贵州特有植物贵州槭的种群结构及动态研究
2016-06-16刘海燕杨乃坤邹天才黄丽华汪建文
刘海燕, 杨乃坤, 邹天才, 黄丽华, 汪建文
( 1. 贵州省植物园, 贵阳 550004; 2. 贵阳中雄林业生态工程勘察设计有限公司, 贵阳 550004; 3. 贵州科学院, 贵阳 550001 )
贵州特有植物贵州槭的种群结构及动态研究
刘海燕1, 杨乃坤2, 邹天才3*, 黄丽华3, 汪建文3
( 1. 贵州省植物园, 贵阳 550004; 2. 贵阳中雄林业生态工程勘察设计有限公司, 贵阳 550004; 3. 贵州科学院, 贵阳 550001 )
摘要:贵州槭(Acer guizhouense)属常绿阔叶乔木,仅分布于贵州省黄平县新州镇海拔770~1 000 m有山峰屏障的常绿阔叶落叶混交林中,颇具科研和经济价值,研究其种群结构特征和数量动态变化对于探索特有植物保护生物学具有重要意义。该研究对贵州槭疑似分布区开展普查,并在密集分布区设置样地进行详查,分析其种群结构与数量动态的变化规律。结果表明:(1)贵州槭的自然分布区极为狭窄,雷坪分布区约0.3 km2,总计约2 600株,其中密集分布区0.03 km2,平均种群密度78 800株/km2,稀疏分布区0.27 km2,平均种群密度≤1 500株/km2;在东坡村飞云崖风景区有约0.05 km2的稀疏分布,且在雷坪至东坡两分布区之间没有发现自然分布;采用SAFE指数和IUCN的标准评价,贵州槭属于濒危物种(EN)。(2)贵州槭幼树占种群的比重达63.23%,种群密度大小为幼树>中龄树>成年树,种群结构为增长型;种群死亡率曲线和消失率曲线分别在Ⅰ龄期和Ⅸ龄期出现2个高峰,在Ⅲ-Ⅷ龄期间和Ⅹ-Ⅺ龄期间出现2个低谷,种群存活曲线趋于Deevey-Ⅲ型特征;(3)人为采伐和生境遭破坏造成的植株死亡或者丧失增加是导致贵州槭种群数量稀少的主要原因,在加强就地保护的同时,开展人工育苗迁地应用栽培是现实而有效的合理保护措施。
关键词:贵州槭, 种群结构, 静态时间生命表, 数量动态, 特有分布
种群生态学是在个体、种群、群落中,以种群为研究对象的生态学分支,在现代生态学研究的各层次之间起着联结和枢纽作用;植物种群的年龄结构及数量动态特征,通常是物种种群过去发生发展到当前现状过程的客观反映,能够为物种未来动态变化的进一步研究提供基础(易雪梅等, 2015; Holeksa et al, 2007)。种群结构变化、种群生命表和种群存活曲线是研究种群数量动态变化的核心内容(闫桂琴等, 2001; 徐学红等, 2005; 张桥英等, 2007、2008)。因此,采用种群结构分析和动态评价的方法研究贵州槭的种群特征对贵州特有植物的保护生物学研究具有重要意义(张亚芳等, 2015)。
贵州槭(Acerguizhouense)属槭树科(Aceraceae)槭属(AcerL.)常绿阔叶乔木,成年树高8~12 m、树干直径20~40 cm,是贵州科学院李永康教授于1987年发现并定名的新种(李永康, 1987)。仅分布于贵州省黄平县海拔770~1 000 m有山峰屏障的常绿落叶阔叶混交林中,生境温暖湿润,资源稀少。该物种自发现以来,邹天才、杜娟等开展了迁地保护试验、种子和叶片化学成分分析研究(邹天才等,2007;杜娟等, 2011),但在生态学方面尚缺乏详实资料,为探明其植物种群分布的受限机理,本文以贵州槭原生境区的种群为研究对象,对种群结构、种群动态等方面开展了实地调查和种群动态特征分析,为进一步研究贵州槭植物资源的有效保护和合理开发利用提供科学依据。
1研究区域与研究方法
1.1 调查区概况
调查区位于黄平县新州镇雷坪村(地理位置为26°59′42″ E、107°56′34″ N,海拔880 m),东坡村(地理位置为26°56′56″ E、107°59′22″ N,海拔790 m),四季温暖湿润,冬无严寒、夏无酷暑,年平均气温14.7~15.1 ℃,年均降雨量1 200 mm,属亚热带季风性湿润气候。调查区植被以次生常绿阔叶落叶林为主,用材林木遭受人为采伐严重,群落结构单一,伴生植物种类稀少。
1.2 研究方法
1.2.1 野外调查及统计方法2015年4月6-20日,在黄平县新州镇西北部东坡村飞云崖—白塘村—雷坪村一带的贵州槭疑似分布区进行野外实地踏查、数据观测和统计。根据踏查和观测结果在位于雷坪村的贵州槭密集分布区(种群密度≥10 000 株/km2,生活史较为完整)和稀疏分布区(种群密度<10 000株/km2,生活史不完整,龄级缺级现象严重)分别选2块具有代表性的样地开展详查,依据调查区的生态环境和地形地貌特征,沿着坡向和垂直于坡向选择4个样地,每样地设置3个20 m × 20 m的样方, 样地面积共计4 800 m2。调查样方内乔木树种数量,对每株贵州槭的高度、胸径、冠幅进行逐一测量记录。在每个20 m × 20 m样方中再设置1个5 m × 5 m的小样方,详查其灌木层盖度、植株高度、植物种类及其比例,同时在每个灌木样方的中心位置划出1 m × 1 m的小样方测定草本植物组成、高度和盖度。资源量统计方法如下:
总资源量=密集分布区资源量+稀疏分布区资源量;密集分布区资源量=密集分布区平均株数(km2)×密集分布区面积(km2);稀疏分布区资源量=稀疏分布区平均株数(km2)×稀疏分布区面积(km2)。
