外源ABA处理的玉米叶片酵母双杂交cDNA文库的构建及评价
2016-06-14马芳芳蒋明义
马芳芳+蒋明义
摘要:为深入研究ABA信号转导机制以及筛选该信号途径中相关重要蛋白的相互作用,利用Gateway技术,以外源ABA处理的玉米叶片为材料首先构建了cDNA初级文库,初级文库再通过LR重组的方式最终构建成以pDEST22为目的载体的酵母双杂交cDNA文库。经质量鉴定,该文库的滴度为3.9×106 CFU/mL,文库总容量达到1.17×107 CFU,平均插入cDNA片段长度大于800 bp,重组率大于95%。结果表明所获酵母双杂交文库质量较高,符合文库构建的标准,保证了文库的覆盖度,为进一步开展互作蛋白筛选的后续工作奠定了基础。
关键词:ABA;玉米;酵母双杂交;cDNA文库;滴度;重组率;互作蛋白
中图分类号: S513.01;Q78
文献标志码: A
文章编号:1002-1302(2016)04-0058-04
脱落酸(ABA)是一种重要的植物激素,它在植物生长发育的各个方面,包括种子发育、萌发和休眠、营养生长及对各种生物、非生物胁迫的响应等方面起作用[1-2]。水分胁迫下(包括干旱、盐渍),植物体通过迅速积累ABA以促进气孔关闭以及调节众多基因的表达,从而使植株对逆境环境做出适应反应[3-6]。大量研究表明,ABA信号转导是一个非常复杂的网络,许多重要的中间组分如胞内钙离子、钙调素、磷酸肌醇、cADPR、H+、活性氧、一氧化氮以及蛋白可逆磷酸化等都在ABA信号转导途径中发挥着重要的作用[7]。然而ABA参与的许多信号途径的详细过程特别是具体的分子作用机理仍有待阐明,鉴定参与ABA信号途径的细胞组分,并对其功能进行阐明分析,将有助于我们提高对ABA信号转导途径的认识。
酵母双杂交系统是一种用来研究蛋白质间相互作用的简便、快速而有效的方法[8],该方法由Fields和Song于1989年首先建立,至今被广泛应用于生物研究的各个领域[9-10]。它的一个很重要的功能就是发现新的蛋白质以及发现蛋白质的新功能,即从cDNA文库中筛选出与已知蛋白相互作用的未知蛋白,进而为探索该蛋白的相关特性及功能奠定基础[11-15]。所以,高质量酵母双杂交 cDNA 文库的构建是利用酵母双杂交技术进行大规模互作蛋白筛选的前提和保障[16]。近年来,针对许多重要农作物如水稻[17]、小麦[18]、大麦[19]、玉米[20]等,众多学者构建了以不同组织、器官、细胞类型或分化时期为材料的酵母双杂交cDNA文库,为相关作物的蛋白质组学研究提供了基础。然而国内外关于外源ABA处理的玉米cDNA文库的相关研究还未见报道,本研究以施用外源ABA的玉米为材料,利用Gateway技术构建了ABA处理的玉米叶片酵母双杂交cDNA文库,旨在有效地分离与已知蛋白相互作用的靶蛋白,以期为深入研究ABA信号网络提供一定的理论依据。
1 材料与方法
1.1 供试材料
1.1.1 植物材料 以玉米(Zea mays L.)杂交种农大108为材料,将玉米种子浸泡12 h后,25 ℃催芽,挑取发芽一致的种子播于含有营养液的沙土中培养[昼/夜温度28 ℃/22 ℃,光照度200 μmol/(m2·s),相对湿度75%,昼/夜光周期14 h/10 h]。当幼苗的第2片真叶完全展开时,用锋利的刀片将玉米幼苗从茎基部快速切割下来,放在纯水中2 h以消除伤害胁迫[21],然后用100 μmol/L外源ABA进行处理[温度25 ℃、光强200 μmol/(m2·s)],在不同的处理时间点(0.25、0.5、1、2、4、8、12、24 h),分别剪取第2片叶,迅速置于液氮中冷冻,将最终所得材料混合均匀,-80 ℃ 保存备用。
