铵硝配比对巴西香蕉生长和氮素营养的影响
2016-05-30王岚黄承和陈玉子朱治强常春荣
王岚 黄承和 陈玉子 朱治强 常春荣

摘 要 采用水培法,在等氮条件下设置5种不同铵硝配比处理,探讨不同铵硝配比对巴西香蕉幼苗生长及其氮素营养特性的影响。结果表明,等氮条件下,适当地提高铵态氮比例可以提高香蕉的生物量;铵硝比为10 ∶ 90最适合香蕉的生长;增铵促进根系的生长,根系中铵、硝含量分别与培养液中铵态氮和硝态氮的含量有关。香蕉幼苗中氮的含量、累积量与铵硝配比有关,增加铵的比例地上部分氮含量和累积量高于全硝处理,促进氮的吸收和利用;铵硝配比影响香蕉叶绿素含量、光合速率以及硝酸还原酶活性,光合速率与叶绿素含量、硝酸盐含量与硝酸还原酶活性没有明显的正相关关系。在香蕉生长早期适当增加铵态氮供应可以促进香蕉根系的生长,有利于香蕉的早发、稳长。
关键词 巴西香蕉;铵硝比;生长;氮素营养
中图分类号 S668.1 文献标识码 A
Abstract Study on growth and nitrogen nutrition physiological characteristics in Brazil Banana seedling by hydroponic culture experiment with five ratio of ammonium to nitrate at equal nitrogen concentration condition. These results were showed that: At this condition, banana biomass increased with suited ammonium concentration and the ratio of NH4+/NO3- 10 ∶ 90 was the most suitable to its growth. Increasing ammonium ratio promoted banana root development, and concentration of ammonium and nitrate in its body related to the concentration of ammonium and nitrate in culture solution respectively. In banana nitrogen concentration and content related to the ratio of NH4+/NO3-; amd increasing ammonium ratio shoot nitrogen concentration and content with higher ratio of utilization and absorption was higher than that of 100% nitrate treatment. Although there were no distinct linearity relation between chlorophyll concentration and photosynthesis rate, or nitrate concentration and nitrate reductive enzyme activity, ratio of NH4+/NO3- influenced these physiological characteristics. Therefore, increasing supply of suited ammonium in banana seedling stage will promote root growth, thus this make it growth fleetly and steadily.
Key words Brazil banana;Ratio of ammonium to nitrate;Growth;Nitrogen nutrition
doi 10.3969/j.issn.1000-2561.2016.01.005