采用Clements et al (2011) 提出的物种抗灭绝能力(species’ ability to forestall extinction, SAFE)指数和IUCN的评价标准(IUCN,2001)来评价贵州槭物种当前的濒危等级。
SAFE指数= log (N)-log (MVP)
式中,N是该物种在已知分布区的种群数量,MVP是根据该物种的种群参数和遗传参数估算的最小可容纳种群数量经验值,此处设定为3 000(Traill et al,2007;Richard et al,2014)。
1.2.2 径级划分和静态生命表编制种群中植株的胸径大小是反映植物种群结构最易测定和值得信赖的有效因子,在研究过程中通常采用径级结构来代替年龄结构,并取得了良好效果(沈年华等,2015)。根据相同环境中树木的龄级和径级的变化具有一致性的特点(Frost et al, 2000),故采用径级代替龄级的方法来分析贵州槭的种群结构(张亚芳等, 2015;易雪梅等,2013)。据实测数据,结合贵州槭的生物学特性,将贵州槭种群按胸径大小每相差5 cm为1个径级划分为12个径级: 0~5 cm 为第Ⅰ径级,5~10 cm 为第Ⅱ径级,10~15 cm为第Ⅲ径级,逐级增加到55~60 cm为第Ⅻ径级;第Ⅰ径级对应第1龄级,第Ⅱ径级对应第2龄级,依此类推,统计各径级贵州槭的个体数量。
1.2.3 种群生存分析运用种群生存率函数S(x)、累计死亡率函数F(x)、死亡密度函数f(x)和危险率函数λ(x))方法统计调查测算值、绘制曲线,并总结贵州槭的种群结构特征和动态变化规律(Kauffman et al, 2006; 项小燕, 2016),其计算公式如下:
S(x)=S1·S2·S3…Sx(Sx为存活率)
F(x)=1-S(x)
f(x)=(Sx-1-Sx)/hx(hx为龄级宽度)
λ(x)=2(1-Sx)/[hx(1+Sx)]
1.2.4 种群动态量化为较客观准确地量化评价贵州槭的种群动态特征,充分考虑种群内两相邻龄级彼此间个体数量变化动态指数(Vx)和种群整体的年龄结构动态变化指数(Vpi)以及未来外部干扰情况下种群的年龄结构变化指数(Vpi'),引入如下公式计算(陈晓德, 1998; 刘普辛, 2011; 吕海英等, 2014; 孟广涛等,2008; 韩路等,2014; 刘海燕等,2016):
式中,Vx为两相邻龄级彼此间个体数量变化动态指数,Vpi为种群整体的年龄结构动态变化指数,Sx、Sx +1分别表示第x和第x+1龄级的个体数,k表示种群最大年龄级max/min(……)取括号中数列极大值/小值,Vx(Vpi)≤1、Vx(Vpi取正、负、零值时分别反映出贵州槭种群两相邻年龄级个体数量(或整个种群年龄结构)的增长、衰退、稳定等结构的动态关系(于倩等, 2008);尚若在未来某一时期受到外部干扰,Vpi′ 就与K值和Sx值相关,即K、Sx对未来外部干扰存在着“稀释效应”,可由K、Sx依据条件概率法则计算其最大概率值,修改为Vpi′式(陈晓德, 1998),Vpi′指标是种群结构动态对于随机干扰影响敏感性大小的特征反映。
2结果与分析
2.1 贵州槭分布区域、种群密度及数量
调查发现,贵州槭自然分布于黄平县新州镇海拔770~1 000 m有山峰屏障的常绿阔叶落叶混交林中,较为集中地分布在雷坪村的自然沟谷地带,在东坡村的飞云崖风景区有零星分布,但在雷坪村~东坡村间隔区间没有发现有自然分布。根据贵州槭自然分布区域地形设置调查样地位置得图1,图1中红色线段圈围部分为自然分布区,蓝色点线圈围部分为密集分布区,A、B、C、D为调查样地的设置标记点。通过实地踏查,发现贵州槭雷坪自然分布区约0.03 km2(500 m × 600 m),其中密集分布区约0.03 km2(250 m × 80 m+250 m × 40 m),稀疏(零星)分布区0.27 km2;另外,在东坡村的飞云崖风景区有约0.05 km2的稀疏分布,且生活史不完整。调查统计结果显示:贵州槭雷坪自然分布范围内存活植株总计2 600余株,其中密集分布区内有2 300余株,平均种群密度78 800 株/km2(72 100株/km2~85 400株/km2);稀疏分布区内有300余株。分布区域极其狭窄,种群分布极不均匀,最密集区域>85 400株/km2,最稀疏区域<1 500 株/km2;东坡村飞云崖风景区分布范围内存活植株总计约550株,平均种群密度约11 000 株/km2。
根据SAFE指数统计得表1,雷坪村分布的贵州槭种群指数为-1<-0.187<0,趋近SAFE指数分布的中位值,而东坡村飞云崖分布的贵州槭SAFE指数为-0.862,趋于-1。根据Clements et al(2011)将IUCN红色名录濒危等级中的灭绝(EX)、极危(CR)、濒危( EN)、易危(VU)、近危(NT)都合并为SAFE指数的“Threatened”(受威胁的),可将贵州槭划分为受威胁物种。同时,根据IUCN的评价标准,由于贵州槭分布区面积狭小(<500 km2),空间分布片段化(严重分割为已知仅有的2个地点),且种群破碎化,成熟个体数仅占整个种群的9.26%,因此,贵州槭可划分为濒危物种( EN)范畴。

