秋石斛兰F1代主要观赏性状的遗传表现
2016-05-30陈和明吕复兵李佐肖文芳尤毅
陈和明 吕复兵 李佐 肖文芳 尤毅
摘 要 以秋石斛兰为亲本,配制成6个杂交组合,对杂交F1代的植株性状、花部性状和开花期进行统计分析。结果表明,杂种F1代的株幅、叶宽、花梗长、花径等4个数量性状的平均值分别占中亲值的99.63%、96.18%、97.01%和99.77%,均较中亲值有所下降,表现出一定程度的衰退现象,但由于组合内存在一定的变异幅度,选育植株较大、花径较大或花梗更长的个体是可能的;在植株的株高、叶长、单枝花朵数、花色及始花期等方面表现出超亲优势,尤其是单枝花朵数和花色遗传优势明显;在花色遗传上,无论是以白色花还是以红色系列花作母本,F1代均表现为红色系列且分离广泛,表明红色系列遗传能力更强。
关键词 石斛兰;杂交;观赏性状;遗传
中图分类号 S682.31 文献标识码 A
Genetic Performance of Main Ornamental Traits in
F1 Hybrids of Dendrobium hybrida
CHEN Heming1,2, Lü Fubing1,2 *, LI Zuo1,2, XIAO Wenfang1,2, YOU Yi1,2
1 Floricultural Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou, Guangdong 510640, China
2 Guangdong Provincial Key Laboratory of Ornamental Plant Germplasm Innovation and Utilization, Guangzhou,
Guangdong 510640, China
Abstract The genetic performance of some characters such as plant,flower and flowering period in F1 generation with six crosses in Dendrobium hybrida was studied. The results showed that the plant width, leaf width, peduncle