人工沙棘林灯下蛾类群落结构及时间生态位1)
2016-05-30门丽娜张志伟王利军郝淑莲韩有志
门丽娜 张志伟 王利军 郝淑莲 韩有志
(山西农业大学,太谷县,030801) (天津自然博物馆) (山西农业大学)
人工沙棘林灯下蛾类群落结构及时间生态位1)
门丽娜张志伟王利军郝淑莲韩有志
(山西农业大学,太谷县,030801)(天津自然博物馆)(山西农业大学)
摘要为了给人工沙棘林(Hippophae rhamnoides Linn.)的害虫防治和健康管理提供重要资料,选择山西农业大学林业站12年生人工沙棘林进行灯下蛾类群落连续调查,分析了人工沙棘林灯下蛾类的群落结构及时间格局。2013—2014年共采集灯下蛾类标本2 948头,隶属22科100种,卷蛾科、螟蛾科、夜蛾科、菜蛾科、尺蛾科为优势科,螟蛾科的物种数最多,为20种,卷蛾科的个体数最多,为655头。灯下蛾类无论是在科数、物种数还是在个体数上由高到低依次为8月份、7月份、6月份、5月份、9月份、10月份;Shannon-Wiener多样性指数、Pielou均匀度指数和Simpson优势度指数在一年中的不同时期没有明显差异,而Margalef丰富度指数与各月平均气温的变化趋势一致;7月份和8月份蛾类群落相似性最大,与10月份的群落相似性最小。人工沙棘林灯下蛾类优势科的时间生态位分析显示:卷蛾科、螟蛾科、夜蛾科、尺蛾科的时间生态位宽度均较大(>0.70),菜蛾科的时间生态位宽度较小(<0.50);卷蛾科、螟蛾科、夜蛾科、尺蛾科两两之间的时间生态位重叠指数均高于0.90,它们分别与菜蛾科的生态位重叠指数均小于0.85,说明菜蛾科物种的发生时间短于其他4科,较其他4科易于集中防治。
关键词人工沙棘林;灯下蛾类;群落结构;多样性指数;时间生态位
分类号S763.42;Q969.42
Characteristics of Community Structure and Temporal Niches of the Light Trapped Moth inHippophaerhamnoidesLinn. Plantation//
Men Lina, Zhang Zhiwei, Wang Lijun
(Shanxi Agricultural University, Taigu 030801, P. R. China); Hao Shulian(Tianjin Natural History Museum); Han Youzhi(Shanxi Agricultural University)//Journal of Northeast Forestry University,2016,44(5):78-83.
The experiment was conducted to analyze the community structure and temporal pattern of light trapped moths in aHippophaerhamnoidesLinn. plantation for pest control and forest management. A 12-oldH.rhamnoidesplantation in Forestry Station of Shanxi Agricultural University was selected for continuous investigation of a population of light trapped moths. The 2 948 light trapped moths collected from 2013 to 2014 belonged to 100 species across 22 families, of which Tortricidae, Pyralidae, Noctuidae, Plutellidae, and Geometridae were the dominant groups. The highest number of moth species was Pyralidae, with 20 species, and the highest number of individuals was Tortricidae, with 655 individuals. The biodiversity indices of light trapped moths in theH.rhamnoidesplantation were calculated and analyzed. The number of the light trapped moths caught varies with temperature (August>July>June>May>September>October), and this pattern is conserved across groups, species, and individuals. Shannon-Wiener’s index, Pielou’s index, and Simpson’s index did not vary across different periods in a year, whereas, Margalef’s index had the same trend with the average temperature of each month. The moth community similarity index from July and August was the highest whereas the moth community similarity index from July and October was the lowest. Analysis of the temporal niche of the dominant light trapped moths group showed that Tortricidae, Pyralidae, Noctuidae and Geometridae had broad temporal niche (>0.70), whereas, Plutellidae had a narrow temporal niche (0.489). The temporal niche overlapping index between families paired with Tortricidae, Pyralidae, Noctuidae and Geometridae were all higher than 0.90 while every family paired with Plutellidae had less than a 0.85 temporal niche overlapping index. The occurrence period of the Plutellidae species was shorter than the other four families, and easy to control intensively.