1.1.2 菌株和载体 大肠杆菌DH10B、文库载体pDONR222(用于构建初级文库)、pDEST22(用于构建酵母双杂交文库)均购自Invitrogen公司。
1.1.3 主要试剂 Trizol Reagent、CloneMiner cDNA Library Construction Kit、FastTrack 2.0 mRNA Isolation Kit、1 kb Plus DNA Ladder、UltraPureTMPhenol-Chloroform-Isoamyl Alcohol (体积比25 ∶24 ∶1) 购自Invitrogen公司;氨苄青霉素、卡那霉素为Sigma公司产品;Taq DNA Polymerase、DNA Marker Ⅲ、琼脂糖凝胶DNA回收试剂盒、质粒小提试剂盒购自天根生化科技有限公司。
1.1.4 引物 M13 Forward,5′-GTAAAACGACGGCCAG-3′; M13 Reverse,5′-CAGGAAACAGCTATGAC-3′; pDEST22 Forward,5′-TATAACGCGTTTGGAATCACT-3′; pDEST22 Reverse,5′-AGCCGACAACCTTGATTGGAGAC-3′。所用引物由上海生工生物技术有限公司合成。
1.2 玉米叶片酵母双杂交cDNA文库的构建
1.2.1 玉米叶片总RNA的提取和mRNA的纯化 取玉米叶片,在预冷的研钵中将其用液氮研磨成细粉,用Trizol提取其总RNA,具体过程参照说明书进行。得到的总RNA经过质量及纯度检测之后,用于下一步的mRNA纯化操作,具体的纯化过程参照FastTrack2.0 Kit说明书进行。得到的mRNA经质量及纯度检测之后,用于下一步的文库构建操作。
1.2.2 cDNA初级文库的构建 参照CloneMiner cDNA Library Construction Kit说明书,按以下步骤进行:首先将上步纯化得到的mRNA进行cDNA第1链的合成;然后进行cDNA第2链的合成及纯化;cDNA与三框重组接头attB1 Adapter连接后进行cDNA的分级分离及收集;利用Gateway技术将收集得到的cDNA与入门文库载体质粒pDONR222混合进行BP重组反应;重组反应产物经Proteinase K处理以及纯化之后,利用电转化法转入大肠杆菌DH10B;加入1 mL SOC培养基,37 ℃培养1 h,即为cDNA初级文库菌液;加甘油至终浓度20%,-80 ℃保存备用。
1.2.3 酵母双杂交cDNA文库的构建 参照CloneMiner cDNA Library Construction Kit说明书,将上述得到的cDNA初级文库菌液接种于含有卡那霉素抗性的肉汤培养基中,30 ℃过夜培养;第2天中抽质粒并检测吸光度;将中抽得到的质粒稀释到终浓度300 ng/μL,取出其中1 μL,利用Gateway技术将其与酵母双杂交文库载体质粒pDEST22(含GAL4 Transcriptional-Activation Domain)混合进行LR重组反应;重组反应产物经Proteinase K处理以及纯化之后,利用电转化法转入大肠杆菌DH10B;加入3 mL SOC培养基,37 ℃培养1 h,即为酵母双杂交cDNA文库菌液;加甘油至终浓度20%,-80 ℃保存备用。
1.2.4 文库质量的鉴定
1.2.4.1 库容量的鉴定 分别从“1.2.2”节和“1.2.3”节得到的转化后文库细菌原液中取出10 μL,稀释1 000倍后,从中吸出50 μL涂布于含有相应抗性的LB平板上(初级文库鉴定平板使用卡那霉素抗性、酵母双杂交文库鉴定平板使用氨苄青霉素抗性),37 ℃培养12~16 h后进行计数,根据公式:文库滴度(CFU/mL)=平板克隆数×稀释倍数/涂板体积;文库总容量(CFU)=文库滴度×文库总体积,计算出文库的滴度以及总容量。
