天然筇竹居群形态遗传多样性1)
2016-05-30董文渊邱月群王逸之杨奕胡戎
董文渊 邱月群 王逸之 杨奕 胡戎
(西南林业大学,昆明,650224) (青岛农业大学) (西南林业大学) (云南省农业科学院) (西南林业大学)
天然筇竹居群形态遗传多样性1)
董文渊邱月群王逸之杨奕胡戎
(西南林业大学,昆明,650224)(青岛农业大学)(西南林业大学)(云南省农业科学院)(西南林业大学)
摘要在野外调查的基础上,对云南省昭通市8个县和四川省2个县的10个筇竹自然居群的11项形态性状进行了研究,探讨其形态多样性水平。结果表明:在物种水平上各个性状存在较丰富变异,变异系数(CV值)为0.018~0.066 ;单因素方差分析显示,10项性状在居群间差异达到显著或极显著水平。从形态水平上揭示了筇竹遗传多样性,为筇竹种质资源保护提供理论与实践参考。
关键词筇竹;形态分化;遗传多样性
分类号S795;Q944.3+
Morphological Genetic Diversity of NaturalQiongzhueatumidinodaPopulations//
Dong Wenyuan
(South West Forestry University, Kunming 650224, P. R. China); Qiu Yuequn(Qingdao Agricultural University); Wang Yizhi(South West Forestry University); Yang Yi(Yunnan Academy of Agricultural Sciences); Hu Rong(South West Forestry University)//Journal of Northeast Forestry University,2016,44(5):101-103.
We studied 11 morphological characters of 10 naturalQiongzhueatumidinodapopulations in Yunnan and Sichuan Provinces by variance to reveal its genetic diversity at morphological level. There was a rich variation in the tested morphological characters with the variation coefficient of 0.018-0.066. By the variation analysis of single factor, there were significant differences or very significant difference in 10 characters among naturalQingzhueatumidinodapopulations.
KeywordsQiongzhuea tumidinoda; Morphological differentiation; Genetic diversity
筇竹(Qingzhueatumidinoda)又称罗汉竹,是禾本科竹亚科筇竹属植物,中小型混生竹类,秆高1.5~8.0 m,胸径0.5~3.0 cm[1]。筇竹笋味道甘甜鲜嫩,营养丰富,历来被视为山珍,其制品远销国内外;筇竹秆由于竹姿优雅秀丽,竹节鼓突,壁厚腔小,在庭院观赏和竹艺加工制作方面均有重要价值[2]。筇竹是国家三级保护的珍稀竹种之一,属西南地区特有种,主要分布在金沙江下游昭通市湿度线以北的8个县,其中大关县木杆镇和永善县细沙乡为滇东北地区筇竹现代分布中心区域,大关县筇竹资源面积达7 136.67 hm2,素有筇竹之乡的美誉,筇竹笋已成为当地林农经济收入的重要来源[3-4]。