KeywordsHippophae rhamnoides Linn. plantation; Light trapped moth; Community structure; Diversity index; Temporal niches
沙棘(HippophaerhamnoidesLinn.)是防风固沙、保持水土和改良土壤的先锋树种[1]。我国是世界上人工沙棘林建设最有成效的国家之一,无论是在种质资源种类,还是分布面积上,都位居世界首位,已成为区域经济发展和维护生物多样性的重要生物资源[2]。人工沙棘林的健康管理在西北半干旱地区具有重要的经济和生态价值。蛾类是人工沙棘林生态系统的重要组成部分,在沙棘林害虫中占有重要地位,从沙棘叶片、枝干到根部都有蛾类幼虫的危害;且蛾类对生态环境变化敏感,具有广谱的生物地理学和生态学功能[3],蛾类的种类和数量可以预示森林的环境质量,可以作为森林中指示鳞翅目物种多样性和森林植被组成变化的指示类群[4-7]。蛾类昆虫种群数量大,物种多样性高,对植物和气候变化敏感,易于灯诱调查,是生物多样性调查的合适类群之一[8-10]。目前已知为害沙棘的蛾类有9科20余种[11-14],研究人工沙棘林蛾类的群落结构和多样性,探讨蛾类做为指示生物判断沙棘林微生态健康状况的可行性;分析蛾类优势类群的时间格局,计算其时间生态位,比较蛾类优势类群的发生期、发生期的长短和不同优势类群发生期的重叠情况,为人工沙棘林蛾类害虫的防治时间及防治策略提供依据。
1研究地概况
研究地为山西农业大学林业站的12年生人工沙棘林,样地面积约0.4 hm2,近方形(65 m×60 m),株行距2 m×3 m,平均株高1.75 m。林下主要是菊科(Asteraceae)蒿属(Artemisia)、禾本科(Poaceae)和旋花科(Convolvulaceae)植物,沙棘品种有中国沙棘、楚伊、丰产、金色、向阳和巨人等多个品种。山西农业大学林业站地处山西省晋中(37°30′N,112°35′E),海拔800 m。属暖温带大陆性季风气候,降水主要集中在6—8月份,年均降水量约479.6 mm,年均蒸发量约1 718.4 mm;年均气温9.4 ℃,无霜期约151 d。
2研究方法
诱虫器是由深圳富巍盛科技有限公司生产的太阳能智能风旋吸入式灭虫器(型号FWS-SP16-0106-0812),诱虫灯电压12 V,功率8 W,波长320~680 nm,光控和雨控自动开、关,适于鳞翅目昆虫的诱集,诱捕面积可达0.27 hm2[15]。诱虫灯改造:移除风扇防止损坏标本,集虫器去底、连接黑色纸箱、箱内放置瓦楞纸做为诱蛾陷阱。
2013年4月下旬—2014年10月底,每隔2 d收集1次陷阱内和扫网捕捉诱虫灯灯光可见范围内的所有昆虫,如遇天气不良则顺延一日,同一诱虫点连续采集3次,根据5点取样法,顺序更换太阳能诱虫灯的位置。4月下旬开始灯诱,因气温处回升阶段,波动较大,5月份开始气温较为稳定,故4月下旬的灯诱数据全部计入5月份。因每个季节昼夜节律不同,5—8月份20:00—22:00采集,9—10月份19:00—21:00采集。蛾类标本及时于实验室内制作成针插标本,待自然干燥后加采集标签(采集时间、地点和采集人)、按科分类整理并装入标本盒。依据外部形态及外生殖器特征分类鉴定到种级水平,主要参考《中国动物志》[16-21]、《中国麦蛾:一》[22]、《Oecophoridae of China:Insecta:Lepidoptera》[23]、《河南昆虫志》[24-25]、《河北动物志》[26]、《秦岭小蛾类》[27]等专著,所有标本保存于山西农业大学林学院昆虫标本室。