1.2.4.2 重组率和插入片段长度鉴定 从平板上随机挑取24个单克隆进行菌落PCR,初级文库菌落PCR使用的引物Primer 1和引物Primer 2分别是M13 Forward和M13 Reverse,酵母双杂交文库菌落PCR使用的引物Primer 1和引物Primer 2分别是pDEST22 Forward和pDEST22 Reverse,PCR反应条件:94 ℃预变性5 min;94 ℃变性30 s,56 ℃复性30 s,72 ℃延伸3 min,共30个循环;72 ℃延伸10 min;4 ℃保温。PCR产物用1% Agarose 凝胶电泳检测,鉴定插入片段大小和阳性克隆重组率。
2 结果与分析
2.1 玉米叶片总RNA和纯化的mRNA质量分析
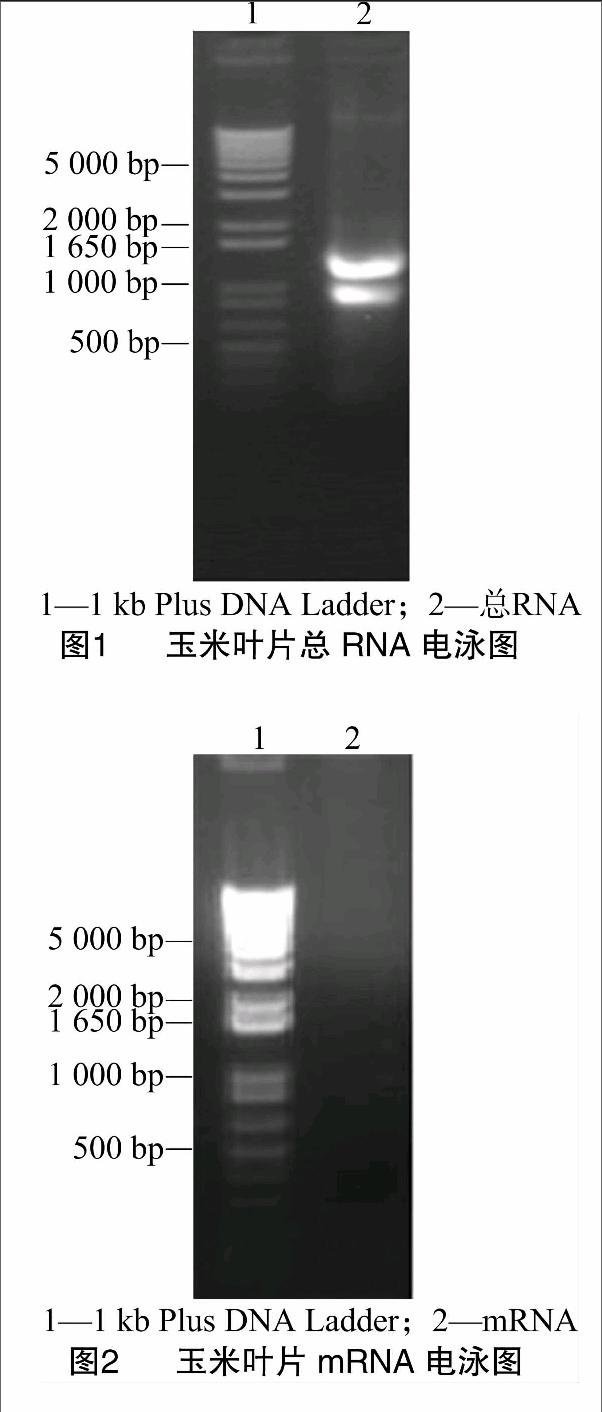
利用Trizol Reagent提取了外源ABA处理的玉米叶片总RNA,用1%琼脂糖凝胶电泳检测,可见28S和18S条带明亮清晰,亮度比约2 ∶1,表明该总RNA完整性好且基本上未发生降解(图1)。经总RNA紫外分析测定,结果显示D260 nm/D280 nm为1.95,表明总RNA纯度好,符合实验要求,可用于后续实验操作。总RNA样品经过分离纯化后得到了mRNA,电泳图结果可见其在较大范围内呈涂布状分布(图2),与预期结果相符,说明纯度较高,完整性较好。
2.2 cDNA初级文库的质量鉴定
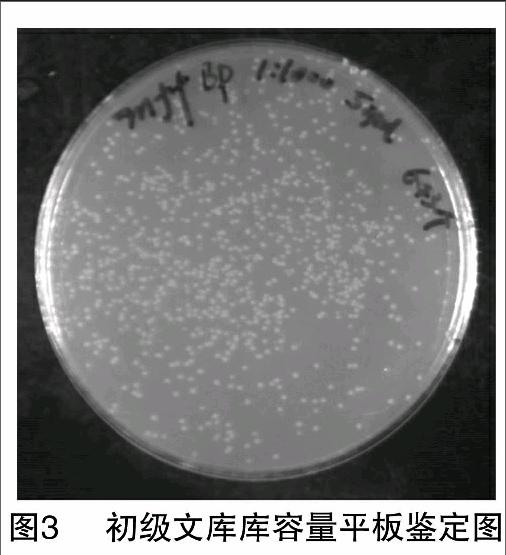
2.2.1 库容量测定 从cDNA初级文库原始菌液中取出10 μL,稀释1 000倍后,吸出50 μL涂布于含卡那霉素抗性的LB平板上,37 ℃培养12~16 h后计菌落数,结果为643个(图3),菌液量为1 mL。根据公式:文库滴度(CFU/mL)=平板克隆数×稀释倍数/涂板体积;文库总容量(CFU)=文库滴度×菌液总体积,计算得出:cDNA初级文库的滴度=643×1 000/0.05=1.28×107 CFU/mL;cDNA初级文库的总容量=1.28×107 CFU/mL×1 mL=1.28×107 CFU。
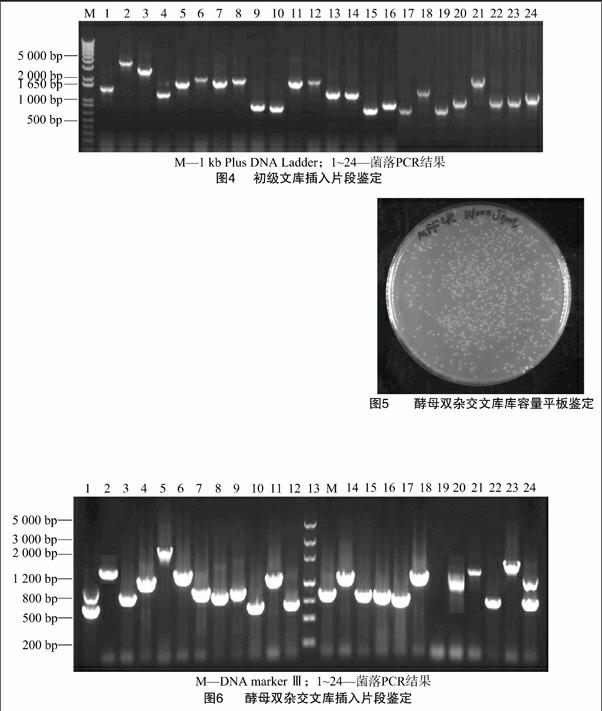
2.2.2 重组率和插入片段大小测定 从平板上随机挑取24个单克隆进行菌落PCR,PCR产物用1% Agarose 凝胶电泳检测,根据电泳图,可以看出所有克隆的PCR结果都呈阳性,重组率达100%,插入cDNA片段长度大小不等,主要分布在500~3 000 bp范围内,多态性较好,且平均插入片段长度大于1 000 bp(图4)。
2.3 酵母双杂交文库cDNA文库的质量鉴定
2.3.1 库容量测定 从酵母双杂交cDNA文库原始菌液中取出10 μL,稀释1 000倍后,吸出50 μL涂布于含氨苄青霉素抗性的LB平板上,37 ℃培养12~16 h后计菌落数,结果为195个(图5),菌液量为3 mL。根据公式:文库滴度(CFU/mL)=平板克隆数×稀释倍数/涂板体积;文库总容量(CFU)=文库滴度×菌液总体积,计算得出:酵母双杂交文库的滴度=195×1 000/0.05=3.9×106 CFU/mL;酵母双杂交文库的总容量=3.9×106 CFU/mL×3 mL=1.17×107 CFU。
2.3.2 重组率和插入片段大小测定 从平板上随机挑取24个单克隆进行菌落PCR,PCR产物用1% Agarose 凝胶电泳检测,从电泳图中可以看出24个克隆中有23个为阳性结果,重组率大于95%,cDNA插入片段长度大小不等,主要分布在500~3 000 bp范围内,多态性较好,且平均插入片段长度大于800 bp(图6)。