图 1 贵州槭自然分布地点及分布区大小Fig. 1 Location of natural distribution and the distribution size of Acer guizhouense
2.2 贵州槭的种群径级分布及年龄结构
图2显示,其种群径级结构呈倒“J”形,Ⅰ龄级个体最多,随后呈幂函数递减。4 800 m2样地内共调查存活个体378株,其中胸径0~5.0 cm的幼龄植株239株,占整个种群的63.23%,5.0~20.0 cm的中龄植株104株,占整个种群的27.51%,而胸径>20.0 cm的成年树仅有35株,占整个种群的9.26%。胸径≤20.0 cm时,种群的个体植株数量随着径级的增加而急剧减少,当胸径>20.0 cm时,这个趋势变慢,这说明贵州槭种群的年龄结构属于增长型。

表 1 贵州槭的种群数量及SAFE指数
2.3 贵州槭种群静态生命表
表2结果显示,贵州槭种群的生命期望在前3个龄级阶段呈递升趋势,Ⅳ龄级阶段开始逐渐降低,之后在Ⅹ龄级阶段又有一个回升,随后急剧下降。生命期望最高值出现在Ⅲ龄阶段,平均生命期望值为2.70。其次是Ⅹ龄级阶段,平均生命期望为2.50。平均生命期望变化规律大体上呈现“低-高-低”模式。贵州槭种群的存活率变化规律与生命期望基本相同,亦呈“低-高-低”的变化模式。这表明贵州槭种群中幼树的绝对数量虽然较多,但由于在Ⅰ、Ⅱ龄级阶段有较高的损耗和死亡, 使得这两个龄级的存活率大大降低。

图 2 贵州槭的种群径级结构图Fig. 2 Histogram of DBH classes of Acer guizhouense population

龄级Ageclass径级DBHclass(cm)xAxaxlxln(lx)dxqxLxTxexSxKxⅠ0.0-5.02.552391000.006.91732.220.73633.891075.311.080.271.32Ⅱ5.0-10.07.5564267.785.59171.550.64182.01441.421.650.361.02Ⅲ10.0-15.012.552396.234.5725.100.2683.68259.412.700.740.30Ⅳ15.0-20.017.551771.134.2620.920.2960.67175.732.470.710.35Ⅴ20.0-25.022.551250.213.9216.740.3341.84115.062.290.670.41Ⅵ25.0-30.027.55833.473.5112.550.3827.2073.222.190.630.47Ⅶ30.0-35.032.55520.923.044.180.2018.8346.032.200.800.22Ⅷ35.0-40.037.55416.742.824.180.2514.6431.381.880.750.29Ⅸ40.0-45.042.55312.552.538.370.678.3718.831.500.331.10Ⅹ45.0-50.047.5514.181.430.000.004.1810.462.501.000.00Ⅺ50.0-55.052.5514.181.430.000.004.186.281.501.000.00Ⅻ55.0-60.057.5514.181.434.181.002.092.090.500.001.43
注: x. 单位时间内的龄级中值(cm); Ax. 龄级宽度(以5cm为1个龄级); ax. 在x龄级内存活的个体数; lx.x龄级开始时的标准化存活个体数(以1 000为基数,lx=ax/a0×1 000) ; dx. 从x到x+1间隔期内标准化死亡数(dx=ax-ax+1); qx. 从x到x+1龄级间隔期间个体死亡率(qx=dx/lx); Lx. 从x到x+1龄级间隔期间的平均存活个体数(Lx=(lx+lx+1)/2); Tx.x龄级及其以上各龄级的存活总数(Tx=ΣLx);ex. 进入x龄级个体的期望寿命(ex=Tx/lx);Sx. 存活率(Sx=lx+1/lx);Kx. 致死力或消失率,Kx=ln(lx)-ln(lx+1)。
Note:x.Midpointage(cm);Ax.Widthofage(cm);ax.Numberofsurvivalindividuals;lx.Proportionindividualssurvivingfromthebeginningtoagex, lx= ax/a0×1000;dx.Numberofdeadindividualsfromagextox+1, dx=ax-ax+1;qx.Mortalityratefromxtox+1, qx= dx/lx;Lx.Meannumberofsurvivalindividualsfromagextox+1, Lx= (lx+lx+1)/2;Tx.Totalnumberofsurvivalindividualsfromagex, Tx=ΣLx;ex.Lifeexpectancyatagex, ex= Tx/lx;Sx.Age-specificsurvival, Sx= lx+1/lx;Kx.Agespecificmortality, Kx=ln(lx)-ln(lx+1).