调查数据应用Microsoft Excel 2010、SPSS 19.0统计分析软件记录和处理,合并两年相同月份的蛾类数据,综合分析优势科的时间生态位。多度的划分标准:个体数>10%,计为优势科(+++);个体数1%~10%,计为常见科(++);个体数<1%,计为稀有科(+)。
采用如下生物多样性指数来评价人工沙棘林蛾类群落多样性:
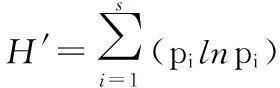
J=H′/lnS。

R=(S-1)/lnN。
式中:H′为Shannon-Wiener多样性指数[28]444-445;J为Pielou均匀性指数[29];C为Simpson优势度指数[28]440-444;R为Margalef丰富度指数[30]。pi=ni/N,为第i种的个体数占所有物种总个体数的比例,即:ni=第i种的个体数,N为所有物种个体总数;S为群落中的总物种数。
以下式计算不同季节蛾类群落的相似性:
Cs=2J/(a+b)。
式中:Cs为Sorensen相似性系数[31],取值0~1,Cs越大则2个调查时间的群落之间越相似。a为调查时间A时蛾类群落所含物种数;b为调查时间B时蛾类群落所含物种数;J为调查时间A和B时蛾类群落的共有种数。
以下式计算主要蛾类物种的时间生态位宽度和它们之间的时间生态位重叠:

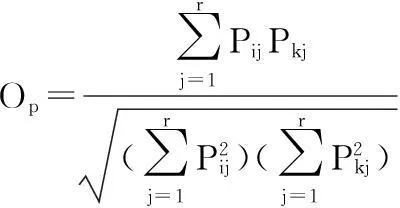
式中:Bi为物种i的时间生态位宽度[32];Op为物种i和物种k的时间生态位重叠指数[33]。Pij表示物种i利用第j等级时间资源占可利用总时间资源等级的比例;Pkj表示物种k利用第j等级资源占利用总资源等级的比例;r是该物种可利用的时间资源序列的等级数。
3结果与分析
3.1人工沙棘林灯下蛾类群落结构
整个调查期内共采集到灯下蛾类2 948头,隶属22科100种(表1),其中小蛾类占71%,表明人工沙棘林的蛾类物种十分丰富。在所采集到蛾类标本中,卷蛾科在个体总数上占总个体数的1/4,属优势科;其次是螟蛾科和夜蛾科。卷蛾科、螟蛾科、菜蛾科、夜蛾科、尺蛾科为研究区域的优势科;鞘蛾科、列蛾科、麦蛾科、谷蛾科、遮颜蛾科、细蛾科、羽蛾科为常见科;木蠹蛾科、祝蛾科、绢蛾科、织蛾科、尖蛾科、天蛾科、舟蛾科、毒蛾科、灯蛾科、刺蛾科为稀有科。