3 讨论
一个高质量酵母双杂交 cDNA 文库的获得,能够为我们进行大规模互作蛋白的筛选提供前提和保障,对于我们研究相关生物的功能蛋白质组学具有重要意义。目前很多生物种类的cDNA文库已经获得,并且在此基础上获得了大量的信息[17-20],然而关于外源ABA处理的玉米cDNA文库的相关研究还未见报道。已有的研究发现,一些蛋白激酶如MAPK[22]、CCaMK[23-25]、CDPK[26]等在ABA诱导的抗氧化防护过程中发挥着重要的作用,我们拟利用酵母双杂交技术筛选与这些蛋白相互作用的靶蛋白,进而探究该蛋白在ABA信号途径中的具体作用机制,因此,外源ABA处理的玉米叶片酵母双杂交cDNA文库的建立就成为了不可或缺的重要基础工具。
RNA质量的好坏直接影响文库质量的高低,本研究中提取的玉米叶片总RNA纯度高、完整性好且基本上未发生降解,经分离纯化后得到的mRNA在较大范围内呈涂布状分布,具有较高的纯度和完整性,可用于后续的文库构建。本研究在构建酵母双杂交cDNA文库的过程中使用了Gateway技术,一种基于已研究的非常清楚的λ噬菌体位点特异性重组系统,该系统能够高效而快速地将DNA定向重组进入不同的载体系统中,进而应用于蛋白的表达与功能分析[27-29]。由于在载体构建时无需经历限制性内切酶的酶切和酶连等过程,并且通过引入3种不同的接头使得插入片段能够包含3种不同的读码方式,这样既降低了嵌合克隆出现的概率、保证了插入片段的序列完整性,同时又克服了由于插入片段中非编码区带有终止密码子而造成的蛋白提前终止翻译,有效地提高了文库的完整性和覆盖性[30]。
通常一个高质量的cDNA文库,其库容量应至少大于 1×106 CFU,这样才能保证cDNA种类的完整性以及文库的代表性。本试验对构建的玉米叶片酵母双杂交cDNA文库的分析结果表明,库容量为1.17×107 CFU,保证了文库的覆盖度。而平均插入cDNA片段长度则体现了文库遗传信息的完整性,如果大部分克隆插入了全长cDNA序列,则说明文库的完整性好。本试验构建的文库重组率大于95%,平均插入cDNA 片段长度大于800 bp,说明该文库达到了高质量文库的标准,可以用来进行后续的大规模互作蛋白的筛选试验。
参考文献:
[1]Zhu J K. Salt and drought stress signal transduction in plants [J]. Annu Rev Plant Biol,2002,53:247-273.
[2]Xu Z Y,Kim D H,Hwang I. ABA homeostasis and signaling involving multiple subcellular compartments and multiple receptors [J]. Plant Cell Rep,2013,32:807-813.
[3]Cutler S R,Rodriguez P L,Finkelstein R R,et al. Abscisic acid:emergence of a core signaling network [J]. Annu Rev Plant Biol,2010,61:651-679.
[4]Fujita Y,Fujita M,Shinozaki K,et al. ABA-mediated transcriptional regulation in response to osmotic stress in plants [J]. J Plant Res,2011,124:509-525.