2.4 贵州槭种群生存力
2.4.1 贵州槭种群存活曲线、死亡率曲线把调查数据统计绘制出贵州槭的种群存活曲线、死亡率曲线(qx)和消失率(Kx)曲线得图3。从图3:A可以看出,贵州槭种群的存活曲线接近Deevey类型划分的Ⅲ型特征表明该物种幼树绝对数量虽然较多,但幼龄级阶段个体死亡率较高,分别为73.22%和64.06%,而存活率为26.78%和35.94%。图3:B显示,在贵州槭的整个生命过程中,死亡率和消失率均出现2次高峰和2次低谷,第一次高峰出现在Ⅰ龄级阶段,随后的两个龄级死亡率逐渐下降,在Ⅲ-Ⅷ龄级阶段出现第一个低谷,紧接着在Ⅸ龄级阶段出现第二次高峰后死亡率和消失率急剧下降到0,到Ⅹ-Ⅺ龄级阶段时出现第二次低谷,死亡率和消失率均为0,之后又开始急剧上升。

表 3 贵州槭种群两相邻年龄级个体数量变化的动态量值(Vx,%)及种群年龄结构的动态指数 (Vpi,%)

图 3 A. 贵州槭种群的存活曲线和B. 贵州槭种群的死亡率(qx)曲线及消失率(Kx)曲线Fig. 3 Survival curve (A) and mortality rate(qx) and hazard rate(Kx)(B) of Acer guizhouensis population
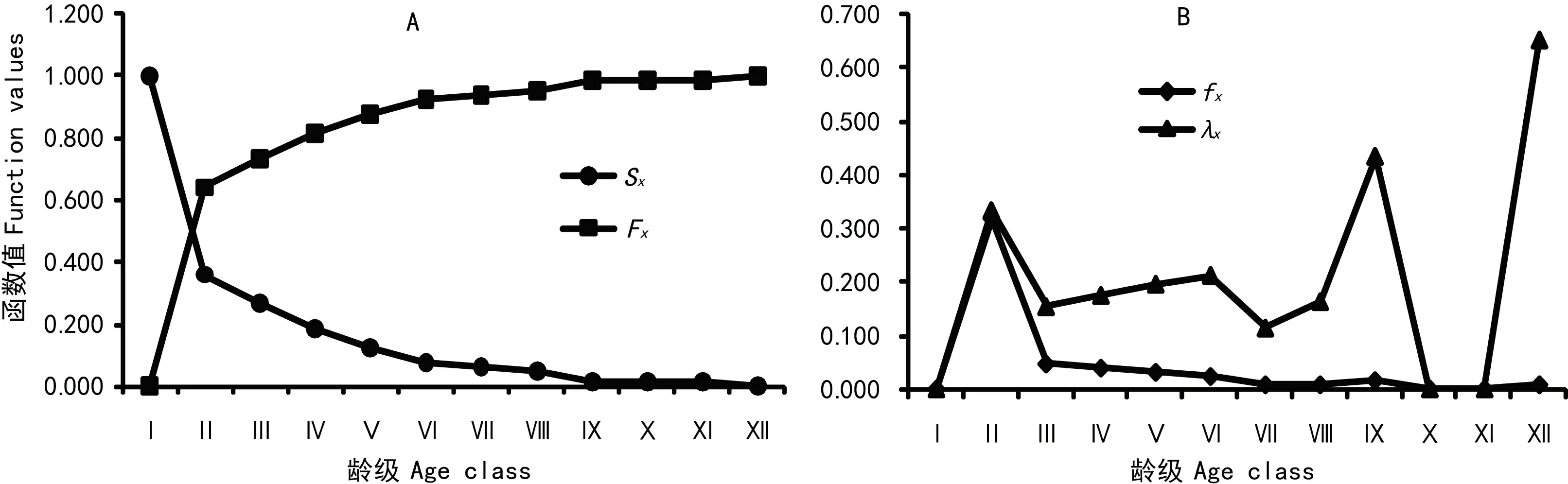
图 4 贵州槭的种群生存曲线Fig. 4 Survival function curve of the Acer guizhouense population
2.4.2 贵州槭的种群生存曲线把调查数据引入种群生存率函数S(x)、累计死亡率函数F(x)、死亡密度函数f(x)和危险率函数λ(x))计算绘制出图4。在图4:A中,贵州槭的种群生存率随着龄级的增大呈显单调下降趋势,但其累计死亡率则相反,且生存率和累计死亡率的升降在Ⅰ至Ⅳ个龄级表现明显;第Ⅳ龄级后,种群生存率和累计死亡率的升降趋于平缓,分别呈均匀的下降、上升,种群趋于稳定状态。图4:B显示出,贵州槭种群死亡密度和危险率都在Ⅱ龄级出现一个峰值,随即在Ⅲ龄级出现一个低谷,Ⅲ-Ⅷ龄级阶段略有起伏但相对平缓,之后危险率函数急剧上升,死亡密度函数依然逐级平缓下降。这表明贵州槭的种群动态存在着前期减少、中期稳定、后期衰退的特点,并且与其种群存活曲线、死亡率曲线和消失率曲线的特征表现一致。
2.5 贵州槭数量动态指数
表3显示,贵州槭种群发育过程中各个龄级为下一龄级的贡献潜力在整体呈下降趋势下存在上下波动的变化。Ⅰ-Ⅲ龄级下降趋势明显,Ⅲ-Ⅵ龄级略有上升,之后起伏较大,说明贵州槭种群中幼苗补充仍然充足,即由幼苗向幼树、小树过渡中呈现明显的增长型结构。种群的Vpi分析结果为正值,表明贵州槭种群呈现增长型的总体结构;同时,种群龄级结构动态指数(Vpi′)大于0,但却远小于Vpi值,表明贵州槭种群对环境变化敏感,种群动态变化受环境因素的影响较大,其主要原因可能是由于土著居民伐木取材砍伐树木或其他经济建设活动的影响所致。
3讨论与结论
3.1 贵州槭分布狭限、物种濒危,种群结构特征及动态变化的研究是其保护生物学的基础
贵州槭喜阴生,在溪流沟谷沿边分布或在阴湿环境中呈小块状分布,自然分布区域狭窄、分布区总面积约0.35 km2,仅仅在黄平县新州镇的雷坪村溪流沟谷地带和东坡村的飞云崖风景区阴湿环境中自然分布,雷坪村的自然分布区约0.30 km2(密集分布区≤0.03 km2),东坡村飞云崖风景区约有0.05 km2稀疏分布,且在雷坪至东坡两分布区之间没有发现有自然分布。贵州槭种群密度的大小为幼树>中龄树>成年树,种群结构表现为增长型;种群死亡率曲线和消失率曲线分别在Ⅰ龄期和Ⅸ龄期出现2个高峰,在Ⅲ-Ⅷ龄期间和Ⅹ-Ⅺ龄期间出现2个低谷,种群存活曲线呈凹形,具有明显的Deevey-Ⅲ型特征,表明贵州槭在幼苗生长期的生态环境脆弱、有较高的死亡率,当生长到6~10龄级期间的种群则较为稳定,但在生长10龄级以后的贵州槭材用价值高、人为采伐的几率加大,植株丧失率或死亡率又逐步增加。