人工沙棘林的优势科所含物种最为丰富。卷蛾科物种数达13种,其中,宽小卷蛾(Olethreutestransversana(Christoph))、点基斜纹小卷蛾(ApotomiscapreanaHübner)、丽江柳小卷蛾(GypsonomarubescensKuznetsov)在数量上分别占卷蛾科个体总数的25.23%、18.55%和13.38%,为优势种。螟蛾科所含物种数达20种,其中库氏歧角螟(EndotrichakuznetzoviWhalley)是优势种,占螟蛾科个体总数的50.86%。夜蛾科所含物种达16种,其中二点委夜蛾(Athetislepigone(Moschler))(16.42%)、红棕灰夜蛾(Poliailloba(Butler))(18.12%)为优势种。尺蛾科包含11种,其中葎草洲尺蛾(Epirrhoesupergressa(Butler))(31.22%)和国槐尺蛾(SemiothisacinereariaBremer et Grey)(19.31%)是优势种。菜蛾科仅1种——小菜蛾(Plutellaxylostella(Linnaeus))。
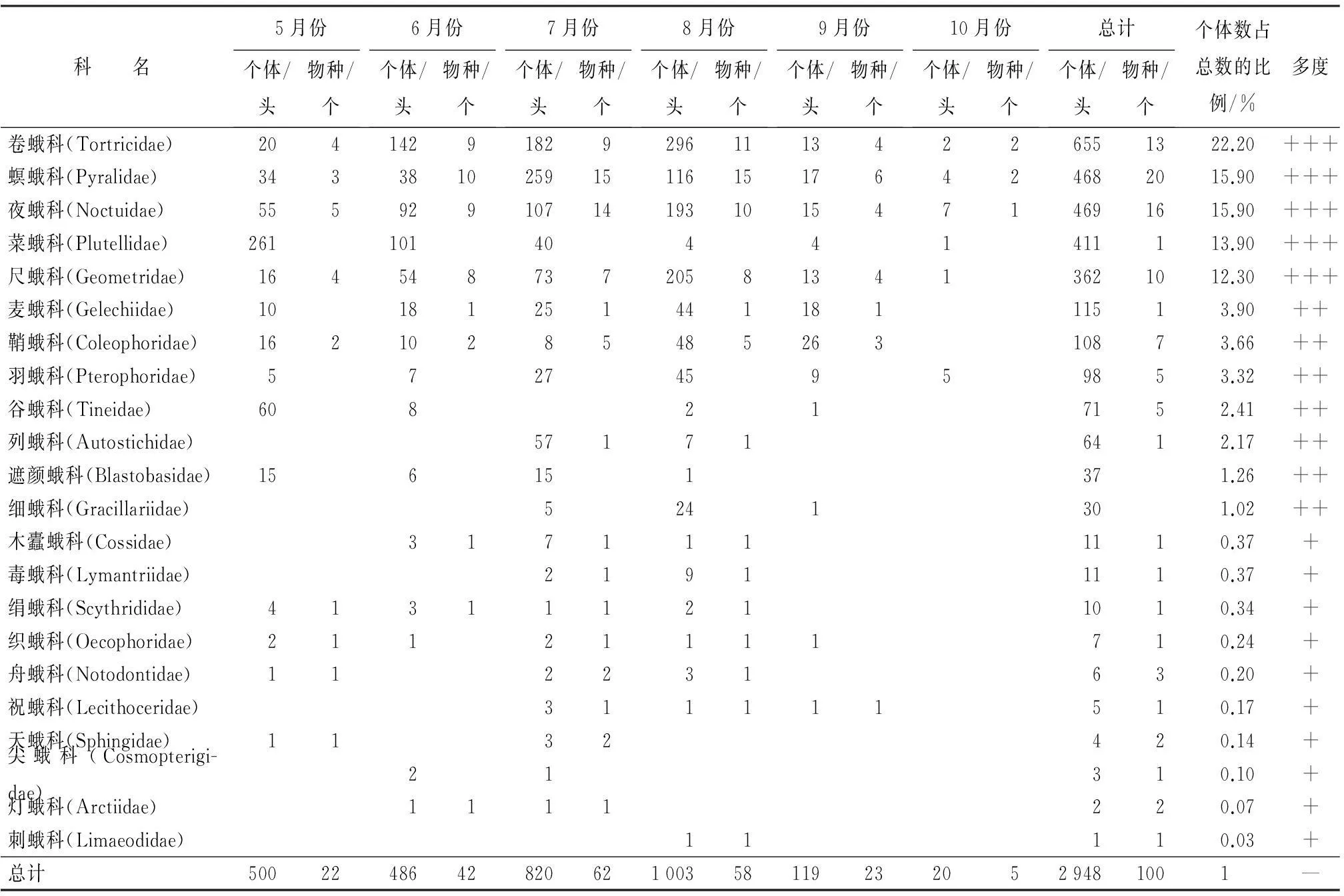
表1 人工沙棘林蛾类的物种组成(灯诱)
注:+++为优势科;++为常见科;+为稀有科。
3.2人工沙棘林灯下蛾类群落多样性
人工沙棘林灯下蛾类群落的Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)、Simpson优势度指数(C)在一年中的不同时期无明显差异,说明试验地蛾类群落较为稳定,没有蛾类害虫爆发为害;而Margalef丰富度指数(R)由高到低依次为7月份、8月份、6月份、9月份、5月份、10月份,即试验样地内7月份、8月份的蛾类丰富度最高,10月份最低,与各月平均气温的变化趋势一致(图1)。
人工沙棘林灯下蛾类群落的多样性指数从5月份至7月份逐渐升高,8月份到10月份逐渐下降,与丰富度指数的变化趋势大体一致;均匀度指数从5月份至7月份逐渐降低,8月份至10月份逐渐升高,既均匀度指数的变化趋势与多样性指数、丰富度指数的变化趋势相反;优势度指数5月份至7月份逐渐下降,7月份至10月份优势度指数相差不大。

图1 人工沙棘林蛾类群落多样性指数
3.3人工沙棘林灯下蛾类不同时期的群落相似性
人工沙棘林的蛾类不同时期群落相似性有一定差异(表2)。7月份和8月份的相似性系数最大,为0.871 8,表明这2个月的昆虫群落结构最相似,7、8月份是研究地平均气温最高的时期,人工沙棘林灯下蛾类不论个体数及物种数都最高,灯下蛾类的共有物种最多;7月份和10月份的相似性系数最小,为0.461 5,表明这两个月的昆虫群落结构相似性最小,灯下蛾类的共有物种最少。10月份研究地已进入秋季,气温渐低渐冷,许多优势科和常见科由于气温的下降逐渐减少甚至消失。10月份只有6个科,而7月份有20个科,2个时期在科数上相差最大,相似性系数也最低。
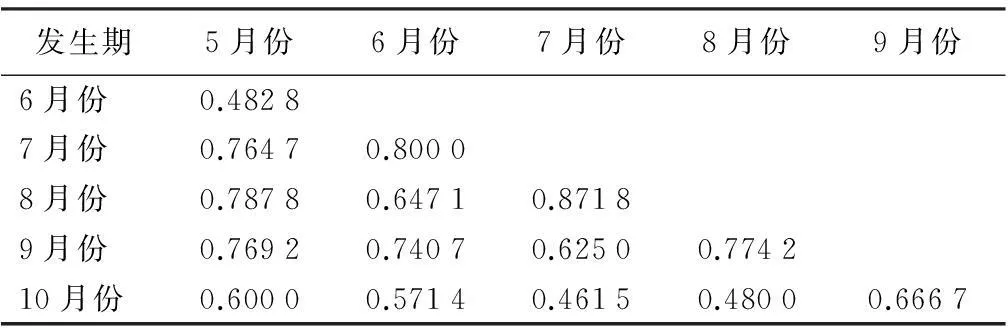
表2 人工沙棘林灯下蛾类群落相似性系数
3.4人工沙棘林灯下蛾类群落的时间格局和时间生态位