length and flower diameter in F1 generation declined, whose mean was 99.63%, 96.18%, 97.01% and 99.77% of parents respectively, but the overparent individuals still existed in plant width or flower diameter or peduncle length. The genetic heritability of plant height, leaf length, flower number, flower color and early flowering period was strong, especially flower number and flower color. F1 generation showed red series and separated widely on the flower color whether maternal parents were white flowers or red ones, which suggested that the genetic ability of red color trait was stronger.
Key words Dendrobium; Hybridization; Ornamental characteristic; Heredity
doi 10.3969/j.issn.1000-2561.2016.09.002
石斛兰为兰科(Orchidaceae)石斛兰属(Dendrobium)多年生植物。栽培上把石斛兰分为温带型落叶种(春石斛兰)和热带型常绿种(秋石斛兰)[1]。其中,春石斛兰的花一般生于茎的各个节上,花枝长2~5 cm,花枝较短,着生1~3朵花,自然花期2~5月,单枝花期约20 d,主要作为盆栽及观赏;秋石斛兰的花主要着生于茎的顶部,花枝长50~100 cm,花枝较长,花序上着生十几朵或数十朵花,自然花期8~12月或更长,单枝花期1~2个月,是重要的商品切花和盆花两用的热带兰之一[2]。
长期以来,泰国、新加坡等国的兰花育种家通过长期的杂交育种,培育出许多秋石斛兰优秀品种,如萨宾(Dendrobium‘Sabine No.1)、大熊猫(Dendrobium‘Ekapolpanda No.1)、白花秋石斛(Dendrobium‘White)、复色秋花石斛(Dendrobium‘Ceaoai)等;我国在改革开放后才陆续引进秋石斛兰,起步较晚,同时由于地域气候的限制,造成秋石斛兰的规模化栽培发展缓慢,以及在育种方面的滞后[3-4]。近年来对秋石斛兰的研究报道主要集中于组织培养[5-8]、栽培管理[9-13]和工厂化育苗[14]等方面,在杂交育种和育种后代的性状分离及观赏性的遗传表现方面未见报道。因此,本试验以6个秋石斛兰杂交组合的F1代及其亲本的性状表现进行观察比较,研究杂交组合性状的遗传表现,以期为秋石斛兰杂交育种后代优良单株的选择和亲本的选配提供参考。
1 材料与方法
1.1 材料
2006年5月在广东省名优花卉种质资源圃内,选取编号D104、D105、D106、D107、D108的秋石斛兰优良品种,每品种切取部分幼嫩侧芽进行组培繁殖,并在2006年10月对以上5个亲本进行人工授粉杂交(表1)。2007年2月获得杂交蒴果并进行无菌播种,2008年5月获得的杂种后代(F1代)植株及亲本的克隆苗,数量为2 100株并进行种植观察,2010年9月F1代及亲本开花,10月进行观赏性状调查。
1.2 方法
从F1代的每个杂交组合中随机挑取100株,调查以下9个性状:株高、株幅、叶长、叶宽、花梗长、花径、花朵数、花色(分为花萼、花瓣颜色和唇瓣颜色)及始花期。其中,株高、株幅、叶长、叶宽、花梗长、花径、花朵数、花色等8个性状的测试方法参见石斛兰DUS测试指南[15]。
1.3 数据分析
利用Excel计算各性状平均值、标准差,并利用SAS软件进行变异系数分析[16]。
2 结果与分析
2.1 植株株高遗传
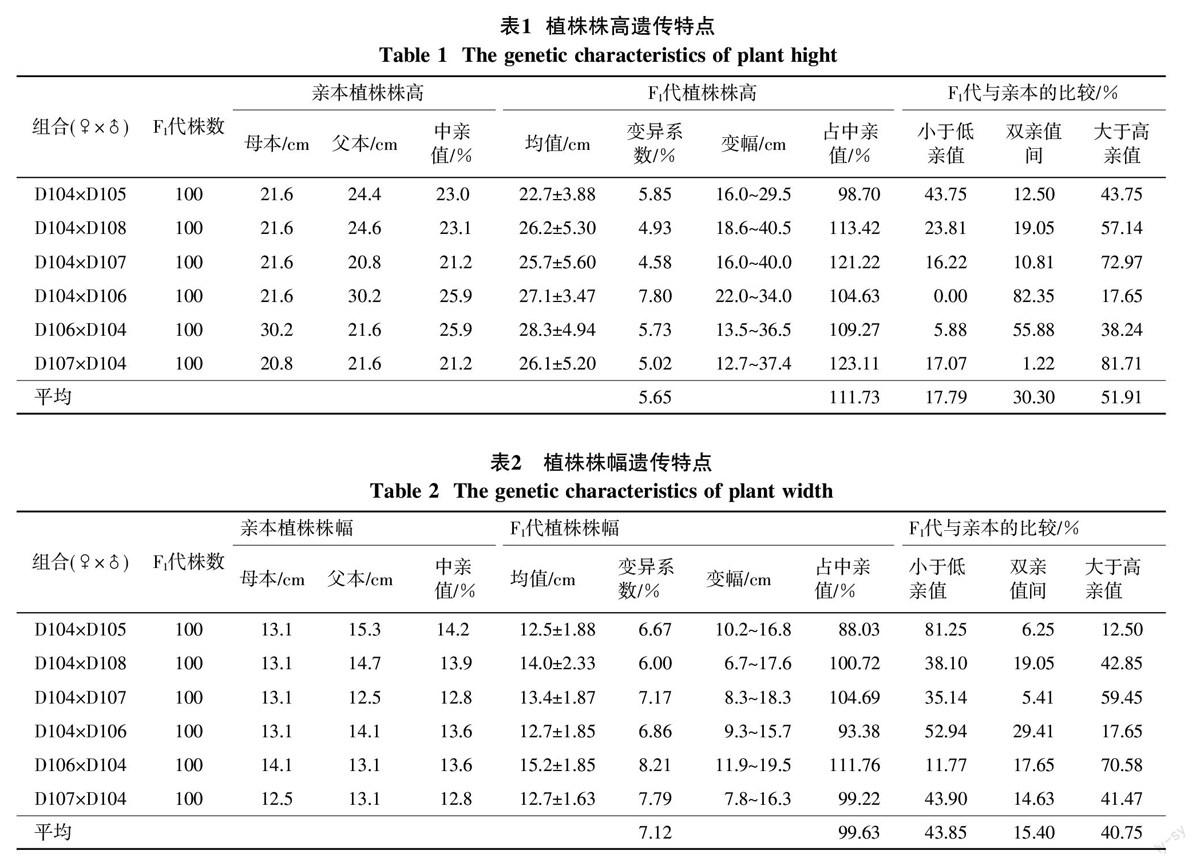
对杂交组合及其双亲的植株株高进行比较和统计分析,其遗传表现见表1。由表1可以看出,F1代的植株株高有增高的趋势,在6个杂交组合中5个组合的F1代株高均值高于中亲值,低于中亲值的仅有1个组合,在杂交组合的总平均株高相当于中亲值的111.73%,大于高亲值的个数占51.91%,小于低亲值的个体占杂种数的17.79%,表明F1代的植株比亲本明显增高,呈现出较大的超亲优势,同时组合内植株株高的变异系数为4.58%~7.80%,极值之差为12.0~24.7 cm。