近些年来,由于林木采集和开山修路等人为经济建设活动的频繁发生,严重干扰了贵州槭的群落结构及其生态环境,具有完整生活史的分布面积不足0.03 km2,形成了贵州槭单优群落,伴生物种的种类数量和个体数量均为稀少,人为砍伐用材林木和生态环境遭破坏导致了种群数量和分布面积逐步减少,物种趋于濒危。物种的种群数量是确定小种群物种濒危等级的关键(Li et al, 2011),保护生物学需要根据物种充分的种群动态信息来决策,对贵州槭种群现状进行的调查、种群动态变化和物种濒危程度的评价能够更好地描述其物种的自然资源现状。因此,深入开展贵州槭种群结构特征及动态变化的研究,可为贵州槭特有植物资源的保护生物学研究与决策提供科学依据。
3.2 贵州槭种群接近增长型结构,生态环境遭破坏和人为采集是其物种濒危的主要原因
种群结构既反映了种群个体在空间和时间上的配置数量和发展动态,也体现出种群与环境之间的相互关系及其不同个体之间的相互作用(康华靖等, 2007)。种群的静态生命表和生存函数分析可以反映出种群不同生活史阶段的动态特征及其与环境之间的相互关系,可作为研究种群动态规律和经营管理的主要参考。通过贵州槭种群年龄结构和静态生命表的分析表明,种群呈现为幼、中个体数量较多,老龄个体数量少,种群存活曲线为Deevey-Ⅲ型,接近于增长型结构;采用种群数量动态的分析表明贵州槭的种群结构在整体上为增长型,但对环境因素的变化比较敏感,种群动态易受环境的影响。这说明在环境条件变化不大且保证没有人为干扰的状态下,种群的幼龄个体补充已可以满足其自身种群的生存繁衍。
植物地理分布的受限通常有两个原因:一是内部因素,即植物的生物学特性,包括植物繁殖特性、扩散和生态适应性;二是外界因素,即生态环境因子的影响和人为干扰导致已成活的个体的死亡率增加(Yoshida et al, 1999; 邹天才, 2001)。贵州槭属于常绿阔叶乔木,雌雄同株,果实为翅果,种子主要靠风力传播。本研究实地调查的两个贵州槭自然种群样地中均有大量幼树出现,但在4 800 m2的样方内调查所得的378株贵州槭个体中仅有139 株胸径5 cm(Ⅰ龄级)以上的成活个体,加之调查过程中发现有大量被砍伐的树桩,这说明种子繁殖和新增幼苗数量并不是贵州槭种群更新的限制性因素,而本土居民伐木取材、毁林造田、以及三施高速和余凯高速的修建等人为经济活动不可避免造成的人为干扰和生态环境破坏致使大量贵州槭植株遭受砍伐和践踏性死亡,进而导致了贵州槭种群由Ⅰ龄级向Ⅱ龄级以及Ⅱ龄级向Ⅲ龄级过渡阶段较高的死亡率是其种群数量稀少和物种濒危的主要原因。
3.3 坚持保护与开发利用相结合,推进贵州槭特有植物应用栽培具有现实意义
贵州槭是贵州喀斯特山区典型性的特有稀有植物,颇具用材林树种和景观植物的应用价值,一直以来遭遇掠夺性采伐,当前,黄平县政府已经在黄平县飞云崖风景区建立了自然保护小区,贵州省植物园对贵州槭进行了引种栽培和迁地保育试验等,使贵州槭得到了一定程度的保护,但系统性的有效保护和合理开发利用措施仍然欠缺。鉴于目前贵州槭植物资源量稀少和分布面积狭窄的现状,建议在贵州槭的自然分布区域建立起就地保护区,维持贵州槭种群的自然繁殖演替,减少人为经济活动和工程建设的影响,以卓有成效地增强贵州槭的就地保护功能和效果;与此同时,深入研究贵州槭人工育苗技术和迁地栽培技术,把贵州槭设计应用于生态景观工程和生态林业建设,在绿化工程中应用栽培,开展贵州槭特有植物生产应用栽培的试验示范,建立和扩大贵州槭应用栽培区域及其种群数量,使之在有效保护特有植物资源的同时,又能够合理开发利用植物资源造福于民,有效推进特有植物资源优势转化为地区特色经济优势。
参考文献:
CHEN XD, 1998. A study on the method of quantitative analysis for plant population and community structural dynamics [J]. Acta Ecol Sin, 18(2): 214-217. [陈晓德, 1998. 植物种群与群落结构动态量化分析方法研究 [J]. 生态学报, 18(2): 214-217.]