试验地的平均气温在调查期内先升后降,9、10月份平均气温低于其他时期,与此规律相对应,人工沙棘林灯下蛾类的群落组成及结构也随时间而变化,在个体数较多的科中表现尤为明显(图2)。5月份,灯诱得到的蛾类物种数和个体数逐渐增多,优势科和常见科的物种均已出现,菜蛾科和夜蛾科是这个时期的优势科;此期菜蛾科、谷蛾科在一年中个体数最多。6—8月份,蛾类的科数和个体数要明显多于其他时期,卷蛾科、螟蛾科、夜蛾科和尺蛾科是这个时期的优势科。9月份、10月份蛾类个体数均较少,一些科的个体数减少至0,如祝蛾科、织蛾科、列蛾科、细蛾科等。每个月都会灯诱得到卷蛾科、螟蛾科、尺蛾科、夜蛾科、菜蛾科、羽蛾科的个体。人工沙棘林一年内蛾类科数的消长变化总体上由高到低依次为7月份、8月份、6月份、5月份、9月份、10月份,个体数的变化趋势与科数的消长变化趋势基本相同,但7月份蛾类的物种数大于8月份,个体数明显小于8月份;6月份物种数略多于5月份,但个体数少于5月份。
人工沙棘林蛾类主要优势科的时间生态位宽度差别较大,从大到小依次为夜蛾科(0.740)、螟蛾科(0.717)、尺蛾科(0.712)、卷蛾科(0.704)和菜蛾科(0.489),表明优势科中夜蛾科、螟蛾科、尺蛾科和卷蛾科的发生期较长,在一年中的不同时期发生也较为均匀,而菜蛾科的物种在试验地调查期内发生的时间短于其他4科,在不同时期发生也不均匀,5月份为其发生的高峰(图2)。
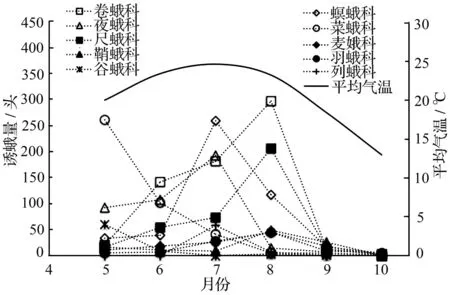
图2 个体总数超过50头的蛾类科时间格局
从时间生态位重叠指数来看,时间生态位宽度指数较高的夜蛾科、螟蛾科、尺蛾科和卷蛾科两两之间时间生态位重叠指数均高于0.90,但它们分别与时间生态位宽度指数较低的菜蛾科时间生态位重叠指数均小于0.85(表3),反映出菜蛾科在一年中的发生时间与其他4个优势科差别较大,而其他4科在一年中伴随发生的几率很大。
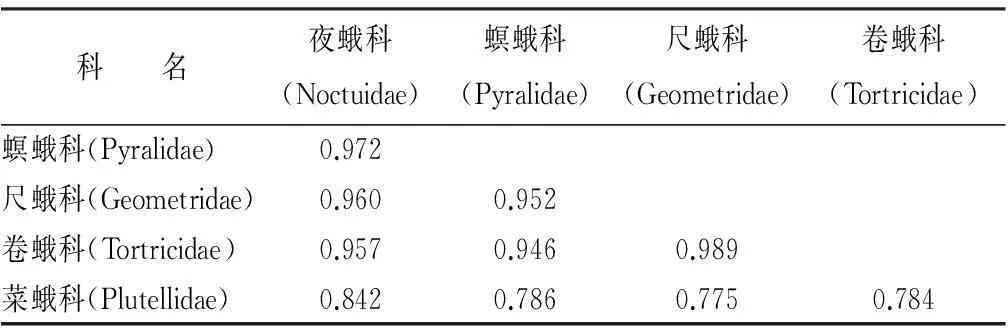
表3 人工沙棘林蛾类主要科的时间生态位重叠指数
4结论与讨论
蛾类群落结构可以反映其所属生态系统的健康程度,蛾类的群落结构往往较好地反应了蛾类对生态系统微环境变化的敏感性[6,34-35]。本研究中人工沙棘林蛾类的群落结构与时间格局同发生期平均气温的变化有密切的关系,说明蛾类作为生态系统健康的指示性生物是可行的,这与尤平等[34-35]、侯沁文等[6-7]的研究结果一致。昆虫作为其所在生态系统的重要成员,是生物多样性的主体之一,其群落结构的变动与生态系统的健康状况密不可分[36-38],昆虫群落的变动可以影响其所在生态系统食物网的组成,并直接或间接影响其寄主生物的分布与丰富度等[5],特别是优势类群可以作为生态系统的指示性生物[39]。本研究中,卷蛾科、螟蛾科、夜蛾科、菜蛾科和尺蛾科是人工沙棘林的优势科,在一年内不同时期的物种数及个体数均较高,用其数量、动态和结构作为反映人工沙棘林健康程度的指标,基础数据易采集,且在一年中的发生时期具有很强的规律性,故敏感性和可靠性强;这些蛾类在人工沙棘林生态系统中取食植物的叶片、花和果实等各种器官,并且为显花植物授粉等,维持该生态系统营养结构的稳定,在该生态系统的物质和能量转化中起着十分重要的作用,因此,可以作为该人工沙棘林生态系统健康的指示性生物。但对于如何建立量化的人工沙棘林生态系统优势科与该生态系统健康等级之间的对应关系,还需积累数据、进一步研究。
