牡竹属3种竹种光合特性及其影响因子分析
2016-05-30刘蔚漪辉朝茂陆燕元陶红梅黄丕高
刘蔚漪 辉朝茂 陆燕元 陶红梅 黄丕高
摘 要 运用Li-6400便携式光合测定仪对牡竹属3种竹种(龙竹、野龙竹、云南甜龙竹)的光合特性进行测定,研究牡竹属竹种光合速率的日变化规律,及其与影响因子的相关性,为牡竹属优良选育提供理论基础。结果表明:不同竹种光合速率的日变化虽有不同,但变化规律具有相似性,即光合速率日变化都呈双峰曲线,具明显的午休现象。叶片厚度依次为野龙竹(0.15 mm)>龙竹 (0.14 mm)>云南甜龙竹(0.13 mm),而比叶重云南甜龙竹最大(0.08 mg/mm2);龙竹和野龙竹相对较小(0.07 mg/mm2),说明云南甜龙竹较龙竹和野龙竹有较多的有机物积累。光合速率受环境因子和生理因子共同影响。其中,叶片生理因子的直接影响更大,尤以胞间气孔导度、CO2浓度、蒸腾速率最为显著,环境因子中,以光照强度的影响最大。
关键词 牡竹属;光合特性;环境因子;生理因子
中图分类号 Q945.1 文献标识码 A
云南甜龙竹(Dendrocalamus brandisii),龙竹(D. giganteus)、野龙竹(D. semiscandens)属于牡竹属的大型丛生竹,其中云南甜龙竹是优良的笋材两用竹,龙竹、野龙竹也是云南山区重要的森林资源之一。光合作用是植物物质代谢和能量转换的基础,地球上一切生命的能量和物质都来源于植物的光合作用[1-2]。植物的光合速率相关指标能够反映林木的生产能力,对林木栽培及管理具有重要的理论指导意义,也是选育高效优良竹种的重要标准。近年来,国内外研究人员对植物光合特性的相关研究做了很多工作,中国学者对江浙一带栽培较多的笋用竹种[3]、园林上用的较多的地被竹进行了光合特性研究[4],不同控制处理,如酸雨胁迫[5]、施肥梯度[6],CO2浓度,光照强度[7]等不同竹种的反应状况,以及对不同竹龄雷竹的光合特性进行了研究[8],但对大型丛生竹的光合特性研究较少。本研究通过对3种牡竹属竹种(龙竹、野龙竹、云南甜龙竹)光合特性及影响因子进行试验分析,揭示了其光合特性及主要影响因子,为研究云南大型丛生竹种的品种选育,引种改良提供科学依据和理论指导。
1 材料与方法
1.1 材料
