20个小粒种咖啡种质生长量及根系形态差异分析
2016-05-30董云萍黄丽芳林兴军孙燕王晓阳陈鹏闫林
董云萍 黄丽芳 林兴军 孙燕 王晓阳 陈鹏 闫林
中国热带农业科学院香料饮料研究所农业部香辛饮料作物遗传资源利用重点实验室海南省热带香辛饮料作物遗传改良与品质调控重点实验室 海南万宁 571533
摘 要 以20个小粒种咖啡种质为试验材料,通过盆栽试验,分析咖啡种质苗期生长量和根系形态差异。结果表明,各种质苗期株高、叶片数、分枝数差异显著,通过苗期株形分析,把小粒种咖啡种质分成高干、中等和矮生3个类型。M13、M14可作为高干型,CATUAI为矮生型鉴定的标准品种;CA ZHONG Ⅲ、M13为高干、多叶、多分枝品种,CATURRA、T8667为株高中等、多叶、多分枝品种,热引2号为矮生、多叶、多分枝品种;各种质主根长除了CA ZHONG Ⅱ较小以外,其他种质间差异不显著,各种质主根直径差异不显著;M13侧根长、根体积、根表面积显著大于CATUAI,参试的小粒种咖啡种质绝大部分都具有细而且数量较多的侧根,各种质间差异较小。侧根直径与侧根长为显著的负相关,地上部干物质累积量与地下部干物质累积量间呈显著的正相关,长势强壮,枝叶繁茂的植株,根系发达;各种质地下部和地上部生长相协调的根冠比在0.40~0.35。
关键词 小粒种咖啡;植株形态;生物量;根系形态
中图分类号 S571.2 文献标识码 A
小粒种咖啡(Coffea arabica L.)原产于非洲的埃塞俄比亚山地雨林。中国最早1902年引进,品种有波帮和铁毕卡[1],直到20世纪50年代之前,一直是生产上的主要栽培品种。为了解决咖啡锈病、果腐病等生产问题,云南省德宏热带农业科学研究所和中国热带农业科学院香料饮料研究所通过国际合作项目出国考察、外国咖啡专家来华赠送等多种渠道引进了多个小粒种咖啡种质,建立了农业部瑞丽咖啡种质资源圃。收集的国外种质主要来自葡萄牙、肯尼亚、哥伦比亚、巴西、布隆迪、越南、马来西亚等国家,共计5个种377份[2-3]。国内对小粒种咖啡种质的农艺性状、产量性状、抗病性、杯品品质等进行了鉴定评价[4-9]。国外对咖啡种质的抗寒性、抗浆果病、抗根结线虫进行了评价[10-12],更多的研究集中在利用分子标记对小粒种咖啡种质的遗传多样性进行分析[13-21]。Daniel等[22]和Burkhardt等[23]研究了咖啡基因型与环境的相互作用,结果表明总蒸腾量较高的咖啡种质具有较高的根系生长量,Kufa[24]研究了埃塞俄比亚野生小粒种咖啡种群根生长特性,干旱地区小粒种咖啡种群相比湿润地区,有更高比例的细侧根,根体积、表面积大,是咖啡种质对干旱胁迫的响应机制。国内对小粒种咖啡种质根系形态的研究未见报道。本文对小粒种咖啡种质苗期地上及地下部形态、生物量进行分析,以期为种质性状的早期筛选提供可行方法。