铵硝配比影响植物的生长发育,不同的作物对铵硝配比的响应特征并不相同[1-9]。香蕉是海南重要的水果作物,其对氮的吸收量仅次于钾[10]。目前就香蕉氮肥用量、施用技术、与其他肥料配比、与水分的耦合等方面开展了广泛的研究[10-11],但氮素形态及合理的配比对香蕉生长发育的影响研究尚未见报道。因此,本试验以巴西香蕉幼苗为材料,研究不同硝铵比对其生长的影响,以期探明适宜巴西香蕉生长的最佳氮素形态配比,为进一步合理施用氮肥提供参考依据。
1 材料与方法
1.1 材料
香蕉苗为海南大学儋州校区种苗组培中心的组培杯苗(四叶一心), 巴西种(Musa AAA Cavendish subgroup cv.Brazil)。
1.2 方法
1.2.1 试验设计 2011年3~4月在海南大學儋州校区农学院教学实习基地塑料大棚内进行。将香蕉根部基质洗去后移栽到自制泡沫板的孔中(Φ2 cm),并用海绵固定,置有供氧装置的20 L不透光周转箱(长×宽×高=48 cm×35 cm×16 cm),根系浸入水中进行培养。每个周转箱培养香蕉苗8株,重复3次。
霍格兰营养液基础上,等氮条件下设5个铵硝比例处理,分别为0 ∶ 100、10 ∶ 90、25 ∶ 75、50 ∶ 50和75 ∶ 25。NO3--N用NaNO3,NH4+-N用NH4CI,总氮浓度均为15 mmol/L,重复3次。并保持其它营养元素用量和比例与改良的霍格兰营养液配方一致。在营养液中加入7 μmmol/L硝化抑制剂二氰胺(DCD),以防止营养液中NH4+-N转化为NO3--N。每天调节pH到5.8,通气12 h/d(6:00~18:00), 供氧机功率为 120 L/min。每3 d更换1次营养液。培养21 d后,采样测定。
1.2.2 测定项目与方法 生物量:重量法。收获后,将幼苗地上和根系分开,分别称鲜重;然后烘干后称干重。叶绿素 ∶ 乙醇和丙酮(体积1 ∶ 1)浸提,分光光度法测定。铵态氮和硝态氮:新鲜的植物样品用沸水浴浸提后,用纳氏比色法测定铵态氮含量和双紫外分光光度法测定硝态氮含量[12]。全氮:样品用浓H2SO4-H2O2消煮,采用凯氏法测定植物全氮。硝酸还原酶:采用活体法[13]进行测定。叶片净光合速率:选择晴天,LI-6400型便携式光合测定仪于上午9:30~11:30测定叶片净光合速率(内置红蓝光源,1 000 μmol/m2·s)。
1.3 数据处理
数据采用Microsoft excel 2003软件进行处理,用SAS 9.0软件进行显著性分析。
2 结果与分析
2.1 巴西香蕉生物量与根冠比
由表1可知,铵硝配比对巴西香蕉生物量有显著影响;地上部分干重和总干重在铵硝比为10 ∶ 90最高,其次为全硝营养和铵硝比为25 ∶ 75,50 ∶ 50和75 ∶ 25处理没有显著差异;铵硝比50 ∶ 50处理最低;香蕉幼苗根系干重以铵硝比75 ∶ 25处理最高,全硝营养处理最低,根冠比均表现出与根系干重相一致的规律。说明铵态氮供应比例增加抑制地上部分的生长,促进根系的生长。
2.2 氮含量及单株氮累积量
由表2可知,铵硝配比显著影响巴西香蕉氮含量和氮累积量;铵硝比50 ∶ 50时香蕉植株地上部分氮含量达到最高,全硝营养时最低,前者是后者的1.2倍;根系氮含量以全硝营养时最高,铵硝比10 ∶ 90时最低。说明增铵有利于提高地上部分氮含量,降低根系氮含量。
地上部分和整个植株的单株氮累积量以铵硝比10 ∶ 90处理最高,其次为铵硝比25 ∶ 75,均显著高于全硝营养的氮累积量;根系单株氮累积量则以处理铵硝比75 ∶ 25为最高,较全硝营养处理提高了28.9%。
比较氮累积量与生物量发现,氮的累积量受到生物量的影响,其差异规律与生物量一致。
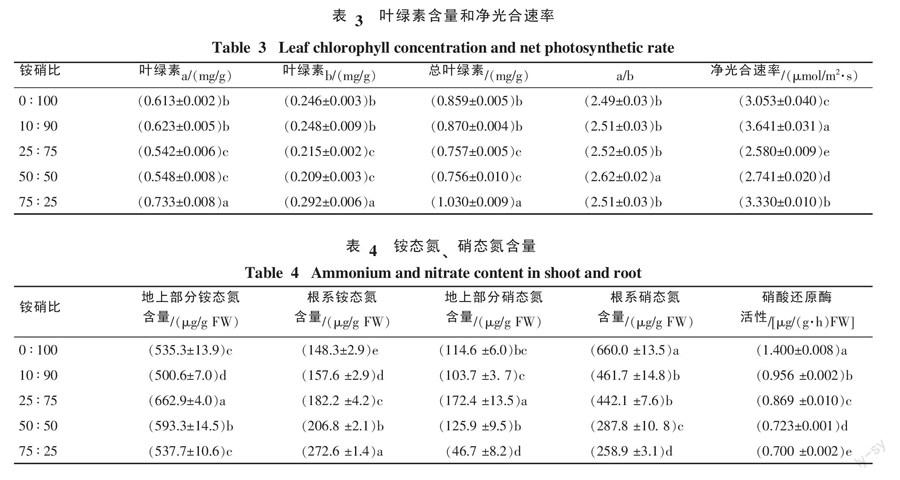
2.3 叶片叶绿素含量和净光合速率
由表4可知,叶绿素a、叶绿素b和总叶绿素含量表现出相同的规律,铵硝比75 ∶ 25处理最高, 50 ∶ 50和25 ∶ 75处理最低,0 ∶ 100处理与10:90处理居中,并且叶绿素a、叶绿素b和总叶绿素含量分别在50 ∶ 50与25 ∶ 75处理、0 ∶ 100与10 ∶ 90处理之间没有显著差异;叶绿素a与叶绿素b比值表现出铵硝比50 ∶ 50处理显著高于其它处理,而其它处理间没有显著差异。