试验材料为牡竹属3种竹种(龙竹、野龙竹、云南甜龙竹)。试验在西南林业大学珍稀特有竹种园进行。该竹园位于云南省昆明市东北方向,东经102°45′、北纬25°4′,海拔1 920 m。昆明市年平均温度为14.7 ℃,最高温度为31.5 ℃,大于10 ℃的活动积温为4 479.7 ℃,年平均降雨量1 011.8 mm,主要集中在5~10月,占全年降水量的88.45%,年平均蒸发量为1 867.7 mm,干湿季明显,气候主要受西南季风大陆气团的影响,春、冬季常为旱季。仪器:美国Li-COR公司生产的Li-6400XT便携式光合作用测定系统。
1.2 方法
1.2.1 生理指标和环境因子测定 2014年12月15~18日(天气晴朗,无持续风向,选择在冬季而不是夏季做光合试验,是因为夏季光照过于强力,容易凸显其光照因子的影响作用)。选取生长良好、无病虫害的2年生竹子为样本,取向阳面、面积相当的健康成熟叶片2~3片,每一竹种选3株。利用自然光照,从8:30~17:30时每隔1 h测定一次,3次重复取平均值。仪器自动记录净光合速率(Pn)、大气温度(Tair)、叶片温度(Tleaf)、叶室温度(Tblock)、胞间CO2浓度(Ci)、气孔导度(Cond),蒸腾速率(Trmmol)、相对湿度(RH-R)和叶室相对湿度(RH-S)、参考气体CO2浓度(CO2R)、叶室气体CO2浓度(CO2S),叶室光合有效辐射(PARi)、外界光合有效辐射(PARo)、仪器工作状态等20余项。
1.2.2 叶绿素含量的测定 叶绿素提取采用浸提法。取一健康植株,选取外围枝的叶片,要求颜色鲜绿,无虫眼,清洗干净后擦干,将叶片剪碎至3 mm2左右,随即取样称取0.25 g(精确到0.01 g)放入小锥形瓶中。用丙酮︰乙醇︰蒸馏水体积比=4.5︰4.5︰1.0的浸提液定容至20 mL,在暗处静置36 h。叶绿素含量采用紫外分光光度法测量。以浸提液为空白对照,用紫外分光光度计分别测得叶绿素提取液在波长665 μm和649 μm处的吸光值,利用公式计算叶绿素的含量。
1.2.3 叶片厚度及比叶重的测定 同上,每个竹种取健康叶片各50片,每10片为1组,用游标卡尺测量叶片厚度(精确到0.02 mm),5次重复。用打孔器从50片叶片中打取直径0.6 cm的圆片100个(避开叶脉)。在80 ℃恒温下持续烘8 h,测得其干重,计算比叶重(比叶重=干重/叶面积),5次重复。
1.3 数据分析
数据采用Microsoft Excel 2007和SPSS软件进行统计分析和图表处理。
2 结果与分析
2.1 3种牡竹属竹种光合速率日变化规律
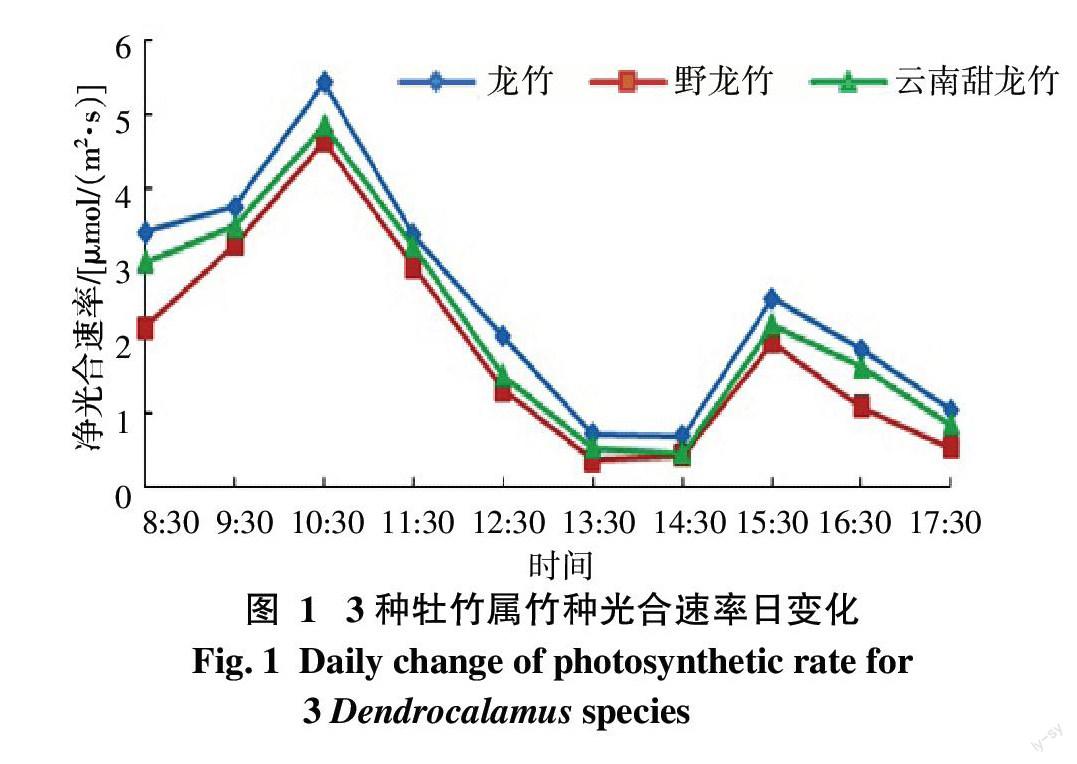