1 材料与方法
1.1 材料
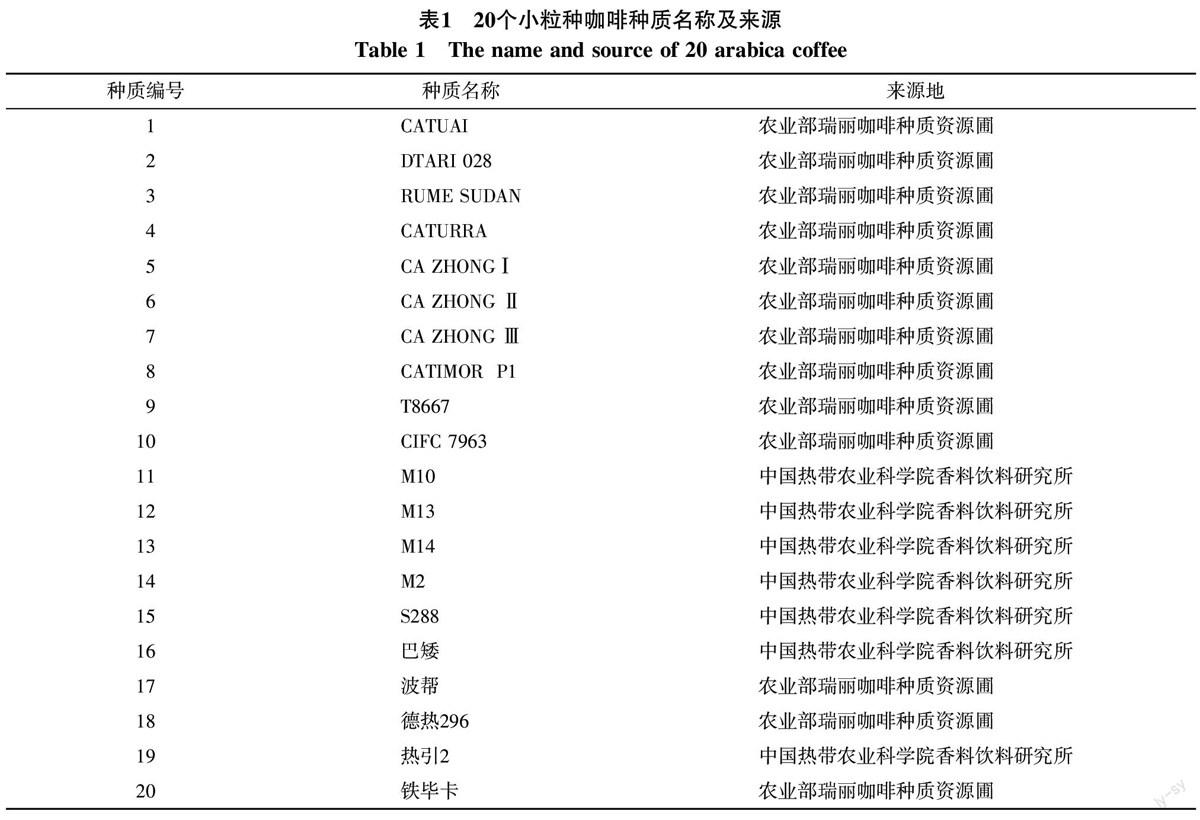
2013年1月从农业部瑞丽咖啡种质资源圃和中国热带农业科学院香料饮料研究所咖啡种质圃采集20个小粒种咖啡种质成熟鲜果,种质名称及来源见表1。
1.2 方法
1.2.1 试验处理方法 各种质咖啡鲜果经脱皮、脱胶、晾干后沙床催芽,待子叶平展时移植到直径10 cm营养袋培育,5对真叶时转入直径25 cm塑料花盆培育,培育基质为中细河沙,每个种质20盆。按改良霍格兰方法配制营养液,浓度为:Ca(NO3)2·4H2O 0.945 g/L、KNO3 0.506 g/L、NH4NO3 0.08 g/L、KH2PO4 0.136 g/L、MgCl2·6H2O 0.508 g/L、FeSO4·7H2O 5.56 mg/L。6月龄前每次每株施1/3浓度配制营养液300 mL,6~15月龄每次每株施配制营养液500 mL,2个月施1次。
1.2.2 测定方法 (1)生长量测定 选择每个种质生长一致的苗木15株编号,挂牌的每一植株即为一个重复,分别于苗龄6、12、15个月,测量株高、茎粗并记录叶片数和分枝数。
(2)根系形态及生物量的测定 苗龄15个月时从挂牌的每个种质中选生长一致的苗木3株,脱盆后,将根部的泥沙用自来水冲洗干净,带回实验室,每株分成叶片、茎干、根3部分,用吸水纸吸干根部水分后,分别称量各部分鲜重。将根系分成侧根和主根2部分,放置于有水的根盒中,让侧根舒展不重叠,用根系扫描仪扫描,WinRHIZO 2013分析主根长、侧根长、主根直径、侧根直径、根体积、根表面积,测定完成后将全株各部分放置于烘箱,105 ℃杀青30 min,75 ℃烘干至恒重,分别称其干重。
1.3 数据分析
试验数据均用Microsoft Excel进行整理,用SPSS统计软件进行方差分析。
2 结果与分析
2.1 不同小粒种咖啡种质株形分析
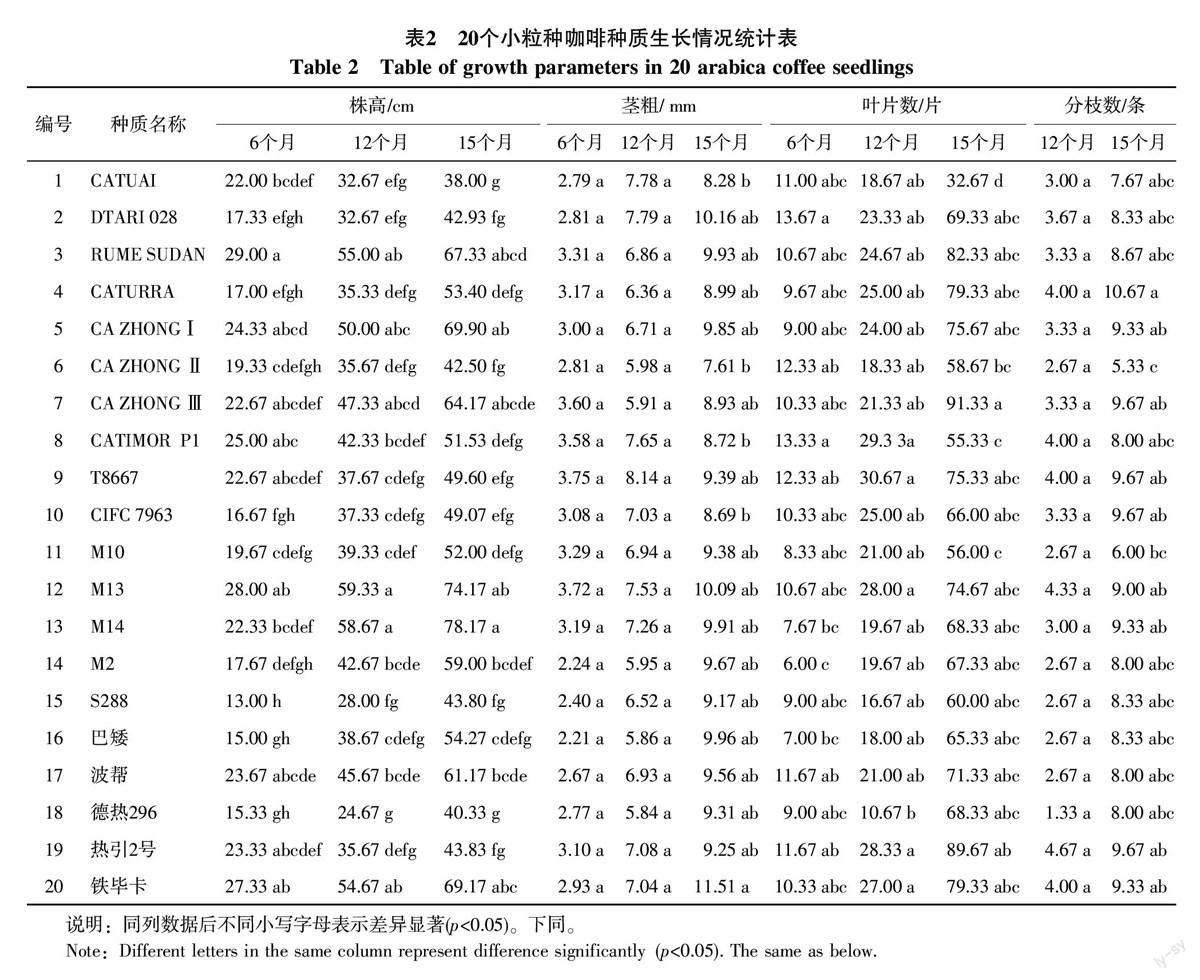