分析净光合速率与a/b比值关系发现,净光合速率与a/b比值之间的关系不明显;分析叶a/b比值与生物量的关系可知,a/b在一定程度上反映植物光能利用率的高低,但植株生物量的增加还与其它因素有关;分析净光合速率与生物量的关系发现,铵硝比10 ∶ 90處理生物量高的原因可能与光合速率高有关,但生物量在不同处理之间的关系与光合速率的规律并不完全一致。
2.4 硝态氮和铵态氮含量
由表4可知,地上部分铵态氮含量以铵硝比25 ∶ 75处理最高,其次为50 ∶ 50处理,10 ∶ 90处理最低,0 ∶ 100和75 ∶ 25处理之间没有显著差异。根系铵态氮含量随营养液中铵的比例增大而提高。分析铵态氮含量与氮含量的关系发现,铵态氮含量与氮含量在各处理间的规律并不完全一致。
地上部分硝态氮含量与地上部分铵态氮含量有一致的分布规律,根系硝态氮含量与根系铵态氮的分布规律相反。与硝酸还原酶活性对比发现,叶片硝酸还原酶的活性并不与植株体内的硝态氮含量正相关。对比根系和地上部分铵态氮含量发现,地上部分显著高于根系,硝态氮含量分布规律则相反。铵硝配比影响香蕉叶片硝酸还原酶活性。叶片硝酸还原酶活性随营养液硝态氮比例的增加而提高,以全硝营养最大,其叶片硝酸还原酶活性是75 ∶ 25处理的2.0倍。
3 讨论与结论
在氮总浓度不变的条件下,适当地提高铵态氮比例可以增加作物的产量[3-4]。本研究结果使得这一结论在香蕉上得到验证。在不同的作物上最佳的铵硝比例并不相同[4-9], 本研究结果以铵硝比为10 ∶ 90处理适合香蕉的生长。本研究结果发现,全铵处理的香蕉苗严重生长不良,根系甚至有中毒症状,表明香蕉不适合全铵营养。铵对根系的影响在不同的作物上并不完全一致[5-8],但本研究结果证明了增铵促进香蕉根系的生长。结合香蕉生长的规律,在香蕉生长早期适当的增加铵态氮肥可以促进香蕉根系的生长,有利于香蕉的早发、稳长。
关于铵硝配比对植株中氮含量的影响,不同的作物表现不一致[6,7,9]。本研究结果发现,香蕉在等氮量的条件下,提高铵比例有利于提高地上部分氮含量,促进氮的合成代谢,降低根系氮含量,与赵丽莉等[1-2]的研究结果一致。铵含量主要代表游离氨基酸的含量[13],根系中铵、硝含量分别与培养液中铵态氮和硝态氮的含量有关[15-16]。本研究结果表明,不同铵硝配比不但影响根系和地上部分氮的含量,同时影响铵和硝的代谢,铵硝比为10 ∶ 90有利于香蕉对氮的吸收和利用,根系中铵态氮和硝态氮的含量分别与营养液中铵态氮和硝态氮含量一致,这与陈巍等[4,8,10,15-16]研究结果一致。在本试验条件下,叶片硝酸还原酶的活性并不与植株体内的硝态氮含量正相关,可能是硝酸盐含量与硝酸还原酶的活性、吸收速度以及基因的表达水平有关[17-18]。
作物叶片的净光合速率受到不同形态的氮素营养供应影响[16,19]。曹翠玲等[20]认为氮素形态主要影响了叶绿素b的含量,从而影响植株叶绿素总含量。王波等[8]认为,与全硝营养相比,增铵营养能够有效地提高生菜叶片中叶绿素含量,进而提高叶片净光合速率。本研究结果表明,铵硝配比影响叶绿素a和叶绿素b含量,叶绿素含量与净光合速率存在正相关关系,可能是高比例的铵态氮营养使作物叶片伸展受阻,叶片变小,物质稀释效应小,导致单位面积上物质积累相应增加,表现出与菠菜的铵硝反应结果相一致的机理[21]。本研究结果发现,光合速率比较低,可能与本试验主要在荫蔽的大棚中培养有关[22-24]。
因此,铵硝配比影响香蕉幼苗生长,以铵硝配比10 ∶ 90最适合香蕉的生长,等氮条件下增铵可以提高根系的生长,不同的铵硝配比对地上部分影响不同。苗期增施少量铵可以促进香蕉根系生长,促进香蕉成活和生长。香蕉幼苗中氮的含量、累积量以及铵、硝的含量与铵硝配比有关,在全硝的基础上增铵可以提高氮在地上部分的含量和累积量,促进氮的吸收和利用。铵硝配比影响香蕉叶绿素含量、光合速率以及硝酸还原酶活性,本研究中光合速率与叶绿素含量、硝酸盐含量与硝酸还原酶活性没有明显的正相关关系。
参考文献
[1] 李海亮,郑秀芳. 硝态氮和铵态氮对观赏百合生长发育的影响[J]. 南方园艺, 2011, 22(1): 14-17.