2.2 植株株幅遗传
植株株幅遗传表现见表2。从表2可以看出F1代株幅总平均值相当于中亲值的99.63%,总体表现一定程度的衰退,但组合间存在差异,6个杂交组合中一半组合的F1代株幅均值低于中亲值,一半组合高于中亲值;其中,小于低亲个体比例为43.85%,大于高亲个体的比例占40.75%。但有的组合具有较大的遗传优势,如杂交组合D106×D104,超高亲个体占F1代总数的70.58%,组合内植株株幅的变异系数为6.00%~8.21%,极值之差为6.4~10.9 cm。
2.3 植株叶长和叶宽遗传
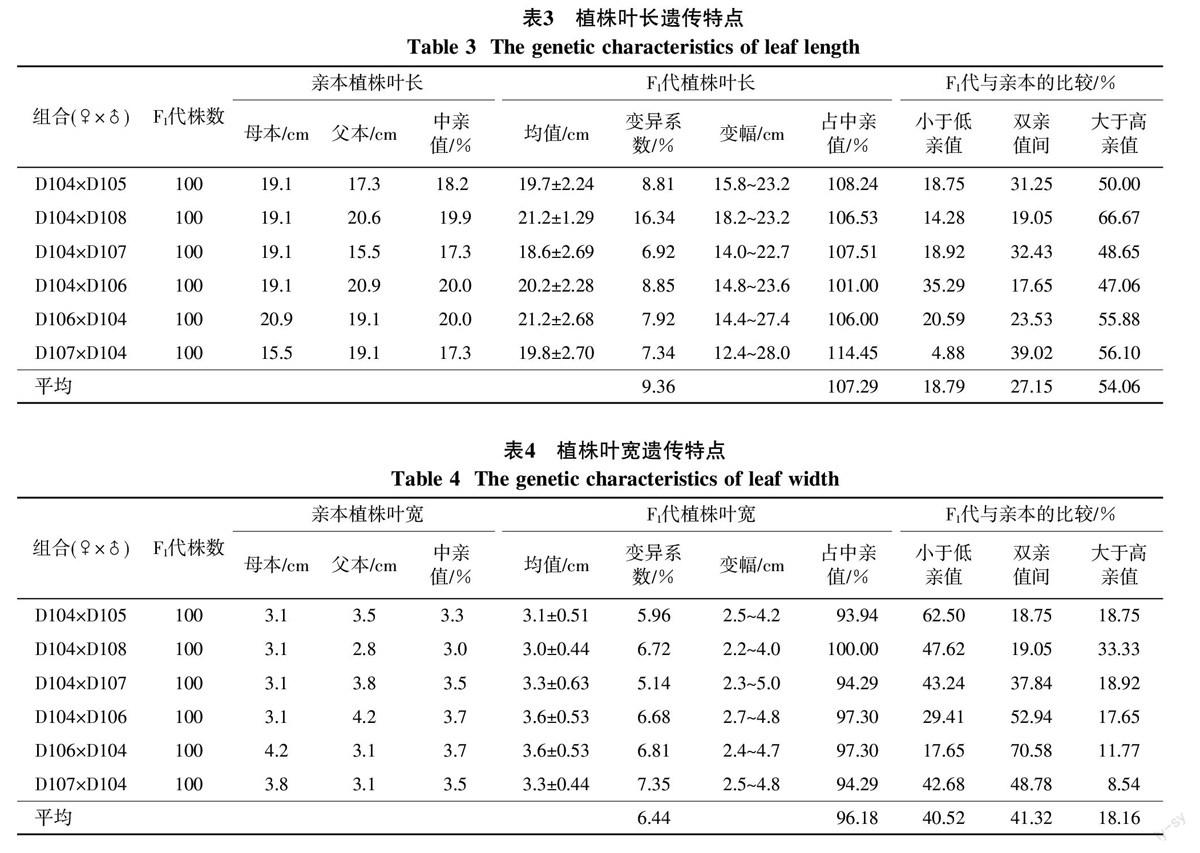
通过对F1代植株叶长、叶宽的统计分析,结果见表3和表4。从表3可知,F1代植株叶长呈现增大趋势,杂交组合的总平均叶长相当于中亲值的107.29%,大于高亲值的个数占杂种数的54.06%,小于低亲值的个数为18.79%,且6个杂交组合全部的F1代叶长均值均高于中亲值,表明F1代的植株叶长比亲本长,超亲优势明显(表3)。但F1代植株叶宽遗传明显低于叶长(表4),由表4可以看出,杂交组合的总平均叶宽相当于中亲值的96.18%,表现出明显的遗传衰退,小于低亲值的个体占杂种数的40.52%,介于双亲之间的占41.32%,大于高亲值的仅占18.16%,尤其是组合D104×D105叶宽的遗传衰退更为明显,其中小于低亲值的个体占杂种数的62.50%。从变异系数来看,叶长总平均变异系数为9.36%,比植株叶宽的总平均变异系数高出2.92%,表明F1代植株叶长分离较大。
2.4 花梗遗传
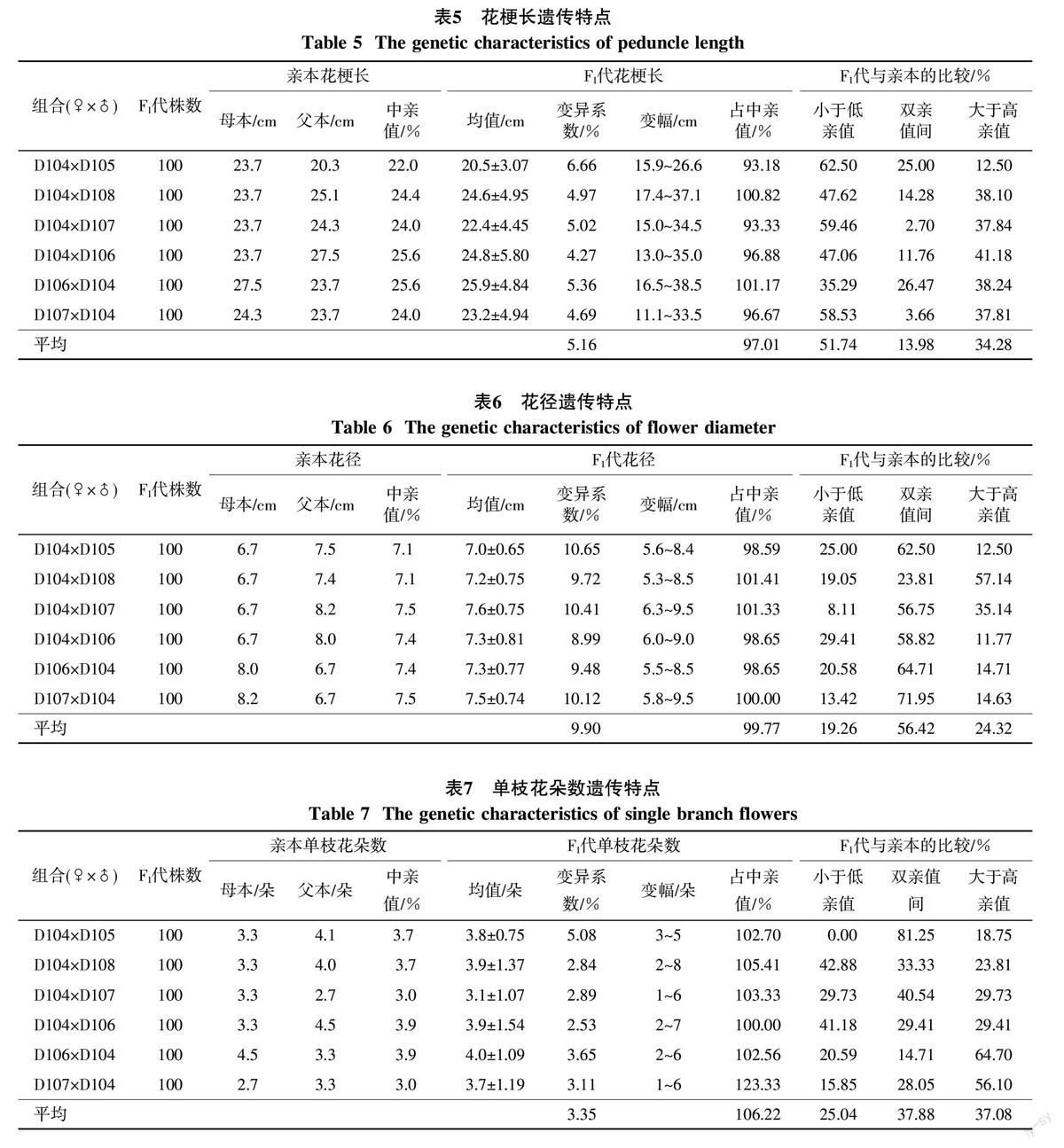
杂种F1代的花梗长与亲本比较表现出一定的衰退现象,但组合间存在差异(表5),表现在6个杂交组合中4个组合的F1代花梗长的均值低于中亲值,高于中亲值的仅有2个组合;在F1代植株总数中,花梗长总平均数相当于中亲值的97.01%,花梗低于双亲值的个体占总个体的51.74%,介于双亲值间的占13.98%,大于高亲值的占34.28%。
2.5 花径遗传