不同小粒种咖啡种质株高和叶片数生长量在苗龄6个月时差异显著,茎粗和分枝数生长量在苗龄15个月时差异显著,各种质在苗龄6个月时都还没有长出分枝。苗龄15个月时,M14、M13、CA ZHONGⅠ、铁毕卡、RUME SUDAN、CA ZHONG Ⅲ株高显著高于热引2号、S288、DTARI 028、CA ZHONG Ⅱ、德热296、CATUAI几个种质,波帮、M2、巴矮、CATURRA、M10、CATIMOR P1、T8667、CIFC 7963株高中等;铁毕卡茎粗显著粗于CATIMOR P1、CIFC 7963、CATUAI、CA ZHONG Ⅱ,其他种质间茎粗差异不显著。CA ZHONG Ⅲ叶片数显著多于CA ZHONG Ⅱ、M10、CATIMOR P1、CATUAI,其他种质之间叶片数差异不显著,叶片数较多的种质有CA ZHONG Ⅲ、热引2号、RUME SUDAN、铁毕卡、CATURRA、CA ZHONGⅠ、T8667、M13、波帮、DTARI 028几个种质。CATURRA分枝数显著多于M10和CA ZHONG Ⅱ,其他种质间分枝数差异不显著,分枝数较多的种质有CATURRA、热引2号、CIFC 7963、T8667、CA ZHONG Ⅲ、铁毕卡、M14、CA ZHONG Ⅰ、M13。咖啡株高中等或矮生、茎干粗、节密叶片数多、分枝多的种质树形紧凑,具有高产潜力,综合以上分析结果,株形较好的种质有CATURRA、T8667、CIFC 7963、热引2号、DTARI 028、波帮、德热296、巴矮。小粒种种质苗期株形选择宜在苗龄15个月进行,应综合株高、叶片数、分枝数几个指标进行筛选。
2.2 不同小粒种咖啡种质地上部生物量差异比较
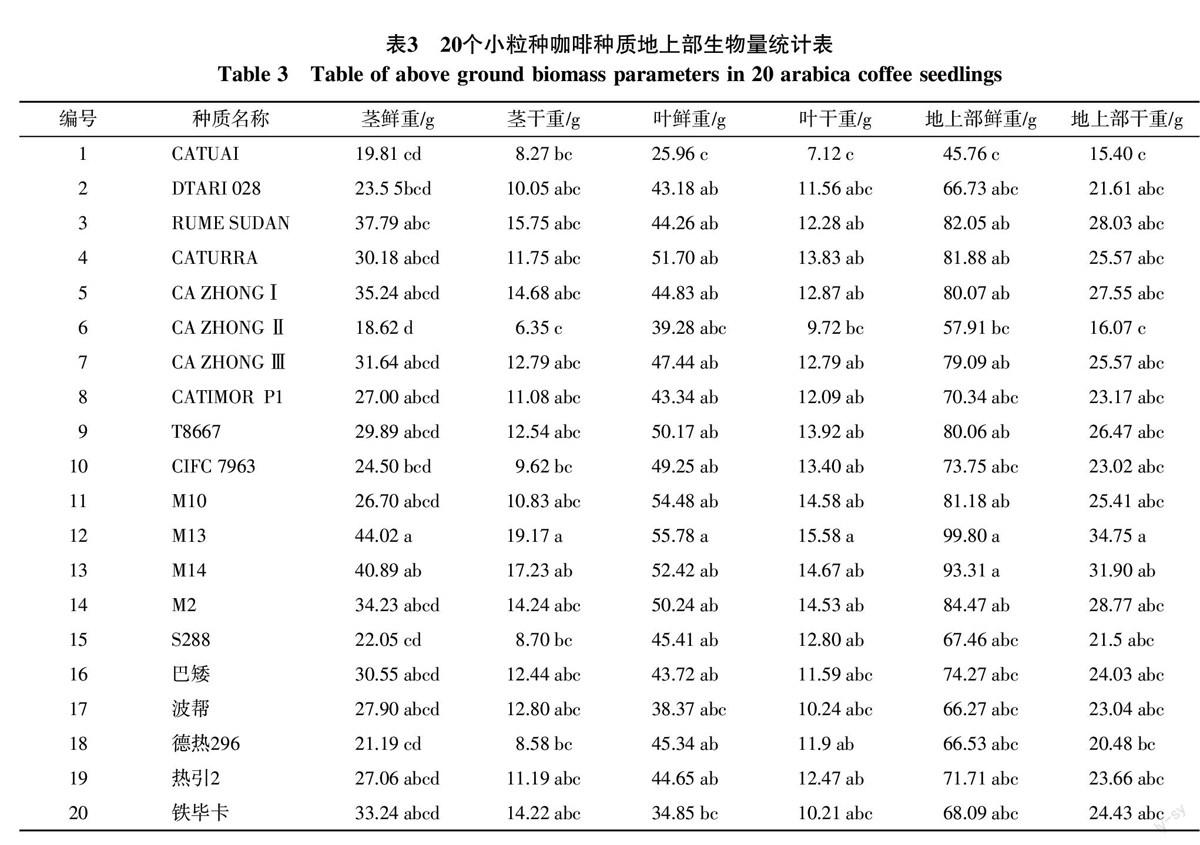
苗龄15个月不同小粒种咖啡种质生物量差异显著,M13茎鲜重、茎干重显著高于S288、德热296、CATUAI和 CA ZHONG Ⅱ,其他种质间差异不显著。茎鲜重、茎干重较重的种质有M13、M14、RUME SUDAN、CA ZHONGⅠ、M2、铁毕卡、CA ZHONG Ⅲ、巴矮、CATURRA、T8667;M13叶鲜重、叶干重显著高于CATUAI,叶鲜重、叶干重较重的种质有M13、M14、M10、CATURRA、M2、T8667、CIFC 7963、CA ZHONG Ⅲ;M13、M14地上部鲜重、干重显著高于CATUAI、 CA ZHONG Ⅱ,其他种质间差异不显著,地上部鲜重、干重较重的种质有M13、M14、M2、RUME SUDAN、CATURRA、M10、CA ZHONGⅠ、T8667、CA ZHONG Ⅲ。结合植株生长量指标可以看出,M13、M14、RUME SUDAN、CA ZHONGⅠ、CA ZHONG Ⅲ几个种质植株高,生物量累计大,而CATUAI、CA ZHONG Ⅱ、德热296几个种质植株矮生,叶片数少、分枝少,生物量累计小。M2、T8667、CATURRA、波帮、热引2号、巴矮植株高度中等,叶片数多,分枝数多,生物累计量也相对大。
