柏木低效林改造对土壤微生物、酶活性及养分的影响1)
2016-05-30韩东苗冯茂松吴韬李文兵铁烈华汪亚琳
韩东苗 冯茂松 吴韬 李文兵 铁烈华 汪亚琳
(四川农业大学,成都,610000)
柏木低效林改造对土壤微生物、酶活性及养分的影响1)
韩东苗冯茂松吴韬李文兵铁烈华汪亚琳
(四川农业大学,成都,610000)
摘要为了解不同改造模式对柏木低效林的改造效果,以德阳市旌阳区柏木(Cupressus funebris)低效林(简称纯柏)为对照,对4种改造模式(核桃+无植草、核桃+菊苣、核桃+苜蓿、核桃+鸭茅)的土壤微生物、酶活性及土壤养分进行了测定和分析。结果表明:与纯柏相比,4种改造模式均能不同程度提高土壤微生物数量、酶活性及养分(P、K)含量,各个改造模式中的土壤微生物总数、蔗糖酶、碱性磷酸酶及有效磷与纯柏差异显著;土壤微生物数量、酶活性及养分各指标含量间相关关系不同,其中,土壤微生物之间相关性不显著,土壤蔗糖酶与过氧化氢酶、脲酶呈显著正相关,其余酶活性之间相关性不显著,土壤有机质、全氮、碱解氮及pH之间、土壤全钾与速效钾、全磷及有效磷之间均呈显著正相关。土壤肥力分析表明,3种林草复合模式均能有效提高土壤肥力,核桃+菊苣模式改造效果最佳。
关键词柏木低效林;改造模式;林草模式;肥力分析
分类号S725
Effects of Transformation ofCypressfunebrisForest on Soil Microorganism, Enzymatic Activity and Nutrient//
Han Dongmiao, Feng Maosong, Wu Tao, Li Wenbing, Tie Liehua, Wang Yalin
(Sichuan Agricultural University, Chengdu 610000, P. R. China)//Journal of Northeast Forestry University,2016,44(5):57-62.
In order to understand the impacts of forest transformations onCypressfunebrisforest by several typical transformation patterns (walnut+no grass, walnut+chicory, walnut+alfalfa, walnut+orchardgrass), the soil microorganism, enzymatic activity and nutrient were determined and analyzed in different patterns of transformation whenC.funebrisforest being used as a control in Jingyang District, Deyang City. Four transformation models increased the amount of soil microorganism, enzyme activities and soil nutrient (P and K) in different degree compared withC.funebrisforest. The differences of total amount of microorganism between typical patterns of transformation andC.funebrisforest were significant, so were the soil sucrose activity, alkaline phosphatase activity and available P. There were different relationships among the soil microorganism, enzyme activity and nutrient, for example, the correlations among soil microorganism were not significant, and significantly positive correlation relationships between the soil sucrose and catalase, urease were observed, while there were no significant correlations among the rest of the enzyme activities, and there were significant positive correlations among organic matter, total N, hydrolysis N and pH. Total K had a significantly positive correlation to rapidly-available K, total P and effective P. Through the soil fertility analysis, three kinds of forest and herbage composite patterns could improve the soil fertility effectively, and the walnut+chicory was the best.