蛾类群落多样性与其所在生态系统的稳定性密切相关。人工沙棘林蛾类群落的多样性指数与丰富度指数的变化趋势大体一致,而与均匀度指数的变化趋势不同。人工沙棘林常常是典型的人为管护下的干旱、半干旱灌木林生态系统,存有的人为干预因素会在一定程度上影响人工沙棘林生态系统,导致蛾类群落结构变动,多样性指数与均匀度指数变化趋势不同,这与稻田昆虫群落和一些自然保护区蛾类群落的研究结果一致[6,40-41]。有研究表明,多样性指数与均匀度指数变化趋势一致的荒漠草原昆虫群落结构稳定[42];而一些自然保护区不同植被类型的蛾类群落研究表明多样性指数与均匀度不一致,群落所处的环境条件也较为优越[6,41];由此看来,多样性指数和均匀度指数的关系较为复杂,不但与所研究的类群有密切关系,还与研究地所处的地域、优势度指数等[41]也有关。
在我国北方,人工沙棘林所处环境往往为干旱、半干旱地区,这些地区的环境较为脆弱,在自然因素(如温度、湿度)波动的胁迫下,常会导致蛾类群落中一些物种的大发生,占据其他物种的生态位,使其个体数减少,从而群落中总的物种数也会减少[43]。从本研究对人工沙棘林蛾类群落结构分析结果看,试验地所处的生态系统蛾类优势科较多、物种丰富,且各科的物种在不同时期也相对均衡。说明人为管护对弱化环境因子对沙棘林蛾类群落的胁迫有积极作用。
群落相似性是反映群落物种组成相似程度的重要指标。研究地人工沙棘林7月份和8月份灯下蛾类的共有物种最多,相似性系数最大,表明这两个月的昆虫群落结构最相似;7月份和10月份灯下蛾类的共有物种最少,相似性系数最小,表明这2个月的昆虫群落结构差别最大。10月份,研究地已进入秋季,气温渐低渐冷,昆虫活动性降低,寻找越冬场所,许多蛾类的优势科和常见科由于温度的下降逐渐减少甚至消失,10月份只有6个科,而7月份有20个科,2个时期在科的数量上相差最大,相似性系数也最低。
时间生态位宽度指数大,说明该类群在一年中的发生时间长,在各个调查期均有发生,且各时期的数量分布也较为均匀;时间生态位宽度指数小,说明被调查类群发生期短、分布不均匀[7,44]。人工沙棘林灯下蛾类时间格局和时间生态位分析表明,卷蛾科、螟蛾科、夜蛾科、菜蛾科和尺蛾科等优势科的时间生态位宽度差别较大,卷蛾科、螟蛾科、夜蛾科和尺蛾科的时间生态位宽度均较大,分别为0.704、0.717、0.740、0.712,而菜蛾科的时间生态位宽度较小,为0.489。即,人工沙棘林内菜蛾科发生期短,且较为集中,较其他4科易于集中防治。生态位宽度和生态位重叠之间关系密切,生态位重叠指数高可以表现为生态位宽度“宽—宽”型或“宽—窄”型[7,45-46],人工沙棘林蛾类优势科的生态位重叠指数表现为典型的生态位宽度“宽—宽”型。
参考文献
[1]黄铨,于倬德.沙棘研究[M].北京:科学出版社,2006.
[2]刘书景,金幼菊,宗世祥,等.沙棘痕量挥发物采集方法探讨及成分分析[J].中国农学通报,2010,26(5):77-81.
[3]AZIZ K B. Pest management and the environment in 2000[M]. Wallingford, Oxon, U.K.: Oxford University Press,1992:293-311.
[4]张金良,李焕芳.秦岭自然保护区群的生物多样性[J].生物多样性,1997,5(2):155-156.