2.3 不同小粒种咖啡种质根系形态分析
苗龄15个月不同小粒种咖啡种质主根长、侧根长、侧根直径、根体积、根表面积差异显著,主根直径差异不显著。CATIMOR P1、CATURRA主根显著长于CA ZHONG Ⅱ,其他种质间差异不显著。M13、T8667、S288、CIFC 7963、DTARI 028侧根显著长于CATUAI,CATUAI、CA ZHONG Ⅱ、RUME SUDAN侧根较少。M13根体积显著大于 CATUAI,其他种质间差异不显著。M13根表面积显著大于CATUAI、CA ZHONG Ⅱ、RUME SUDAN、波帮,其他种质间差异不显著。咖啡种质侧根数量在整个根系统中所占比例最大,侧根数量越多,根总长越长,根表面积、根体积越大。综合以上分析结果可以看出,M13、T8667、S288、CIFC 7963、DTARI 028、德热296、M2侧根数多,根体积、根表面积均较大,具有发达的根系。经双变量Pearson 相关性分析,侧根直径与侧根长的相关系数为-0.574,两者在0.01水平呈上显著负相关。CATUAI、CA ZHONG Ⅱ侧根直径显著粗于其他种质,但这2个种质的侧根长明显比其他种质小,而M13侧根明显比其他种质细,但其侧根数明显比其他种质多。此次试验的大部分种质都具有细而且数量较多的侧根。
2.4 不同小粒种咖啡种质地下部生物量差异比较
苗龄15个月不同小粒种咖啡种质根鲜重差异不显著,根干重差异显著。M13根干重显著大于 CA ZHONG Ⅱ和CATUAI,其他种质间差异不显著,根干物质累积量较大的种质有M13、T8667、CATIMOR P1、S288、巴矮、CATURRA、M2、DTARI 028,根干物质累积量较小的种质有CA ZHONG Ⅱ、 CATUAI。各种质根干重与地上部干重的比值即根冠比差异显著,S288、德热296、 T8667、CATIMOR P1、DTARI 028根冠比显著大于RUME SUDAN、M10、CA ZHONG Ⅲ、M14、CA ZHONG Ⅱ、CA ZHONGⅠ,后者根冠比较小的各种质中,CA ZHONG Ⅱ侧根数较少,其他几个种质由于植株高,叶片数多,其地上部干物质累积量相比地下部干物质累积量更大,因而他们的根冠比均较小。根冠比较大的几个种质根系发达,但植株相对矮生,因而根冠幅比大。综合各种质的性状分析可见,小粒种咖啡种质地下部和地上部生长相协调的根冠比在0.40~0.35。