KeywordsCypress funebris forest; Transformation pattern; Forest and herbage pattern; Fertility analysis
柏木(Cupressusfunebris)林是四川盆地丘陵区的主要森林类型[1],也是该地区主要的低产低效林分[2]。目前,对柏木低效林分改造主要采取纯林补植其他树种变成混交林、改善经营管理措施等改造方法[3-5]。黎燕琼等[6]在川中丘陵区开展了以不同带宽强度的带状采伐和采伐带补阔的林分改造试验,结果表明,林分改造总体降低了林分的林冠截留量。把林下植被作为林分结构调整的因子,通过改造措施来对林下植被进行培育已经得到广泛地认可和应用[7]。但采用经济林改造模式的研究还不够广泛,所以本研究因地制宜栽植核桃经济树种,并结合林草复合模式,其改造模式对提高低效林的生态与经济效益具有一定的研究价值。土壤微生物、酶活性和养分是森林生态系统的重要组成成分,直接影响林木生长,参与或促进土壤中一系列复杂的生理生化反应,可以客观地反映土壤肥力状况[8]。本研究通过土壤微生物、酶活性及养分对比研究,评价该区柏木低效林采用不同核桃林草复合改造模式的土壤肥力变化情况,为该区筛选合理的改造模式提供理论依据。
1研究区概况
研究区位于德阳市旌阳区,地处成都平原边缘(东经104°15′~104°35′,北纬31°1′~31°19′)。市境内地形地貌多样,属典型的低山丘陵区。研究区位于亚热带湿润季风区,常年平均气温16 ℃,极端最高气温36.5 ℃,极端最低气温-6.7 ℃。年平均日照时间1 150 h,年平均降水量900 mm,雨热同季。土壤以碱性紫色土为主,土层浅薄。20世纪80年代初营建的以柏木为主的人工林由于多种原因,林相残败,树种单一,生物多样性低,生境质量较差,林分产量和生态功能低下,属典型的次生低效人工林。
2材料与方法
2.1样地设置
试验地位于德阳市旌阳区和新镇永新村,于2011年对该区柏木低效纯林进行带状或块状皆伐并引入核桃经济树种,2013年9月初将不同牧草布置于核桃林下。试验设立3个随机区组,区组内土壤肥力等环境条件相对均匀一致,每个区组划分4个10 m×10 m的小区,设置核桃+菊苣、核桃+苜蓿、核桃+鸭茅3种林草及核桃+无植草模式,并以柏木低效林(简称纯柏)为对照,分别设立样地进行观测,样地重复3次。
2.2样品采集及测定
在5种样地内按S形5点混合取样法取0~30 cm土层土样,采样时间为2014年9月。一部分低温保存用于微生物数量测定,分析采用稀释平板法[9],其中细菌采用牛肉膏蛋白胨培养基,真菌采用改良马丁培养基,放线菌采用改良高氏一号培养基;另一部分风干后用于土壤酶活性和养分测定,其中过氧化氢酶活性采用高锰酸钾滴定法,蔗糖酶活性采用3,5-二硝基水杨酸比色法,脲酶活性采用苯酚钠比色法,碱性磷酸酶活性采用磷酸苯二钠比色法[10],土壤有机质采用重铬酸钾氧化-外加热法,土壤全氮采用凯氏法,土壤碱解氮采用碱解扩散法,土壤全磷采用氢氧化钠碱熔-钼锑抗比色法,有效磷采用双酸浸提法,土壤全钾采用氢氧化钠碱熔-火焰光度法,速效钾采用乙酸铵浸提-火焰光度法,pH采用酸度计测定[11]。
2.3土壤肥力评价指标系数标准化
由于各个评价指标具有不同的量纲,因此在采用主成分分析评价土壤肥力之前必须对其原始数据进行标准化处理[12-13]。使得每个变量的平均值为0,标准差为1。
各肥力指标标准化公式为:

(1)

2.4数据处理
采用Microsoft Excel 2007、SPSS 17.0统计软件对土壤微生物、酶活性及养分数据进行基本统计、相关性分析、显著性检验及主成分分析。将主成分分析中各主分量的贡献率作为权重,计算出各模式土壤肥力的综合指标值(IFI)[14],公式如下:
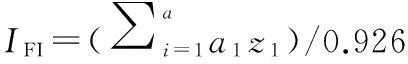
(2)
式中:i表示主分量编号;z表示主成分量;a表示主成分量的方差贡献率。
3结果与分析
3.1不同模式下土壤微生物的组成和数量
由表1可见,各模式之间真菌数量差异不显著。放线菌的数量由高到低依次为核桃+鸭茅、核桃+苜蓿、核桃+菊苣、核桃+无植草、纯柏,其中,核桃+苜蓿、核桃+鸭茅与纯柏之间差异显著(p<0.05),可见这2种林草模式对放线菌的生长与分布具有较大的影响。细菌数量在微生物类群中贡献最大,其变化趋势与微生物总数由高到低依次为核桃+菊苣、核桃+苜蓿、核桃+鸭茅、核桃+无植草、纯柏,即4种改造模式的细菌数量与纯柏之间、核桃+菊苣与核桃+无植草之间差异显著(p<0.05),且3种林草模式与纯柏存在极显著差异(p<0.01)。从土壤微生物总量来看,4种改造模式与纯柏皆为差异极显著(p<0.01)。
三大类群微生物数量均以纯柏最低,每克干土中真菌、放线菌、细菌平均数量仅分别为0.84×104、47.37×104、277.55×104菌落。

表1 不同模式下的土壤微生物量和酶活性
注:表中数据为平均值±标准差。同列数据后不同小写字母表示差异显著(p<0.05),不同大写字母表示差异极显著(p<0.01)。
3.2不同模式下的土壤酶活性
土壤酶是生态系统的物质循环和能量流动等生态过程中最活跃的生物活性物质[15]。采伐柏木后,林地光照、水分条件及耕作、管理方式的改变会引起酶活性发生一系列的转变。而改种核桃并结合牧草栽植后,植物由于自身的生理特性不同,在生长过程中就有相异的代谢功能,必然导致植物向土壤释放酶和对原有土壤酶作用的差异。4种土壤酶活性在不同模式中的表现不同(表1),总体表现为林草模式土壤酶活性高于核桃+无植草和纯柏。其中,核桃+菊苣模式中土壤酶活性最高,过氧化氢酶、蔗糖酶、脲酶、碱性磷酸酶活性分别为126.0、356.0、17.3、19.8 g·g-1·h-1。各模式间过氧化氢酶活性大小无显著性差异。蔗糖酶活性在各模式土壤中的高低顺序为核桃+菊苣、核桃+鸭茅、核桃+苜蓿、纯柏、核桃+无植草,3种林草模式相差不大,但皆显著高于核桃+无植草、纯柏,3种林草模式与核桃+无植草之间、核桃+菊苣、核桃+鸭茅与纯柏之间皆为差异极显著(p<0.01),核桃+苜蓿与纯柏差异显著(p<0.05)。各模式土壤脲酶活性差异不显著,其大小表现跟蔗糖酶一致。土壤碱性磷酸酶表现不同于其他酶,在各模式中的活性高低顺序为核桃+菊苣、核桃+无植草、核桃+苜蓿、核桃+鸭茅、纯柏,其中核桃+菊苣与纯柏差异显著(p<0.05)。
3.3不同模式下的土壤养分
由表2可见,不同土壤养分及pH在各个模式中的分布状况具有差异。各模式林地0~30 cm土层有机质质量分数,除核桃+菊苣与核桃+鸭茅之间差异不显著外,其余模式之间皆差异极显著(p<0.01)。质量分数最高的为纯柏,达38.11 g·kg-1,显著高于其他模式,最小的为核桃+无植草,仅10.78 g·kg-1。此外,种植牧草均显著提高核桃林的土壤有机质质量分数。不同牧草对土壤有机质的影响效果差异不显著,核桃+苜蓿相对核桃+菊苣、核桃+鸭茅较高,三者有机质质量分数分别为19.74、14.63、14.08 g·kg-1。