从19世纪达尔文等发现物种变异到20世纪60年代,表型性状一直是研究物种变异和进化的标记。利用表型性状研究遗传变异快速简便,同时结合野外调查、标本采集等手段,可以在短期内对所研究物种的遗传变异水平有一个基本认识。因此,许多学者目前仍广泛应用表型性状揭示居群遗传变异及其格局[5-7]。表型变异包括表型变异格局和表型可塑性。表型变异格局通常指表型变异在居群内和居群间的分布。表型可塑性是指一个基因型在不同环境条件下表型表达的变化程度[8]。竹类无性系的形态可塑性是指其对环境的适应性,主要表现为无性系分株的数目、高度、胸径、分枝长度及角度、枝下高等方面的差异。
数十年来,筇竹的生长环境发生很大的变化,随着筇竹笋市场价格的大幅度上扬,对天然筇竹笋的过度开发,使得筇竹笋笋体小型化趋势加重,竹秆高度和粗度也不断下降,表明筇竹种群退化加快。筇竹是中山湿性常绿阔叶林群落组成的关键种、渐危种和特有种,是遗传多样性的重要载体,一旦灭绝,将使其生长的群落也趋于衰亡,筇竹及其他物种潜在的开发利用价值也将不复存在。文中从形态水平上揭示了筇竹遗传多样性,为筇竹种质资源保护提供理论与实践的参考。
1材料与方法
根据天然筇竹林自然分布情况,在金沙江沿岸选取10个乡镇进行调查取样(图1)。并对取样地的相关环境特征进行记录(表1)。
形态特征指标:调查时在每个筇竹居群中设置1 m×1 m的样方,在其中随机选取30株生长良好、无明显病虫害的植株,对每株个体测量其11项形态学指标,即株高、胸径、胸节长、分枝长、分枝角度、叶长、叶宽、叶基夹角、枝盘数、总节数、年龄。各指标用直尺、卷尺、游标卡尺等测量并计算。
数据处理:用Excel 2003与SPSS 18.0对数据进行统计分析处理。

图中画圈处为10个天然居群位置。

居群序号地 点海拔/m坡度/(°)坡向经度纬度1云南省镇雄县雨河镇茶坝村157710北偏西10°104°50'25.5″E27°45'02.4″N2四川省叙永县分水镇龙洞村169825南偏东40°105°14'23.88″E27°55'04.43″N3云南省威信县麟凤乡龙塘村大雪山林区141820北偏西10°104°47'00.55″E27°53'01.65″N4云南省彝良县小草坝镇三道村罗汉社202520南偏东30°104°17'58.72″E27°45'11.81″N5云南省盐津县中和镇中和村凉风凹138732北偏东40°104°00'24.58″E28°06'55.87″N6云南省水富县太平镇二溪村铜锣坝林场147235南偏西50°104°07'58.87″E28°25'00.50″N7云南省绥江县板栗镇罗坪村茶叶坳156015南偏东40°104°04'26.91″E28°22'40.80″N8云南省永善县细沙乡小岩坊自然保护区163145南偏东50°103°58'55.40″E28°15'49.89″N9四川省雷波县菁口乡双龙桥村199323北偏东45°103°43'26.42″E28°20'21.24″N10云南省大关县木杆镇银吉村134645北偏东15°104°00'05.19″E28°06'45.86″N
2结果与分析
2.1不同居群形态特征
不同居群的形态特征用形态特征指标来表示,主要包括株高、胸径、胸节长、分枝长、分枝角度、叶长、叶宽、叶基夹角、枝盘数、总节数、竹龄。用形态特征指标的变异系数来反映同一性状在不同居群中的变异幅度和不同性状在同一居群中的变异幅度的差异。从筇竹不同居群形态指标的变异系数(表2)可以看出,筇竹同一居群中不同性状的变异系数差别较大,同一形态性状的变异系数值在不同居群中存在一定程度的差异。11个形态性状变异系数的总体平均值差别较大,叶长的变异系数最小,为0.009;胸径的变异系数最大,为0.066。