2.5 小粒种咖啡种质地上部干物质累积量与地下部干物质累积量间的相关性分析
经双变量Pearson相关性分析,由表6可见,小粒种咖啡种质地上部干重与根干重相关系数为0.707,茎干重与根干重的相关系数为0.673,叶干重与根干重的相关系数为0.571,可见小粒种咖啡种质地上部干物质累积量与地下部干物质累积量之间在0.01水平上呈显著的正相关。
3 讨论与结论
苗龄15个月,通过苗期株形分析,把小粒种咖啡种质分成高干、中等和矮生3个类型。M13、M14可作为小粒种咖啡种质高干型鉴定的标准品种,CATUAI为小粒种咖啡种质矮生型鉴定的标准品种。在苗期,各种质茎粗除了铁毕卡相对较粗外,其他种质差异不明显,因而在苗期鉴定种质的茎干粗度性状并不理想。综合各种质的株高、叶片数、分枝数性状,其中CA ZHONG Ⅲ、M13为高干、多叶、多分枝品种,CATURRA、T8667为株高中等、多叶、多分枝品种,热引2号为矮生、多叶、多分枝品种,CATURRA、T8667、热引2号枝节密,分枝多,株形紧凑。根据萨尔瓦多咖啡协会[25]2005年的研究报告,波帮、铁毕卡植株高大,分枝长,节间间距长,成龄植株呈圆锥形。CATURRA是1935年在巴西的波帮咖啡种植园中突变出的更为矮生的品种,CATUAI是由Mundo Novo和黄果CATURRA杂交的品种。在中美洲种植园,由于CATUAI植株矮,为了获得和波帮品种相当的单位面积产量,种植密度为5 988株/hm2,波帮为3 333株/hm2。吴坤南[4]的研究表明M13、M14等品种植株高,树型松散。本研究通过苗期性状分析鉴定出的高干、中等和矮生3个类型品种,与上述研究结果一致。
地上部干物质累积量与地下部干物质累积量间呈显著的正相关,地上部生物量累积较高的品种为M13,累积量较小的品种为CATUAI和CA ZHONG Ⅱ。M13根干重显著大于CA ZHONG Ⅱ和 CATUAI,表明长势强壮,枝叶繁茂的植株,其根系发达。各种质根冠比差异显著,侧根数较少、植株较高或植株较矮生的种质根冠比较小或较大,地下部和地上部生长比例不协调,在本试验条件下,小粒种咖啡种质地下部和地上部生长相协调的根冠比在0.4~0.35之间。本研究通过综合分析,筛选出株高中等或矮生、节密叶片数多、分枝多、干物质累积量大、根冠比合理的品种如CATURRA、T8667、CIFC 7963、热引2号、DTARI 028等在王开玺[26]、吕玉兰[27]、周华[28]的研究中表明这些品种株高中等或矮生,分枝多,果节短,产量高,是生产上的主要栽培种。可见,通过苗期综合性状分析,筛选出具有高产潜力的品种的方法是可靠的。进行咖啡种质早期性状鉴定,可在苗圃设定一定的试验管理条件,对大批量的杂交后代或种质进行鉴定,具有占地面积小,试验观测劳动强度小、周期短的优点。