6个杂交组合总平均花径相当于中亲值的99.77%,但杂交组合间存在差异(表6),6个组合中一半组合的F1代花径均值高于中亲值,一半组合低于中亲值;在总的F1代植株中,花径低于双亲值的个体占总个体的19.26%,介于双亲值间的占56.42%,大于高亲值的占24.32%。
2.6 单枝花朵数遗传
由表7可知,杂种F1代花朵数表现出明显的杂种优势,杂交组合的总平均花数相当于中亲值的106.22%,小于低亲值的个体占杂种数的25.04%,介于双亲之间的占37.88%,大于高亲值的占37.08%,尤其是杂交组合D106×D104和D107×D104花数的遗传优势更强,表现为小于低亲值的个体占杂种数的20.59%和15.85%,大于高亲值的个体占杂种数的64.70%和56.10%,高出其正交组合D104×D106和D104×D107的35.29%和26.39%。
2.7 花色遗传
从表8可以看出,杂种F1代花萼、花瓣颜色分离广泛,由亲本的4种颜色:白色、红色/白色、紫红/白色、深紫红,分离出另外7种颜色且没有白色的出现,分别为粉红、红色、浅紫、紫红、粉红/白色、浅紫/白色、深紫/白色,但唇瓣颜色的分离较小些(表9),由亲本的4种颜色:白色、红色/白色、紫红/白色、深紫/白色,分离出另外5种颜色:浅紫、紫红、深紫、粉红/白色、浅紫/白色,且没有白色的出现。
由表8和表9可知,以白色花为母本,红色系列花作父本,其后代花色(包括花萼、花瓣颜色和唇瓣颜色)均表现为红色系列且分离更为广泛;以红色系列花作为母本,白色花为父本,其后代花色(包括花萼、花瓣颜色和唇瓣颜色)同样表现为红色系列且分离广泛,以上表明红色系列的花色遗传能力更强。
2.8 始花期遗传
杂种F1代的始花期见表10。从表10可以看出,F1代的始花期在9~11月均有,与亲本的始花期集中于9月相比,杂种F1代发生了分离。但无论是那个杂交组合,杂种F1代的始花期主要集中在9~10月份,始花的个体数分别占总个数的62.33%和26.00%,尤其是9月份始花的数量在60.0%以上,表明杂种F1代的始花期趋向亲本,但分离更为广泛。
3 讨论与结论
杂交是为获得具有优良性状或优势基因的一种有效方法,其中分析性状的遗传规律和不同性状杂种优势对选择亲本组合、提高育种效率及选育出性状优良的新品种具有重要意义[17]。在兰花方面,研究杂交后代遗传表现较多的报道仍然是蝴蝶兰,俞继英等[18-19]及陈和明等[20]均对蝴蝶兰杂交后代的开花植株进行了研究,但在石斛兰方面杂种后代遗传表现还未见报道。