由图1可知,龙竹、野龙竹、云南甜龙竹净光合速率日变化规律基本相同;3个竹种的光合速率日变化趋势均表现为双峰曲线,上午随光照的增强而提高,在10:30达到第一个高峰值,中午前后受到高温和强光的抑制,光合速率降低,而产生“午休”现象。下午14:30~15:30由于光强下降,强度较为合适,净光合速率有所提高,在15:30时达到第二个高峰值。下午16:30之后太阳渐渐西斜,光照强度开始大幅度减弱,净光合速率随之降低。下午的高峰值低于上午的高峰值,光合速率平均相当于第1峰值的44.61%,比较第一高峰值,平均下降幅度达89.50%。
叶片对光能的利用率与日最大净光合速率直接相关,从图1可以看出,龙竹的日最大净光合速率最高[5.42 μmol/(m2·s)],野龙竹的日最大净光合速率最低[4.64 μmol/(m2·s)]。日均净光合速率以龙竹的最高,为2.487 μmol/(m2·s),野龙竹的最低,为1.867 μmol/(m2·s)。龙竹的杆型高大,枝叶茂盛,其生产力要大于云南甜龙竹和野龙竹,这也正好说明净光合速率是影响光合生产力的重要因素。
2.2 3叶片特性对光合速率的影响
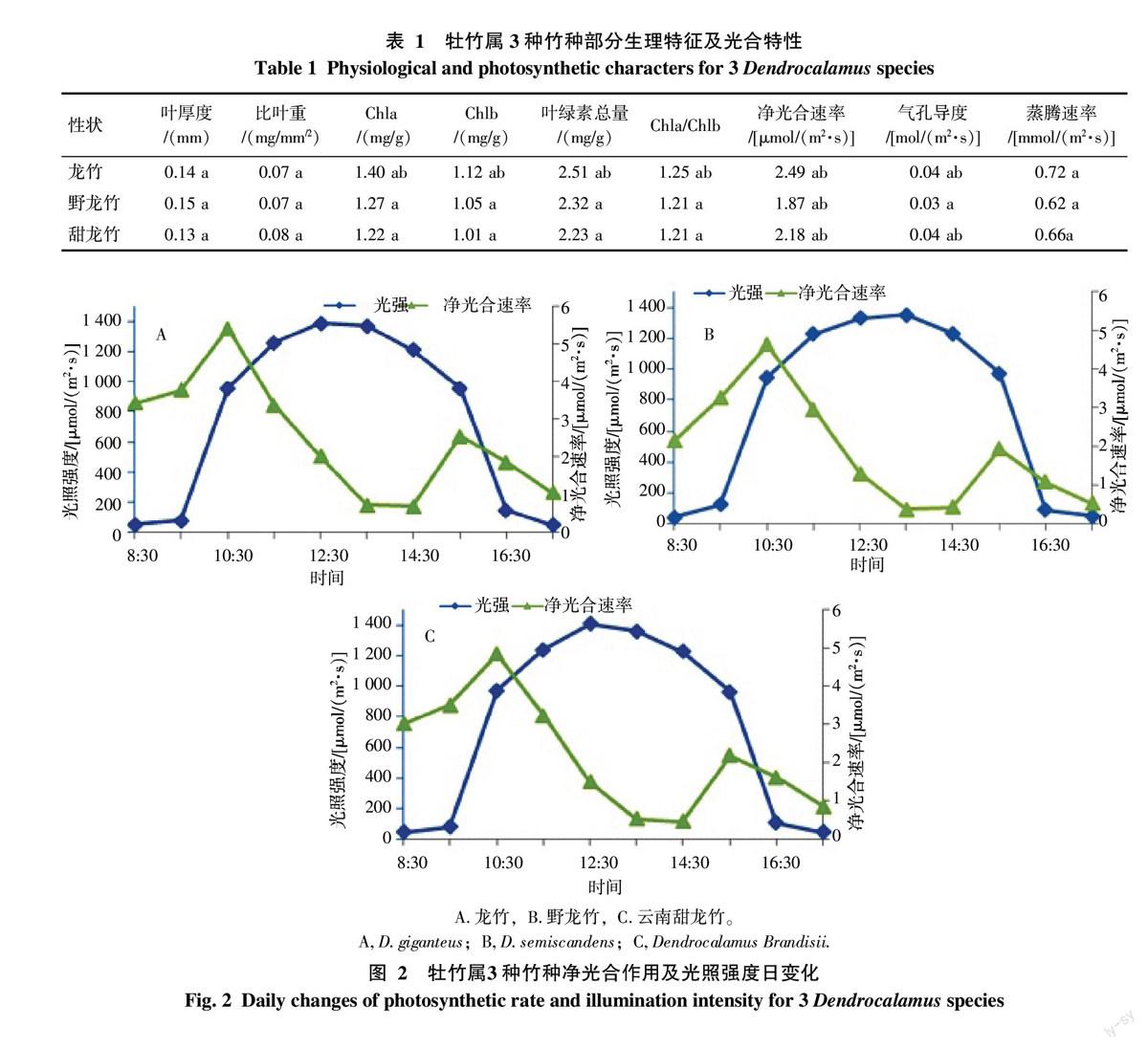
植物光合作用性能用叶片厚度和比叶重来衡量。从表1可知,野龙竹叶片厚度最大(0.15 mm),而比叶重相对小(0.07 mg/mm2);而云南甜龙竹叶片厚度最小(0.13 mm),比叶重最大(0.08 mg/mm2);龙竹叶片厚度相对居中(0.14 mm),而比叶重小(0.07 mg/mm2)。叶绿素是植物进行光合作用的主要色素,它在光合作用的光吸收中起核心作用[8]。从表1可知,就叶绿素总量而言,龙竹叶绿素总量最大(2.51 mg/g),野龙竹次之(2.32 mg/g),云南甜龙竹最小(2.23 mg/g)。而3个竹种的叶绿素a(Chla)含量与叶绿素b(Chlb)含量的比值相比,也是龙竹最大(1.25),野龙竹次之(1.213),云南甜龙竹最小(1.209)。
2.3 光合速率与影响因子的关系
2.3.1 光照强度对净光合速率的影响 如图2所示,龙竹、野龙竹及云南甜龙竹的净光合速率并非随着光强的增加而增加。在一定范围内,净光合速率确实与光强正相关,随光强的增强而提高。但当光强增强到一定范围,强光引起气温升高,大气水蒸气压亏缺的增大,使植株蒸腾速率持续增加。蒸腾速率的持续上升可导致叶片水势的下降,并引起气孔导度下降,从而导致净光合速率的下降。这就是净光合速率出现“午休”现象的原因。而14:30之后,随着气温和光强的降低和减弱,叶片的生理活动能力在适合环境下得到再次激发,净光合速率第二次上升,形成第二次高峰,直到光强逐渐减弱,最终导致光合速率的减弱。