通过根系形态分析可见,苗期各种质主根长除了CA ZHONG Ⅱ较小以外,其他种质间差异不显著,主根直径差异不显著。侧根数量在整个根系统中所占比例最大,侧根数量越多,根总长越长,根表面积、根体积越大。侧根直径与侧根总长为显著的负相关,咖啡种质苗龄15个月时,已形成由一级、二级、三级侧根组成的复杂的根系统,二、三级侧根都非常的细而且长,往往二、三级侧根数量越多,根系越发达,侧根的平均直径就越细,反之侧根平均直径就越粗。M13侧根直径明显比CA ZHONG Ⅱ和 CATUAI细,但其根总长、根表面积、根体积明显大于CA ZHONG Ⅱ和CATUAI。Taye Kufa[24]研究结果表明咖啡种质侧根细,根体积、表面积大,是咖啡种质对干旱胁迫的响应机制。本研究筛选出M13侧根细,其根表面积、根体积比其他种质大,下一步对该种质的抗旱性进行验证,有望选出抗旱强的品种。
然而,试验结果也表明,参试的小粒种咖啡种质绝大部分都具有细而且数量较多的侧根,各种质间差异较小。由于小粒种咖啡属自花授粉,商业种植的品种多为波帮、铁毕卡及二者的突变种或种内杂交种,导致种质遗传多样性贫乏[13,17,29-31]。因而,在今后的工作中,收集更多的野生小粒种种质进行根系性状鉴定评价,是进行养分高效利用、抗寒、抗旱品种选育的基础。
参考文献
[1] 罗朝良. 宾川县志[M]. 昆明: 云南人民出版社, 1997: 135.
[2] 李锦红, 周 华. 德宏的咖啡选育种[J]. 热带农业科技, 2003, 26(4): 5-8, 14.
[3] 周 华, 张洪波, 李锦红, 等. 咖啡种质资源收集、保存、评价及创新利用研究[J]. 热带作物学报, 2012, 33(9): 1 554-1 561.
[4] 吴坤南, 龙宇宙, 张籍香. 墨西哥11号等小粒种咖啡在兴隆地区的试种表现[J]. 热带农业科学, 1992(3): 19-24.
[5] 陈振佳, 陈月梅. 咖啡种质资源收集与抗锈鉴定(初报)[J]. 热带作物研究, 1991(4): 32-36.
[6] 周 华, 李文伟, 张洪波, 等. 咖啡种质资源的引进、研究及利用[J]. 云南热作科技, 2002, 25(2): 1-6.