本试验中通过对秋石斛兰亲本性状及其F1代的分离性状的研究,杂种F1代的株幅、叶宽、花梗长、花径等4个数量性状的平均值,均较中亲值有所下降,表现出一定程度的衰退现象,在观赏花卉非洲菊[21]、杜鹃[22]和百合[23]等上也有类似现象,其原因可能是经过有性过程后,优势降低了,致使杂种群体的性状平均值下降,由于组合内存在一定的变异幅度,选育植株较大、花朵较大或花梗更长的个体是可能的,主要通过改善栽培条件和提高管理水平,在同样衰退的程度下,仍可提高杂种平均值,从而提高选择率。但在植株的株高、叶长、单枝花朵数、花色及始花期等5个性状方面表现出超亲优势,尤其是单枝花朵数和花色遗传优势明显,其杂交组合的总平均花数相当于中亲值的106.22%,大于高亲值的占37.08%,而杂种F1代花萼、花瓣颜色由亲本的4种颜色分离出另外7种颜色,唇瓣颜色由亲本的4种颜色分离出另外5种颜色等,研究结果与陈和明等[20]一致。同时无论是以白色花为母本,还是以红色系列花作母本,其后代均表现为红色系列且分离广泛,表明红色系列的花色遗传能力更强,与董雪娜等[24]认为非洲菊杂交后代中白色与红色组合的亲本颜色在后代中无遗传优势不一致,可能与不同种类的花卉其遗传能力不同有关。
李娟娟等[17]认为遗传变异系数分为3个等级:0~15%(遗传变异度较小),15%~30%(遗传变异度中等),30%以上(遗传变异度较大),一般认为变异程度达到中等即可以认定该性状有较丰富的选择潜力。本试验中花色、始花期遗传变异较大,从亲本的4种颜色分离出另外7种颜色,始花期从亲本的9月到子代8~11月均有始花,但株高、株幅、叶长、叶宽、花梗长、花径、花朵数均表现出较小的遗传变异(表1~7)。总体来说,秋石斛F1代各组合内的数量性状的遗传表现较为一致,性状分离较小,但花色和始花期2个性状的F1代优势明显并产生了广泛分离,因此,选育出综合性状优良的秋石斛兰新品种是可能的。本研究中由于设置的组合在花色、花型等组配上还不够全面,需要进一步完善,以便深入了解秋石斛兰观赏性状的遗传特点,提高杂交育种效率。
参考文献
[1] 陈亚鸿, 洪 磊, 陈雄庭. 减少秋石斛在组织培养中的褐化研究[J]. 现代农业科学, 2009(3): 44-48.