10个居群形态指标变异系数的平均值都较大,达到了0.141以上。表明筇竹表型多样性较为丰富,其中,居群1的平均变异系数最大,为0.219;居群6的平均变异系数最小,为0.141。
2.2形态分化
单因素方差分析表明(表3),在调查的11个性状中,除叶宽在居群间的差异未达到显著水平外,其余10个形态性状在居群间的差异达显著水平(P<0.05)或达极显著水平(P<0.01),占总体的91.8%,说明居群之间出现了一定程度的形态分化。
通过比较F值发现,在居群间的差异程度最小的是叶宽(F=1.550),其变异系数也最小(CV=0.018),说明,叶宽对环境变化的反应相对迟钝,可以作为判断筇竹系统进化关系的一个重要指标。
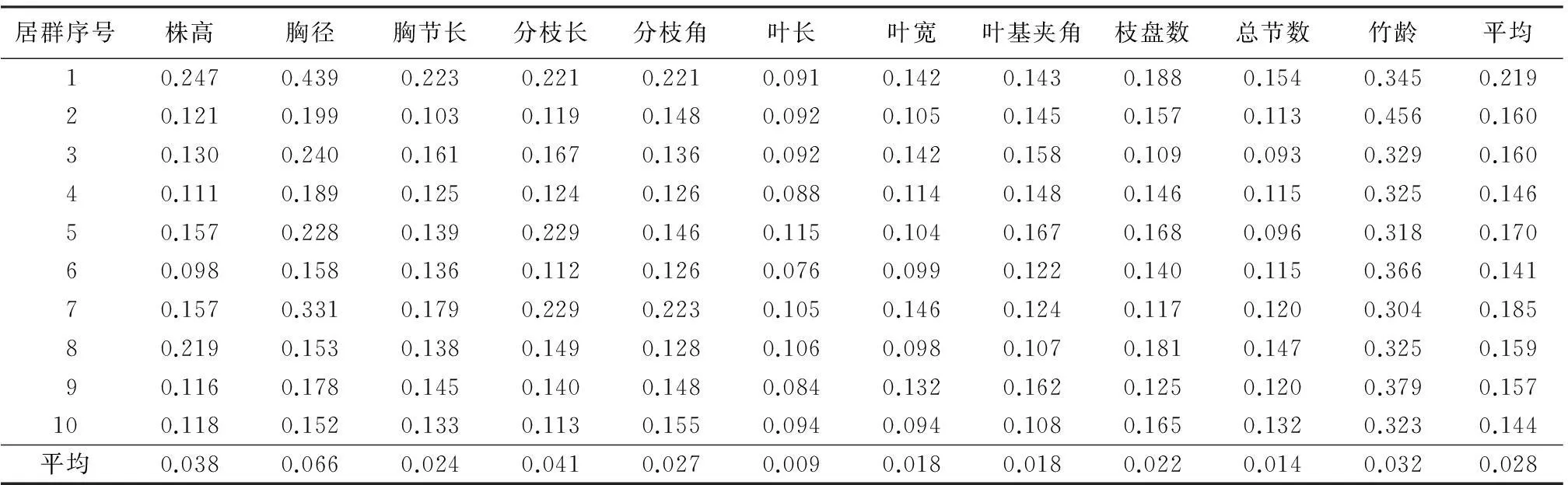
表2 筇竹不同居群形态指标变异系数

表3 筇竹形态特征的单因素方差分析
注:*表示差异显著;** 表示差异极显著。
3结论与讨论
筇竹在个体水平和居群水平上都出现一定程度的形态分化。从变异系数上看,每一居群的每一性状几乎都存在个体间的差异,在不同居群内,不同性状间其差异程度也不同。单因素方差分析显示,居群间出现一定程度分化,尤其在株高、胸径和总节数三个形态性状中差异水平达到极显著,这可能是由于受环境因子影响较大的原因造成的[9]。
筇竹的形态分化表明,该物种在居群间存在一定程度的形态异质性和遗传异质性。这种异质性一方面说明该物种仍有较大进化潜力,另一方面显示出筇竹对环境的变化反应比较敏感,形态可塑性强。野外资源调查发现,筇竹资源濒危状况日益严重,对其适宜生境的保护刻不容缓。筇竹对生境的要求比较严格,分布地域比较狭窄,生态极端脆弱,随着分布区内大面积无节制采笋作业等人类干扰活动的加剧,筇竹生境遭到很大程度的破坏,若不加强保护,筇竹无性系种群退化的速度将进一步加快。
参考文献
[1]耿伯介.中国植物志:第9卷:第1分册[M].北京:科学出版社,1996:349-356.
[2]董文渊.筇竹无性系种群退化及恢复机制的研究[D].北京:中国林业科学研究院,2006:12-18.
[3]杨奕,董文渊,邱月群,等.筇竹笋生长过程中营养成分的变化[J].东北林业大学学报.2015,43(1):80-83.
[4]董文渊,黄宝龙,谢泽轩,等.筇竹无性系种群生物量结构与动态研究[J].林业科学研究.2002,15(4):416-420.
[5]陈家宽.植物进化生物学[M].武汉:武汉大学出版社,1994:67-73.
[6]葛颂,洪德元.泡沙参复合体(桔梗科)的物种生物学研究:I表型的可塑性[J].植物分类学报.1994,32(6):489-503.
[7]JONAS C S, GEBER M A. Variation among population ofClarkiaunguiculata(Onagraceae) among altitudinal and latitudinal gradients[J]. Amerian Journal of Botany,1999,86(3):333-343.
[8]DUTILLEUL P, POTVIN C. Among-environment heteroscekasticity and genetic utocorrelation: implications for the study of phenotypic plasticity[J]. Genetics,1995,139:1815-1829.
[9]张桂萍,何平,邓洪平.濒危植物缙云卫矛的形态分化研究[J].西南师范大学学报(自然科学版),2001,26(6):703-708.
收稿日期:2016年1月15日。
第一作者简介:董文渊,男,1962年11月生,西南林业大学云南生物多样性研究院,教授。E-mail:wydong6839@sina.com。
1)国家林业公益性行业科研专项(201204103) 。
责任编辑:任俐。