[5]王义平,吴鸿,徐华潮.以昆虫作为指示生物评估森林健康的生物学与生态学基础[J].应用生态学报,2008,19(7):1625-1630.
[6]侯沁文,铁军,白海艳.山西蟒河猕猴国家级自然保护区蛾类多样性[J].生态学报,2014,34(23):6954-6962.
[7]侯沁文,白海艳,铁军.蟒河猕猴国家级自然保护区蛾类群落生态位特征[J].生态学杂志,2015,34(4):1038-1045.
[8]HOLLOWAY J D. Insect surveys-an approach to environmental monitoring[C]. Roma: Atti Congresso Nazionale Italiano Entomologia,1980,121:231-261.
[9]HOLLOWAY J D. The larger moths of the Gunung Mulu National Park; a preliminary assessment of their distribution, ecology and potential as environmental indicators[J]. The Sarawak Museum Journal,1984,30(51):150-191.
[10]HOLLOWAY J D. Moths as indicator organisms for categorizing rain forest and monitoring changes and regeneration processes[C]//CHADWICK A C, SUTTON S L. Tropical rain forest: the leeds symposium. Leeds: Philosophical and Literary Society,1985:235-242.
[11]陈孝达,张学武.沙棘昆虫种群结构及综合防治研究[J].陕西林业科技,2000(2):36-39.
[12]陈孝达,党心德,李锋.陕西省沙棘昆虫区系的特点及分析[J].沙棘,2001,14(2):23-26.
[13]范仁俊,董晋明,曹满,等.山西省沙棘病虫草害及相关天敌的调查[J].沙棘,1994,7(3):10-13.
[14]同金侠,窦春蕊,陈孝达,等.陕西沙棘主要害虫及危害特点[J].国际沙棘研究与开发,2006,4(2):46-48.
[15]阿衣巴提·托列吾,穆肖云,张茂新,等.太阳能灭虫器对吐鲁番葡萄产区害虫的诱捕效果及对天敌安全性评价[J].应用昆虫学报,2012,49(4):1033-1042.
[16]陈一心.中国动物志:昆虫纲:第16卷鳞翅目:夜蛾科[M].北京:科学出版社,1999.
[17]薛大勇,朱弘复.中国动物志:昆虫纲:第15卷:鳞翅目:尺蛾科:花尺蛾亚科[M].北京:科学出版社,1999.
[18]方承莱.中国动物志:昆虫纲:第19卷:鳞翅目:灯蛾科[M].北京:科学出版社,2000.
[19]刘友樵,李广武.中国动物志:昆虫纲:第27卷:鳞翅目:卷蛾科[M].北京:科学出版社,2002.