[2] 卢思聪. 春石斛和秋石斛有哪些不同[J]. 中国花卉盆景, 2003(11): 4-5.
[3] 卢思聪. 热带洋兰秋石斛(上)[J]. 中国花卉盆景, 2003(9): 4-5.
[4] 陆顺教, 易双双, 任 易, 等. 秋石斛新生侧芽中部茎段组培快繁体系的建立[J]. 南方农业学报, 2015, 46(8): 1 436-1 441.
[5] 张晓申, 王慧瑜, 李晓青. 蝴蝶石斛的组培快繁技术研究[J]. 北方园艺, 2007(9): 193-194.
[6] 王云惠, 陈雄庭, 张秀娟. 蝴蝶石斛兰高效再生体系研究[J]. 广东农业科学, 2007(5): 46-48.
[7] 罗 岚, 关仕港, 刘建昌, 等. 秋石斛原球茎增殖培养[J]. 花木盆景, 2003(8): 4-5.
[8] 张 娟. 秋石斛组织培养的研究[J]. 湖北林业科技, 2015, 44(3): 20-22.
[9] 江秀娜, 李 军, 詹海洋, 等. 解读秋石斛的四大繁殖方法[J]. 中国花卉盆景, 2004(9): 38.
[10] 卢思聪. 秋石斛及其栽培[J]. 中国花卉盆景, 1999(7): 6-7.
[11] 武荣花, 李振坚, 王 雁. 秋石斛品种及其温室栽培[J]. 农业工程技术(温室园艺), 2007(1): 34-35.
[12] 邬秉左. 石斛兰的品种及栽培[J]. 花木盆景, 2002(3): 14-15.
[13] 杨志娟, 陈冠铭, 柯用春, 等. 热带洋兰秋石斛的主要病害及其防治[J]. 现代园艺, 2013(11): 66-67.
[14] 黄志明, 林庆良, 佘慧敏. 蝴蝶石斛兰工厂化育苗技术的研究[J]. 莆田学院学报, 2002, 9(3): 22-26.
[15] TG/209/1. Guidelines for the conduct of tests for distinctness, uniformity and stability(Dendrobium Sw.)[S]. Geneva, Switzerland: UPOV, 2003.
[16] 黄少伟, 谢维辉. 实用SAS编程与林业试验数据分析[M]. 广州: 华南理工大学出版社, 2001.
[17] 李娟娟, 陈洪伟, 王红利, 等. 一串红若干观赏性状在F1的遗传表现[J]. 华北农学报, 2014, 29(6): 113-120.
[18] 俞继英, 张 阳, 郑锦凯, 等. 蝴蝶兰黄花系品种Phalaenopsis ‘Taipei Gold 和P. ‘Sara Gold 杂交及其后代的遗传表现[J]. 浙江林学院学报, 2010, 27(4): 550-553.
[19] 俞继英, 郑勇平, 范文锋. 蝴蝶兰杂交育种研究进展[J]. 林业科技开发, 2009, 23(6): 5-10.
[20] 陈和明, 吕复兵, 朱根发, 等. 1个正反交蝴蝶兰若干性状在F1的遗传表现[J]. 华北农学报, 2011, 26(增刊): 28-33.
[21] 李绅崇, 李淑斌, 蒋亚莲, 等. 非洲菊品种间杂交主要观赏性状在F1代的遗传表现[J]. 云南农业大学学报, 2007, 22(2): 197-201.
[22] 刘晓青, 苏家乐, 陈 璐, 等. 杜鹃花品种(种)间杂交F1代主要观赏性状的遗传表现[J]. 江苏农业学报, 2012, 28(2): 459-460.
[23] 王秀刚, 胡翠平, 杨 涛, 等. 百合品种粉美与多安娜杂交F1代主要性状遗传分析[J]. 作物杂志, 2012(4): 90-94.
[24] 董雪娜, 陈 希, 蒋甲福, 等. 非洲菊F1代观赏性状的遗传表现[J]. 南京农业大学学报, 2015, 38(2): 226-232.