[5]Hubbard K E,Nishimura N,Hitomi K,et al. Early abscisic acid signal transduction mechanisms:newly discovered components and newly emerging questions [J]. Genes Dev,2010,24:1695-1708.
[6]Umezawa T,Nakashima K,Miyakawa T,et al. Molecular basis of the core regulatory network in ABA responses:sensing,signaling and transport [J]. Plant Cell Physiol,2010,51:1821-1839.
[7]Wasilewska A,Vlad F,Sirichandra C,et al. An update on abscisic acid signaling in plants and more [J]. Mol Plant,2008,1:198-217.
[8]张雨良,张树珍,王健华,等. 感染高粱花叶病毒甘蔗叶片cDNA文库构建及评价 [J]. 热带作物学报,2012,33(6):1096-1100.
[9]Fields S,Song O. A novel genetic system to detect protein-protein interactions [J]. Nature,1989,340:245-246.
[10]Song O,Dolan J W,Yuan Y L,et al. Pheromone-dependent phosphorylation of the yeast STE12 protein correlates with transcriptional activation [J]. Genes Dev,1991,5:741-750.
[11]Walhout A J,Boulton S J,Vidal M. Yeast two-hybrid systems and protein interaction mapping projects for yeast and worm [J]. Yeast,2000,17:88-94.
[12]Ito T,Ota K,Kubota H,et al. Roles for the two-hybrid system in exploration of the yeast protein interactome [J]. Mol Cell Proteomics,2002,1:561-566.
[13]Auerbach D,Thaminy S,Hottiger M O,et al. The post-genomic era of interactive proteomics:facts and perspectives [J]. Proteomics,2002,2:611-623.
[14]Chen Y,Xu D. Computational analyses of high-throughput protein-protein interaction data [J]. Curr Protein Pept Sci,2003,4:159-181.
[15]Cho S,Park S G,Lee D H,et al. Protein-protein interaction networks:from interactions to networks [J]. J Biochem Mol Biol,2004,37:45-52.