[2] 赵丽莉, 邓光存,吴晓玲. 不同铵态氮和硝态氮配比对黄芩幼苗生长及生理特性的影响[J]. 北方园艺, 2010(5): 191-193.
[3] 黄东风, 李卫华, 邱孝煊. 不同硝、铵态氮水平配施对小白菜生长及硝酸盐累积的影响[J]. 土壤通报, 2010, 41(2): 394-398.
[4] 陈 巍, 罗金葵, 姜慧梅, 等.不同形态氮素比例对不同小白菜品种生物量和硝酸盐含量的影响[J]. 土壤学报, 2004, 41(3): 420-425.
[5] 邓兰生, 齐 庆, 振龚林,等. 滴施不同铵硝比例氮肥对马铃薯生长的影响[J]. 中国土壤与肥料, 2011(4): 13-16.
[6] 段英华, 张亚丽, 王松伟, 等. 铵硝比(NH4+/NO3-)对不同氮素利用效率水稻的生理效应[J]. 南京农业大学学报, 2007, 30(3): 73-77.
[7] Duan Y H, Yin X M, Zhang Y L, et al. Mechanisms of enhanced rice growth and Nitrogen uptake by nitrate[J]. Pedosphere, 2007, 17(6): 697-705.
[8] 王 波, 沈其荣, 赖 涛,等. 不同铵硝比对营养液对生菜生长发育影响的研究[J]. 土壤学报, 2007, 44(3): 561-565.
[9] 许如意, 别之龙,黄丹枫. 不同氮素形态配比对网纹甜瓜干物质分配和氮代谢的影响[J]. 农业工程学报, 2005, 21(2): 147-150.
[10] 杨 阳, 郑秋玲, 裴成国,等. 不同硝铵比对霞多丽葡萄幼苗生长和氮素营养的影响[J]. 植物营养与肥料学报, 2010, 16(2): 370-375.
[11] 于俊红, 彭智平, 黄继川,等. 水氮耦合对香蕉养分吸收和产量的影响[J]. 灌溉排水学报, 2010, 29(6): 136-138.
[12] Kuang Shizi1, Tian Shiyao, Li Chunyu, et al. Effect models of Nitrogen, phosphorus and potassium fertilizer formulation in banana[J]. Agricultura Science & Technology, 2010, 11(9-10): 130-135.
[13 呂伟仙, 葛 滢, 吴建之, 等. 植物中硝态氮、 氨态氮、总氮测定方法的比较研究[J]. 光谱学与光谱分析, 2004, 24(2):204-206.
[14] 李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社,2001:123-124.
[15] 张英鹏, 林咸永, 章永松, 等. 不同氮素形态对菠菜生长及体内抗氧化酶活性的影响[J]. 浙江大学学报(农业与生命科学版), 2006, 32(2): 139-144.
[16] 戴廷波, 曹卫星, 孙传范,等. 增铵营养对小麦光合作用及硝酸还原酶和谷氨酰胺合成酶的影响[J]. 应用生态学报, 2003, 14(9): 1 529-1 532.
[17] Cao Y, Fan X R, Sun S B, et al. Effect of nitrate on activities and transcript levels of nitrate reductase and glutamine synthetase in rice[J]. Pedosphere, 2008, 18(5):664-673.
[18] Zhao X Q, Zhao S P, Shi W M. Enhancement of NH4+ uptake by NO3- in relation to expression of nitrate-induced genes in rice(Oryza sativa)roots[J]. Pedosphere, 2008, 18(1):86-91.
[19] 肖 凯, 张树华, 邹定辉,等. 不同形态氮素营养对小麦光合特性的影响[J]. 作物学报, 2000, 26(1): 53-58.
[20] 曹翠玲, 李生秀. 氮素形态对小麦中后期的生理效应[J]. 作物学报, 2003, 29(2): 258-262.
[21] Steingrover E, Woldeudorp J, Sijtsma L. Nitrate accumulation and its relation to leaf elongation in spinach leaves[J]. Journal of Experimental Botany, 1986, 37(181): 1 093-1 102.
[22] 张智顺, 张庆费, 夏 檑,等. 遮阴对几种绿化植物光合特性和生长的影响[J]. 东北林业大学学报, 2010, 38(3): 47-50.
[23] 张斌斌, 姜卫兵, 翁忙玲,等. 遮阴对园艺园林树种光合特性的影响[J]. 经济林研究, 2009, 27(3): 115-119.
[24] 张往祥, 曹福亮, 吴家胜,等. 遮阴对银杏光合性能及其叶片产量的影响[J]. 南京林业大学报, 2000, 24(4): 11-15.
责任编辑:古小玲