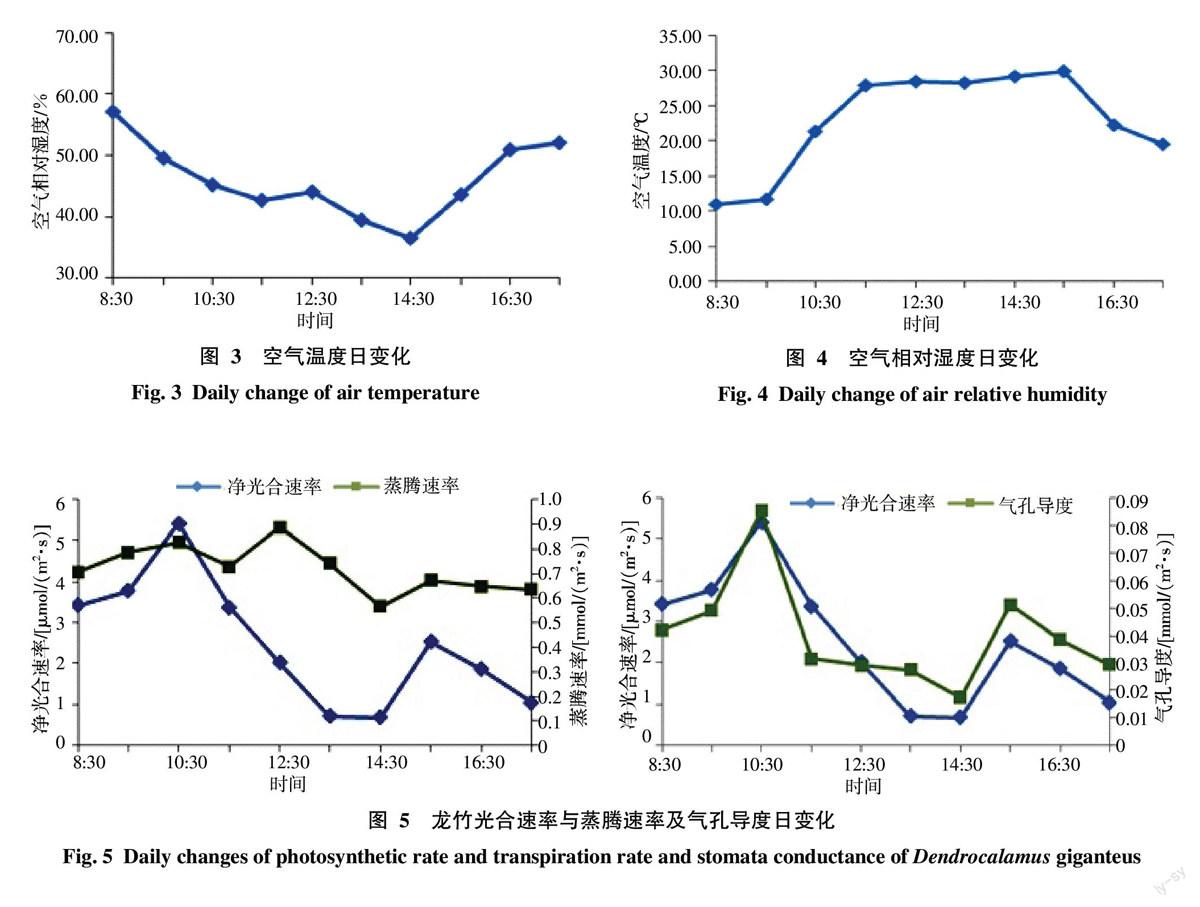
2.3.2 气温和湿度对光合速率的影响 由图3与图1可知,净光合速率从8:30时起,随着气温的升高而升高,10:30时当气温在21.30 ℃时达到最大值,中午前后受到高温的抑制而明显下降。在13:30~14:30时气温达29 ℃时,蒸腾速率持高,导致叶片水势及气孔度下降,光合速率下降到最低值。之后出现反弹,在15:30时达到第二次高峰。
由图3、图4和图1综合可知,空气相对湿度与空气温度的变化曲线相反,空气湿度在早晨和傍晚时最高,而此时3个竹种的净光合速率都是最低值,当净光合速率达最大时,空气相对湿度为45.21%。13:30~14:30空气相对湿度低于35%,湿度饱和差达到一天中的最大值,同时净光合速率达到一天中的低谷。
2.3.3 蒸腾速率和气孔导度对光合速率的影响
由图5可知,蒸腾速率在中午12:30达最高值,中午11:30开始,气温上升,到下午15:30到达最高值,而这期间,光照强度也在不断增强,并保持高值。但在中午12:30前后,由于光照强烈引起的午休现象,为保护叶片而降低叶片表面温度,蒸腾速率达到一天的最高峰,而净光合速率开始减低。
由图5可知,龙竹的气孔导度日变化与净光合速率日变化趋势基本一致,两者呈现明显的正相关。分别在上午10:30时与下午15:30时,气孔导度与净光合速率同时达到峰值,在14:30时,气孔导度与净光合速率也同时达到低谷。
2.4 光合性能相关性分析
为分析光合作用中各因素的相关性,利用龙竹叶片光合速率及其影响因子的数据,对光合速率(Pn)与气孔导度(Cond)、细胞间隙CO2浓度(Ci)、蒸腾速率(Tr)、气温(Ta)、大气CO2浓度(Ca)、相对温度(RH),光合有效辐射(Par)等变量进行多元回归分析,结果见表2。
由表2可知,龙竹光合速率与光合有效辐射、气孔导度、蒸腾速率呈极显著正相关,与气温显著相关,与细胞间隙CO2浓度极显著负相关。将龙竹光合速率(Y)与气孔导度(X1)、细胞间隙CO2浓度(X2)、蒸腾速率(X3)进行回归分析,得到光合速率与影响因子之间的回归方程:Y=13.236+65.447X1-0.0012X2+1.186X3(p<0.01,r=0.906),经t检验(t=1.20),偏回归系数具有显著性意义。