[20]武春生,方承莱.中国动物志:昆虫纲:第31卷:鳞翅目:舟蛾科)[M].北京:科学出版社,2003.
[21]韩红香,薛大勇.中国动物志:昆虫纲:第54卷:鳞翅目:尺蛾科:尺蛾亚科[M].北京:科学出版社,2011.
[22]李后魂.中国麦蛾:一[M].天津:南开大学出版社,2002.
[23]WANG S X. Oecophoridae of China: Insecta: Lepidoptera[M]. Beijing: Science Press,2006.
[24]李后魂,任应党.河南昆虫志:鳞翅目:螟蛾总科[M].北京:科学出版社,2009.
[25]武春生,方承莱.河南昆虫志:鳞翅目:刺蛾科、枯叶蛾科、舟蛾科、灯蛾科、毒蛾科、鹿蛾科[M].北京:科学出版社,2010.
[26]李后魂,王淑霞.河北动物志:鳞翅目:小蛾类[M].北京:中国农业科技出版社,2009.
[27]李后魂.秦岭小蛾类:昆虫纲:鳞翅目[M].北京:科学出版社,2012.
[28]KREBS C J. Ecological methodology[M]. 2nd ed. New York: Harper & Row,1998.
[29]PIELOU E C.数学生态学[M].2版.卢泽愚,译.北京:科学出版社,1988:309-331.
[30]MARGALEF R. Perspective in ecological theory[M]. Chicago: University Chicago Press,1968:111.
[31]SORENSEN T J. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons[J]. Kongelige Danske Videnskabernes Selskab,1948,5:1-34.
[32]LEVINS R. Evolution in changing environments: some theoretical explorations[M]. Princeton: Princeton University Press,1968:39-53.
[33]Pianka E R. The structure of lizard communities[J]. Annual Review of Ecology and Systematics,1973,4:53-74.
[34]尤平,李后魂,王淑霞,等.天津七里海湿地蛾类多样性[J].昆虫学报,2003,46(5):617-621.
[35]尤平,李后魂,王淑霞.天津北大港湿地自然保护区蛾类的多样性[J].生态学报,2006,26(4):999-1004.
[37]EDGAR C B,BURK T E. A simulation study to assess the sensitivity of a forest health monitoring network to outbreaks of defoliating insects[J]. Environmental Monitoring and Assessment,2006,122(1/2/3):289-307.
[38]BROWN K S Jr, FREITAS A V L. Butterfly communities of urban forest fragments in Campinas, São Paulo, Brazil: Structure, instability, environmental correlates, and conservation[J]. Journal of Insect Conservation,2002,6(4):217-231.
[39]张红玉,欧晓红.以昆虫为指示物种监测和评价森林生态系统健康初探[J].世界林业研究,2006,19(4):22-25.
[40]万方浩,陈常铭.综防区和化防区稻田害虫-天敌群落组成及多样性的研究[J].生态学报,1986,6(2):159-170.
[41]问荣荣,马玲,刘哲强,等.小兴安岭森林恢复期不同植被区域蛾类组成及其多样性[J].生态学报,2016,36(15).doi:10.5846/stxb201501140116.
[42]贺答汉,田畴,任国栋,等.荒漠草原昆虫的群落结构及其演替规律初探[J].中国草地,1988,10(6):24-28.
[43]刘文萍,邓合黎.木里蝶类多样性的研究[J].生态学报,1997,17(3):266-271.
[44]马玲,顾伟,王利东,等.扎龙湿地的昆虫群落生态位[J].林业科学,2012,48(5):81-87.
[45]陈俊华,刘兴良,何飞,等.卧龙巴朗山川滇高山栎灌丛主要木本植物种群生态位特征[J].林业科学,2010,46(3):22-28.
[46]柴宗政,王得祥,张丽楠,等.秦岭山地天然油松群落主要植物种群生态位特征[J].生态学杂志,2012,31(8):1917-1923.
收稿日期:2016年1月11日。
第一作者简介:门丽娜,女,1981年12月生,山西农业大学林学院,山西农业大学林学博士后科研流动站。E-mail:linamen81@163.com。通信作者:韩有志,山西农业大学林学院,教授。E-mail:hanyouzhi@sxau.edu.cn。
1)国家自然科学基金项目(31402007);山西省林业厅林业关键技术攻关项目(201002);山西农业大学博士后基金项目(141668)。
责任编辑:程红。