Clements GR, Bradshaw CJA, Brook BW, et al, 2011. The SAFE index: using a threshold population target to measure relative species threat [J]. Fron in Ecol Environ, 9: 521-525.
DU J, LIU HY, ZOU TC, 2011. Analysis on main chemical components of seed and leaves ofAcerguizhouenseY. K. Li [J]. Seed, 30(4): 94-96. [杜娟, 刘海燕, 邹天才, 2011.贵州槭种子和叶片主要化学成分分析 [J]. 种子, 30(4): 94-96.]
FROST I, RYDIN H, 2000. Spatial pattern and size distribution of the animal-dispersedQuercusruburin two spruce-dominated forests [J]. Ecoscience, 7(1) : 38-44.
HOLEKSA J, SANIGA M, SZWAGRZYK J, et al, 2007. Altitudinal variability of stand structure and regeneration in the subalpine spruce forests of the Pol’ana Biosphere Reserve, Central Slovakia [J]. Eur J For Res, 126(2): 303-313.
IUCN (World Conservation Union), 2001. IUCN Red List Categories and Criteria: Ver 3.1[M]. IUCN Species Survival Commission.
HAN L, WANG JQ, WANG HZ, et al, 2014. The population structure and dynamics ofPopuluseuphraticaat the upper reaches of the Tarim River [J]. Acta Ecol Sin, 34(16): 4 641-4 651. [韩路,王家强,王海诊,等,2014,塔里木河上游胡杨种群结构与动态 [J]. 生态学报, 34(16): 4 640-4 651.]
KANG HJ, CHEN ZL, LIU P, et al, 2007. The population structure and distribution pattern ofEmmenopteryshenryiin Dapanshan Natural Reserve of Zhejiang Province [J]. Acta Ecol Sin, 27(1): 389-396. [康华靖, 陈子林, 刘鹏, 等, 2007. 大盘山自然保护区香果树种群结构与分布格局 [J]. 生态学报, 27(1): 389-396. ]
KAUFFMAN MJ, MARON JL, 2006. Consumers limit the abundance and dynamics of a perennial shrub with a seed ban [J]. Am Nat, 168(4): 454-470.
LI B, YANG C, LIN P, 2000. Ecology. [M]. Beijing: Higher Education Press:43-44. [李博,杨持,林鹏, 2000. 生态学 [M],高等教育出版社:43-44.]
LI CL, JIANG ZG, PING XG, et al, 2011. Current status and conservation of the endangered Przewalski’s gazelleProcapraprzewalskii, endemic to the Qinghai-Tibetan Plateau, China [J]. Oryx, 46: 145-153.
LI YK, 1987. Two new species ofAcerfrom Guizhou [J]. Guihaia, 7(3): 211-213. [李永康, 1987. 贵州槭属二新种 [J]. 广西植物, 7(3): 211-213.]
LIU PX, 2011. Study on population structure and dynamics ofPopuluseuphraticain the middle and lower reaches of the Shule River Basin Oasis, Hexi Corridor [J]. J Nat Resour, 26(3): 429-439. [刘普辛, 2011. 疏勒河中下游绿洲胡杨种群结构与动态研究 [J]. 自然资源学报, 26(3): 429-439.]
LÜ HY, WANG XA, LI J, et al, 2014. Structure and dynamics of China Rare PlantAmmodendronbifolium(Pall. )Yakovl Natural population [J]. Acta Bot Boreal-Occid Sin,34(1): 177-183. [吕海英, 王孝安, 李进, 等, 2014. 珍稀植物银砂槐中国分布区的种群结构与动态分析 [J]. 西北植物学报, 34(1): 177-183.]
LIU HY, YANG NK, LI YY,et al, 2016. The population structure and dynamic analysis of the rare and endangered plantCamellialongistyla[J], Plant Sci J,34(1):89-98. [刘海燕,杨乃坤,李媛媛等, 2016. 稀有濒危植物长柱红山茶种群特征及数量动态研究.植物科学学报,34(1):89-98.]