野龙竹和云南甜龙光合速率与影响因子之间的回归方程与龙竹的类似,分别是:
野龙竹: Y=24.226+45.142X1-0.006 7X2+2.141X3(p<0.01, r=0.896)
云南甜龙竹:Y=40.133+23.431X1-0.021X2+0.159X3(p<0.01,r=0.931)
总体来说,龙竹、野龙竹、云南甜龙竹3个竹种受植株自身因子的直接影响较环境因子更为明显。
3 讨论与结论
本研究结果发现,3个竹种光合速率出现“午休”现象,这在9个笋用竹种[3](慈竹Neosinocalamus affinins、梁山慈竹D. farinosus、桂竹Phyllostachys bambusoides、 刚竹P. viridis、 高节竹P. prominens、红哺鸡竹P. iridenscens、白哺鸡竹P. dulcis、毛竹P. pubescen、 假毛竹P. kwangsiensis), 以及4种地被竹[4](铺地竹Arundinaria argenteostriata、菲白A. fortunei、鹅毛竹Shibataea chinensis、黄条金刚竹Sasaella masamuneana f.aureostriata)的、四季竹Oligostachyum lubricum(Wen)King f.)[9]光合速率中都有出现。但是, 杜旭华等[10]在不同纬度引种地马来甜龙竹D. asper光合特性的研究中发现,纬度更高,温度和光强更相对较低的浙江省瑞安市引种的马来甜龙竹净光合速率日变化成单峰曲线,而福建省华安县,广东省广宁县引种地马来甜龙竹净光合速率日变化成双峰曲线,说明外部环境因子会影响竹类植物的净光合速率日变化。
在一定程度上,比叶重和叶绿素含量可以反映植物的光合速率,3个竹种叶片厚度依次为野龙竹(0.15 mm)>龙竹(0.14 mm)>云南甜龙竹(0.13 mm),而比叶重云南甜龙竹最大(0.08 mg/mm2);龙竹和野龙竹相对较小(0.07 mg/mm2)。说明云南甜龙竹较龙竹和野龙竹有较多的有机物积累,这与前人的研究相吻合,杨清等[11]研究表明云南甜龙竹叶片的生物量为8 071 kg/hm2,而唐建维等[12]研究表明龙竹叶片的生物量为 2 353.3 kg/hm2,但不能确定云南甜龙竹生产力就一定比龙竹和野龙竹大,因为其运输能力大小无法确定,三者生产同化物的能力叶不能确定[16]。