[7] 董云萍, 龙宇宙. 卡帝姆, 等. 几个小粒种咖啡在兴隆地区的引种试种[J]. 热带作物科技, 1999(5): 115-117.
[8] 高 敏, 张迎春, 陈 莹. 低温霜冻天气对咖啡杯品的影响[J].热带农业科技, 2015, 38(2): 29-32.
[9] 李学俊, 黎丹妮. 普洱咖啡产区主要咖啡种质资源的抗锈性评价研究[J]. 中国热带农业, 2016(2): 53-57.
[10] Marcos R P, Tumoru S, Marcos Z A. Genetic variability for frost resistance among Coffea accessions assessed in the field and in a cold chamber[J]. Brazilian Archives of Biology and Technology, 2005, 48(1): 15-21.
[11] Gichuru E K, Agwanda C O, Combes M C, et al. Identification of molecular markers linked to a gene conferring resistance to coffee berry disease(Colletotrichum kahawae)in Coffea arabica[J]. Plant Pathology, 2008, 57(6): 1 117-1 124.
[12] Villain L, Aribi J, eversat ′G R, et al. A high throughput method for early screening of coffee(Coffea spp.)genotypes for resistance to root-knot nematodes(Meloidogyne spp.)[J]. European Journal of Plant Pathology, 2010, 128(4): 451-458.
[13] Anthony F, Combes M C, Astorga C, et al. The origin of cultivated Coffea arabica L. varieties revealed by AFLP and SSR markers[J]. Theor Appl Genet, 2002, 104: 894-900.
