh tissue[J].Focus,1980,12:13-15.
[7] Li G,Quiros C F.Sequence-related amp lified polymorphism(SRAP),a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica[J].Theoretical and Applied Genetics,2001,103:455-461.
[8] Rana D,Boogaart T,O'Neill C M,et al.Conservation Of the microstructure of genome segments in Brassica napus and its diploid relatives[J].The Plant Journal,2004,40 (5):725-733.
[9] Cheung F,Trick M,Drou N,et al. Comparative analysis between homoeologous genome segments of Brassica napus and its progenitor species reveals extensive sequence-level divergence[J].The Plant Cell,2009,21(7):1912-1928.
[10] Hoenecke M,Chyi Y S.Comparison of Brassica napus and B.rapa genomes based on restriction fragment length polymorphism mapping[A].Proceedings of the Eighth International Rapeseed Congress GCIRC [C].Saskatoon,Canada,1991:1102-1107.
[11] 余凤群,傅廷栋.甘蓝型油菜几个雄性不育系花药发育的细胞形态学研究[J].武汉植物学研究,1990,8(3):209-216.
[12] 施展,万正杰,徐跃进,等.大白菜新型细胞质雄性不育系6w-9605A 的育性鉴定和花药败育的细胞学观察[J].植物科学学报,2012,30(1):49-54.
[13] Wan Z J,Tan Y B,Shi M H,et al.Interspecific introgression of male sterility from tetraploid oilseed Brassica napus to diploid vegetable B.rapa through hybridisation and backcrossing[J].Crop & Pasture Science ,2013,64,52-659.
[14] 万正杰.芥菜型油菜细胞质雄性不育hau CMS 的研究[D].武汉:华中农业大学,2008.
[15] 王涛涛,蔡晓峰,张俊红,等.芥菜型油菜雄性不育系与甘蓝远缘杂交胚培养及早代育性鉴定[J].园艺学报,2010,37(10):1661-1666.
(责任编辑白雪娜)
Trans-breeding Hau cytoplasmic male sterile line in Chinese flowering cabbage
ZHANG Yan,CHEN Han-cai,LI Gui-hua,LI Ting-yao
(Vegetable Research Institute,Guangdong Academy of Agricultural Sciences/Guangdong Key Laboratory for New Technology Research of Vegetables,Guangzhou 510640,China)
Abstract:Using the Hau cytoplasmic male sterile line of Brassica napus as sterile material,interspecific cross and successive backcrossing with 7-2B were conducted to transfer the Hau CMS to Chinese flowering cabbage.In the course of acquiring cytoplasmic male sterility lines,the morphology,molecular biology and cytology in various backcross generations were studied.The results showed that,the morphological characteristics of F1plants were segregated and more like the female parent,the progeny became stable and more similar to Chinese flowering cabbage by BC3.Analysis by SRAP indicated that 2.3% to 50.42% of the total bands eliminated from F1to BC4 were female parent specific,and 4.16% of female parent were remained in the BC4 plants.The dissection of anther development indicated that the anther abortion occurred at the early stage of stamen sporogonium differentiation,and no butterfly anther appeared.
Key words:Chinese flowering cabbage;Hau cytoplasmic male sterile line; interspecific hybridization;sequence-related amplified polymorphism(SRAP); anther development
中图分类号:S634.5
文献标识码:A
文章编号:1004-874X(2016)01-0030-04
收稿日期;2015-10-02
基金项目:广东省科技计划项目(2012A020100006,2012B020303004,2014A020208144);广东省自然科学基金项目(2014A030313565)
作者简介:张艳(1981-),女,博士,助理研究员,E-mail:18520164058@163.com
通讯作者 :陈汉才(1962-),男,研究员,E-mail: chen6206@21cn.com