同时,在一定程度上高的Chla/Chlb比值能够反映出高的光合速率,分析试验的结果可知,龙竹的叶绿素总量、叶绿素a和叶绿素b的比值都在3个竹种中最大,且前面分析龙竹的净光合速率也是最大的。由此,可以推断出,在一定程度上高的Chla/Chlb比值也能够反映出高的光合速率。
通过龙竹叶片光合速率与其影响因子相关性及回归方程分析,确定胞间CO2浓度、气孔导度和蒸腾速率是影响光合速率的主要因素,野龙竹与甜龙竹与此相似,这与前人研究结果一样[1,9]。在多个影响因子中,胞间CO2浓度对光合速率的直接影响最大,王亚萍等[7]对缺苞箭竹Fargesia denudata的光合速率研究表明光强和CO2浓度是影响植物光合生理的2个重要因子,气孔导度和蒸腾速率次之。蒸腾速率的增高,使得气孔导度降低,从而降低了净光合速率,也进一步表明了气孔导度是影响蒸腾速率和光合速率的重要原因之一。而气孔导度同时也是受到光照强度和蒸腾作用的影响,从而间接的影响光合作用进行[14]。光照强度是直接因子,影响大气温度和大气相对湿度,而这三者又对植株叶片的气孔导度和蒸腾速率产生很大影响,在研究结果表明龙竹光合速率与光合有效辐射、气孔导度、蒸腾速率呈极显著正相关,与气温显著相关。光照强度增加导致蒸腾作用加速,叶片为免受伤害,减低了气孔导度,使得光合速率减低。湿度饱和差能够加速蒸腾作用,降低叶片水势,进一步影响气孔的开度和光合速率[15-16]。环境因子和生理因子的复杂多变性,两者相辅相成,从而影响竹子的光合作用。因此在选育优良种源的时候,既要考虑竹种的品质的优良性,也要考虑生境条件是否适应,两者综合才能使生产力达到最大值。
参考文献
[1] Pinheiro C, Chaves M M. Photosynthesis and drought: can we make metabolic connections from available data[J]. Journal of experimental botany,2011,62(3):869-882.
[2] Zlatev Z, Lidon F C. An overview on drought induced changes in plant growth, water relations and photosynthesis[J]. Emir J Food Agric, 2012, 24(1): 57-72.