[14] Missio R F, Caixeta E T, Zambolim E M, et al. Genetic characterization of an elite coffee germplasm assessed by SSR and EST-SSR markers[J]. Genet Mol Res, 2011, 10(4): 2 366-2 381.
[15] Manoj K M, Sandhyarani N, Jayarama. Genetic relationship among indigenous coffee species from india using RAPD, ISSR and SRAP markers[J]. Biharean biologist, 2011, 5(1): 17-24.
[16] Mishra M K, Suresh N, Bhat A M, et al. Genetic molecular analysis of Coffea arabica(Rubiaceae)hybrids using SRAP Markers[J]. Rev Biol Trop, 2011, 59(2): 607-617.
[17] Anthony F, Bertrand B, Quiros O, et al. Genetic diversity of wild coffee(Coffea arabica L.)using molecular markers[J]. Euphytica, 2001, 118(1): 53-65.
[18] 黄丽芳, 董云萍, 王晓阳, 等. 利用RAPD标记分析咖啡种质资源的遗传多样性[J]. 热带作物学报, 2014, 35(12): 2 313-2 319.
[19] Mulatu Geleta, Isabel Herrera, Arnulfo Monzón, et al.Genetic diversity of arabica Coffee(Coffea arabica L.)in Nicaragua as estimated by simple sequence repeat markers[J]. The Scientific World Journal, 2012(2): 1-11.
[20] Silvestrini M, Junqueira M G, Favarin A C, et al. Genetic diversity and structure of Ethiopian, Yemen and Brazilian Coffea arabica L. accessions using microsatellites markers[J]. Genetic Resources and Crop Evolution, 2007, 54(6): 1 367-1 379.
[21] Aga E. Molecular genetic diversity study of forest coffee tree(Coffea arabica L.)populations in Ethiopia: implications for conservation and breeding[D]. Uppsala: Swedish University of Agricultural Sciences, 2005: 1 652-6 880.
[22] Daniel G, Venkataramanan D, Awati M G, et al. Studies on genetic variability for root characteristics and water use efficiency in robusta coffee[C]. Bangalore: Proceedings of 20th International Conference on Coffee Science, 2004: 1 089-1 093.
[23] Burkhardt J, Kufa T, Beining A, et al. Different drought adaptation strategies of Coffea arabica populations along rainfall gradient in Ethiopia[C]. Montpellier: Proceedings of the 21st International Conference on Coffee Science, 2006: 1 032-1 036.
[24] Kufa T, Burkhardt J. Studies on root growth of Coffea arabica populations and its implication for sustainable management of natural forests[J]. Journal of Agricultural and Crop Research, 2013, 1(1): 1-9.
[25] Salvadoran Coffee Council. Exploring distinctive characteristics & virtues of coffee varieties: The Bourbon & Pacamara Case[EB/OL]. [2005-03-02].www.salvadorancoffees.com.
[26] 王开玺, 魏定耀, 陆明金, 等. 卡杜拉7号等小粒种咖啡品种比较试验[J]. 热带作物学报, 1991, 12(2): 17-22.
[27] 吕玉兰, 黄家雄.小粒种咖啡品种的灰色关联度分析[J]. 西南农业学报, 2014, 27(4): 1 393-1 398.
[28] 周 华, 夏红云, 李锦红, 等.优良咖啡品种-DTARI028选育报告[J].中国热带农业, 2010, 35(4): 49-51.
[29] Moncada P, McCouch S.Simple sequence repeat diversity in diploid and tetraploid Coffea species[J]. Genome, 2004, 47(3): 501-509.
[30] Aggarwal R K, Hendre P S, Varshney R K, et al.Identification, characterization and utilization of EST-derived genic microsatellite markers for genome analyses of coffee and related species[J]. Theoretical and Applied Genetics, 2007, 114(2): 359-372.
[31] Cubry P, Musoli P, Legnat′e H, et al.Diversity in coffee assessed with SSR markers: Structure of the genus Coffea and perspectives for breeding[J]. Genome, 2008, 51(1): 50-63.