RICHARD BP, MA KP, JIANG ZG, 2014. Essentials of conservation biology (15th ed) [M]. Beijing: Science Press, 6: 234-234. [Richard B. Primack,马克平,蒋志刚, 2014. 保护生物学 [M]. 北京: 科学出版社, 6: 234-234.]
MENG GT, CHAI Y, FANG XJ, et al, 2008. A preliminary study of population dymamics ofPseudotsugasinensispopulation in Northeastern Yunnan [J]. J NW For Univ, 23(6): 54-59. [孟广涛,柴用,方向京,等, 2008. 滇东北黄山种群数量动态的初步研究 [J]. 西北林学院学报, 23(6): 54-59.]SHEN NH, LI CL, TANG GG, 2015. Population dynamics ofQuercusvariabilison Mount Zijinshan [J]. J Fujian For Sci Technol, 42(3): 104-106. [沈年华, 李传磊, 汤庚国, 2015. 南京紫金山栓皮栎种群动态 [J]. 福建林业科技, 42(3): 104-106.]
Traill LW, Bradshaw CJA, Brook BW, 2007. Minimum viable population size:a meta-analysis of 30 years of published estimates [J]. Biol Conserv, 139:159-166.
XIANG XY, WU GL, DUAN RY, et al, 2016. Studies on population structure and dynamics ofPinusdabeshanensis[J]. Resourc Environ Yangtze Basin, 25(1): 55-62. [项小燕, 吴甘霖, 段仁燕, 等, 2016. 大别山五针松种群结构及动态研究 [J]. 长江流域资源与环境, 25(1): 55-62.]
XU XH, YU MJ, HU ZH, 2005. The structure and dynamics ofCastanopsiseyreipopulation in Gutian Mountain Natural Reserve in Zhejiang, East China [J]. Acta Ecol Sin,25(3): 645-653. [徐学红, 于明坚, 胡正华, 2005. 浙江古田山自然保护区甜储种群结构与动态 [J]. 生态学报, 25(3): 645-653. ]
YAN GQ, ZHAO JF, HU ZH, 2001. Population structure and dynamics ofLarixchinenesisin Qinling Mouuntain [J]. Chin J Appl Ecol, 12(6): 824-828. [闫桂琴, 赵桂仿, 胡正海, 2001. 秦岭太白红杉种群结构与动态的研究 [J]. 应用生态学报, 12(6): 824-828.]
YI XM, ZHANG Y, JI LZ. 2013, Population structure ofPhellodendronamurensein Changbai Mountains of Northeast China. [J]. Chin J Ecol, 32(9) : 2 257-2 262. [易雪梅,张悦,姬兰柱. 2013,长白山黄檗种群结构 [J]. 生态学杂志, 32(9) : 2 257-2 262.]YI XM, ZHANG Y, WANG YX, et al, 2015. Population structure ofFraxinusmandshuricaon Changbai Mountain [J]. Acta Ecol. Sin, 35(1) : 91-97. [易雪梅, 张 悦, 王远遐, 等, 2015. 长白山水曲柳种群动态 [J]. 生态学报, 35(1) : 91-97.]
Yoshida T, Kamitani T,1999. Growth of a shade-intolerant tree species,Phellodendronamurense, as a component of a mixed-species coppice forest of central Japan [J]. For Ecol Manag, 113(1): 57-65.
YU Q, XIE ZQ, XIONG GM, et al, 2008. Community characteristics and population structure of dominant species ofAbiesfargesiiforests in Shennongjia National Nature Reserve [J]. Acta Ecol Sin, 28(5): 1 931-1 941. [于倩, 谢宗强, 熊高明, 等, 2008. 神农架巴山冷杉(Abiesfargesii)林群落特征及其优势种群结构 [J]. 生态学报, 28(5): 1 931-1 941.]
ZHANG J, SHANGGUAN TL, DUAN YH, et al, 2014. Age structure and dynamics ofQuercuswutaishanicapopulation in Lingkong Mountain of Shanxi Province, China [J]. Chin J Appl Ecol, 25(11): 3 125-3 130. [张婕, 上官铁梁, 段毅豪, 等, 2014. 灵空山辽东栎种群年龄结构与动态 [J]. 应用生态学报, 25(11): 3 125-3 130.]
ZHANG QY, ZHANG YC, LUO P, 2007. Ecological characteristics of aSabiansaltuariapopulation at timberline on the south-facing slope of Baima Snow Mountain, Southwest China [J]. J Plant Ecol, 31(5): 857-864. [张桥英, 张运春, 罗鹏, 2007. 白马雪山阳坡林线方枝柏种群的生态特征 [J]. 植物生态学报, 31(5): 857-864.]
ZHANG QY, LUO P, ZHANG YC, et al, 2008. Ecological characteristics ofAbiesgeorgeipopulation at timberline on the north-facing slope of Baima Snow Mountain, Southwest China [J]. Acta Ecol Sin, 31(5): 857-864. [张桥英, 罗 鹏,张运春,等, 2008. 白马雪山阴坡林线长苞冷杉(Abiesgeorgei)种群结构特征 [J]. 生态学报, 28(1): 129-135.]