[3] 陈建华, 毛 丹, 朱 凡, 等. 9个笋用竹种的光合特性[J]. 中南林业科技大学学报, 2008, 28(6): 9-13.
[4] 刘国华, 王福升, 丁雨龙, 等. 4种地被竹光合作用日变化及光合光响应曲线[J]. 福建林学院学报, 2009, 29(3): 258-263.
[5] 谢寅峰, 杨万红, 杨 阳,等. 外源一氧化氮对模拟酸雨胁迫下箬竹光合特性的影响[J]. 生态学报, 2007, 27(12): 5 193-5 201.
[6] 李伟成, 王树东, 哲 科,等. 不同施肥梯度下酒竹的形态和光合响应[J]. 竹子研究汇刊, 2010, 29(2): 12-17.
[7] 王亚萍, 王开运, 张远彬,等. 缺苞箭竹气体交换对CO2浓度及光照强度的响应[J]. 西北林学院学报, 2008, 23(1): 6-9.
[8] 王俊刚,宋新青. 不同竹龄雷竹若干光合特性的比较研究[J]. 浙江林业科技, 2002, 22(1): 11-13.
[9] 李迎春, 李艳红, 郭子武,等. 夏季笋用竹种四季竹季节性光合生理特性[J]. 江西农业大学学报, 2011, 33(4): 719-725.
[10] 杜旭华,丁兴萃,陈 岩,等. 不同维度引种地马来甜龙竹Dendrocalamus.asper光合特性的比较[J]. 南京林业大学学报(自然科学版, 2012, 36(6): 53-57.
[11] 杨 清, 苏光荣, 段柱标,等. 版纳甜龙竹种群生物量结构及其回归模型[J]. 西北农林科技大学学报(自然科学版), 2008, 36(7): 127-134
[12] 唐建维, 萧自位, 王小李,等. 西双版纳不同林龄人工龙竹林的生物量及其模型[J]. 中南林业科技大学学报, 2011, 31(12): 54-60.
[13] 任 军, 范丽颖, 郭继勋,等. 花楸幼苗光合特性及其影响因子分析[J]. 东北师大学报, 2006, 38(2): 96-100.
[14] 关锦毅, 郝再彬, 张 达,等. 叶绿素提取与检测及生物学功效的研究进展[J]. 东北农业大学学报, 2009, 40(12): 130-134
[15] 金爱武, 郑炳松, 陶金星,等. 雷竹光合速率日变化及其影响因子[J]. 浙江林学院学报, 2000, 17(3): 271-275.
[16] 陈卫元. 锦竹光合特性的初步研究[J]. 江苏农业学报, 2008, 24(6): 901-905.