ZHANG WH, XU XB, ZHOU JY, et al, 2005. Population dynamics of endangered plant speciesAbieschensiensis[J]. Chin J Appl Ecol, 16(10): 1 799-1 804. [张文辉, 许晓波, 周建云, 等, 2005. 濒危植物秦岭冷杉种群数量动态 [J]. 应用生态学报, 16(10): 1 799-1 804.]
ZHANG YF, LI DW, WANG M, et al, 2015. Population structure and dynamics ofJuniperusrigidain different regions of Loess Plateau [J]. Sci Sil Sin, 51(2): 1-10. [张亚芳, 李登武, 王梅, 等, 2015. 黄土高原不同地区杜松种群结构与动态 [J]. 林业科学, 51(2): 1-10.]
ZHANG ZX, LIU P, CAI MZ, et al, 2008. Population quantitative characteristics and dynamics of rare and endangeredTsugatchekiangensisin Jiulongshan Nature Reserva of China [J]. J Plant Ecol, 32(5): 1 146-1 156. [张志祥,刘鹏,蔡妙珍,等, 2008. 九龙山珍稀濒危植物南方铁杉种群数量动态 [J]. 植物生态学报, 32(5): 1 146-1 156.].
ZOU TC, 2001. Guizhou endemic and rare spermatophyta [M]. Guiyang: Guizhou Science and Technology Publishing House: 29-34. [邹天才, 2001. 贵州特有及稀有种子植物 [M]. 贵阳: 贵州科技出版社: 29-34.]ZOU TC, HUANG CM, ZHOU HY, et al, 2007. Studies on seedling propagation and the cultivation expansion of 10 narrow limited distribution speciesin Guizhou [J]. Seed, 26(7): 52-56. [邹天才, 黄彩梅, 周洪英, 等, 2007. 10种贵州狭限分布植物的种苗扩繁与栽培试验研究 [J]. 种子, 26(7): 52-56.]
Population structure and dynamics of the endemic plantAcerguizhouensein Guizhou Province
LIU Hai-Yan1, YANG Nai-Kun2, ZOU Tian-Cai3*,HUANG Li-Hua3, WANG Jian-Wen3
( 1.GuizhouBotanicalGarden, Guiyang 550004, China; 2.GuiyangZhongxiongForestryEcologicalEngineeringInvestigation&DesignCo.Ltd., Guiyang 550004, China; 3.GuizhouAcademyofSciences, Guiyang 550001, China )
Abstract:Acer guizhouense is a kind of evergreen broad-leaved tree with scientific and economic value, which only distributed at 770-1 000 m in evergreen and deciduous broad-leaved mixed forest with mountain barrier in Xinzhou Town of Huangping County in Guizhou Province. So, it has significant value to explore conservation biology of endemic plants by studying its population structure and dynamic change. We took general survey in suspected distribution area, set up plots in dense area for full investigation, and analyzed the population structure and quantity dynamics pattern. The results were as follows: (1) The natural distribution region of A. guizhouense was very narrow, covered about 0.3 km2 in Leiping with a total of 2 600 trees. The dense area was 0.03 km2, average population density 78 800 trees/km2. The sporadic distribution was 0.27 km2, average population density ≤1 500 trees/km2. The species occasionally distributed in about 0.05 km2 Feiyunya of Dongpo Village. But there was no natural distribution between Leiping and Dongpo. According to the SAFE index and evaluation standard of IUCN, A. guizhouense was EN species. (2) The structure of A. guizhouense population was increasing and the proportion of young tree in population was 63.23%. The size of population density was young trees>middle-aged trees>adult trees. There were 2 peaks in the Ⅰ and Ⅸ age-classes on the mortality rate curve and disappearance rate curve respectively, then there were 2 troughs in Ⅲ-Ⅷ andⅩ-Ⅺ age-classes. So the survival curve of population was more in line with the characteristics of Deevey-Ⅲ model. (3) The main reason of few population individuals of A. guizhouense was the plant death or loss caused by human deforestation and habitat destruction. It is a practical and effective reasonable protection measure to carry out artificial breeding and ex situ cultivation when we strengthen the in situ conservation.
Key words:Acer guizhouense, population structure, time-specific life table, quantity dynamics, endemic distribution
DOI:10.11931/guihaia.gxzw201512006
收稿日期:2015-12-07修回日期: 2016-03-14
基金项目:国家自然科学基金(31360075, 31560097); 国家农业科技成果转化基金(2014GB2F200245)[Supported by the National Natural Science Foundation of China(31360075, 31560097); Agriculture Science Technology Achievement Fund (2014GB2F200245)]。
作者简介:刘海燕(1982-),女, 安徽庐江人,副研究员,从事植物资源学研究,(E-mail)liuhaiyan301@163.com。*通讯作者: 邹天才,研究员,从事植物资源学研究,(E-mail)1211111951@qq.com。
中图分类号:Q948.12
文献标识码:A
文章编号:1000-3142(2016)05-0548-09
刘海燕,杨乃坤,邹天才,等. 贵州特有植物贵州槭的种群结构及动态研究 [J]. 广西植物, 2016, 36(5):548-556
LIU HY, YANG NK, ZOU TC, et al. Population structure and dynamics of the endemic plantAcerguizhouensein Guizhou Province [J]. Guihaia, 2016, 36(5):548-556