各模式中全氮、碱解氮质量分数的大小顺序不同,说明土壤含氮量高的林地中真正能被植物吸收利用的那部分氮元素质量分数不一定高,但均以纯柏最高,而且明显高于其他模式,其纯柏的全氮、碱解氮质量分数与4种模式差异极显著(p<0.01)。豆科牧草提高碱解氮的能力具有逐步强于禾本科牧草的趋势,但核桃+菊苣、核桃+鸭茅模式土壤全氮质量分数低于核桃+无植草。全磷、全钾、有效磷和速效钾质量分数大小顺序大致为核桃+菊苣、核桃+鸭茅、核桃+苜蓿、核桃+无植草、纯柏,其中核桃+菊苣与核桃+无植草、纯柏之间的有效磷质量分数差异极显著(p<0.01),核桃+苜蓿与纯柏差异显著(p<0.05),虽然改造后土壤中全磷质量分数变化不明显,但可以直接被植物吸收利用的速效磷质量分数明显增加。改造后促使全钾发生转移,质量分数上升,但全钾释放是一个长期的过程,因此在短时间内变化不明显,而速效钾作为钾素的实时供应指标,其质量分数较改造前有所增加,但差异并不显著。
土壤pH值范围为7.6~8.4,表明土壤为碱性(pH>7.5),土壤pH值最高为纯柏,其次为3种林草复合模式,核桃+无植草略低,即纯柏与核桃+无植草差异显著(p<0.05)。

表2 不同模式下的土壤养分质量分数及pH值
注:表中数据为平均值±标准差。同列数据后不同的小写字母表示差异显著(p<0.05),不同的大写字母表示差异极显著(p<0.01)。
3.4不同模式下土壤微生物、酶活性及养分间的相关性
由表3可知,土壤微生物之间相关性不显著;土壤蔗糖酶与过氧化氢酶、脲酶呈显著正相关,其余酶之间相关性不显著;土壤有机质与全氮、碱解氮及pH之间、全钾与全磷、有效磷之间皆呈极显著正相关关系;真菌与过氧化氢酶、蔗糖酶、脲酶呈显著正相关,放线菌与4种土壤酶相关性皆不显著,细菌与过氧化氢酶显著正相关,与其余3种酶相关性不显著;土壤三大类群菌与有机质、全氮、碱解氮、pH呈负相关。土壤真菌与土壤养分含量的相关性普遍较高,与土壤全磷、全钾达极显著正相关,与土壤速效钾呈显著正相关;过氧化氢酶、蔗糖酶、脲酶与全磷、有效磷、全钾呈极显著或显著正相关,说明该3种酶与土壤磷、钾的转化有一定的关系,而3种酶中只有过氧化氢酶与速效钾存在显著正相关关系,表明过氧化氢酶活性直接影响林地土壤钾素的含量,土壤碱性磷酸酶与全磷、有效磷、全钾、速效钾相关性不显著。4种土壤酶与有机质、全氮、碱解氮、pH皆呈负相关,这可能与林分类型有关,由于研究对象的不同,得出的结论也不同。

表3 土壤微生物、酶活性及养分相关关系
注:*表示显著相关(p<0.05);** 表示极显著相关(p<0.01)。
3.5不同改造模式土壤肥力的评价
3.5.1评价指标体系的建立
根据本研究已测定的指标,选择土壤微生物(真菌、放线菌、细菌)、土壤酶活性(过氧化氢酶、蔗糖酶、脲酶、碱性磷酸酶)、土壤养分质量分数(有机质、全氮、碱解氮、全磷、有效磷、全钾、速效钾)指标数值建立土壤肥力综合评价指标体系。根据公式(1)计算,各肥力指标标准化值见表4。

表4 土壤生物学指标的标准化值
3.5.2土壤肥力指标的主成分分析
由表5可知,经过主成分分析,原来14个土壤肥力因子缩减到2个新的变量(Z1、Z2)。Z1、Z2包含原始数据信息总量达到了92.66%,完全能代表原来14个指标所反映的土壤综合肥力。因此前2个主成分即可反应土壤肥力系统的变异信息。

表5 土壤肥力指标的成分因子得分系数矩阵、特征值及贡献率
由表5可见,第1主成分贡献率达75.81%,主要综合了真菌、放线菌、细菌、过氧化氢酶、蔗糖酶、脲酶、碱性磷酸酶、全磷、有效磷、全钾、有效钾的变量信息,主要反映的是土壤微生物、土壤酶和土壤磷、钾养分。其对土壤肥力评价的作用最大。第2主成分贡献率为16.85%,主要综合了蔗糖酶、脲酶、有机质、全氮、碱解氮的变量信息,主要反映的是土壤酶及有机质、氮养分。这2个综合变量从土壤微生物、土壤酶、部分土壤养分方面影响土壤肥力,用它们来评价土壤肥力具有92.66%的可靠率。X1、X2、X3…X14这14个变量与2个主成分Z1、Z2的关系方程为:
Z1=0.2824X1+0.2612X2+0.2922X3+0.2962X4+
0.2499X5+0.2413X6+0.2103X7-0.2290X8-
0.2649X9-0.2367X10+0.2735X11+0.2867X12+
0.2962X13+0.3014X14。
(3)
Z2=0.179 1X1-0.252 0X2-0.121 8X3+0.115 3X4+
0.360 8X5+0.395 3X6-0.161 5X7+0.409 0X8+
0.325 0X9+0.411 6X10+0.256 6X11+0.162 8X12+
0.156 3X13-0.069 0X14。
(4)
3.5.3土壤肥力综合指标值的计算
将各模式标准化的土壤肥力指标(表4)带入上述表达式(3)、(4),得各模式主成分得分(表5)。利用前面主成分分析得出的2个指标Z1、Z2,以每个主分量Zj的方差贡献率aj作为权重进行加权汇总,用土壤微生物、土壤酶活性及土壤养分含量对低效林不同改造模式土壤肥力进行综合评估,计算结果见表6。5种模式土壤肥力大小顺序依次为核桃+菊苣、核桃+鸭茅、核桃+苜蓿、核桃+无植草、纯柏。从各模式综合评分可知,核桃+菊苣模式得分最高,说明该样地土壤肥力最强;纯柏林地得分最低,说明该样地土壤肥力最差。

表6 各样地主要成分得分及综合评估得分
4结论与讨论
4.1不同模式对土壤微生物的影响
本研究结果显示,不同模式间土壤真菌数量差异不显著,可能与该菌对土壤pH的要求比放线菌和细菌所要求的低有关。陶勇等[16]在江汉平原杨树林下土壤微生物的年动态变化研究中得出,碱性林地土壤中放线菌和细菌数量皆显著大于酸性林地土壤。因为真菌适合生长在酸性土壤中,而各个模式样地皆为碱性土壤,造成该类群菌活性相应减弱,则数量在各个模式中的差异不明显。核桃+苜蓿和核桃+鸭茅模式能显著提高土壤放线菌数量,因该2种牧草根系,特别是须根较发达,能向土壤中提供大量死亡根和根系分泌物等残余物,丰富了该土壤微生物的营养源。同时,温度、湿度、通透性及养分等的土壤环境的改变都会引起土壤放线菌数量发生变化。4种改造模式均能提高土壤细菌数量,可能跟该微生物类群菌本身特性及其所生活的微环境有关。
三大类群微生物数量均以纯柏最低,主要归因于柏木林地土壤黏重板结。改造后,土质相对疏松,土壤通透性增大,且林地光照和水热条件相对较好,顺应了土壤微生物的生长。
总体而言,从微生物数量上看,3种林草复合模式较核桃+无植草改造效果更优,不同草种其特性不同,对土壤发挥的作用也各异,但三者对土壤肥力的影响差异不明显,可能是栽培时间较短,彼此间的差异性暂未表现。
4.2不同模式对土壤酶活性的影响
各个模式间的土壤过氧化氢酶、脲酶活性大小差异皆不显著,说明核桃林地种植牧草后土壤解除呼吸过程中产生的过氧化氢的能力未得到加强。3种林草模式中的蔗糖酶活性相差不大,但皆显著高于核桃+无植草、纯柏,主要是植被覆盖度相应提高,土壤中的植物根系尤其是细根比较丰富,其分泌物和脱落细胞促使酶进入土壤。核桃林地种植菊苣能显著提高土壤碱性磷酸酶,这与菊苣自身的根系分泌物有关,不同植物由于根系分泌物差异可以引起土壤磷酸酶活性的变化[17-18]。
4.3不同模式对土壤养分的影响
采伐后林地土壤水热状况的改变能引起土壤磷、钾养分含量的升高,说明林下植物具有促进营养元素在地表富集的作用。且生草能够通过减少地表径流量来降低土壤磷、钾元素的流失。与孙霞等[19]和林兴生等[20]生草能提高有效磷、钾含量的报道一致。但是土壤有机质和氮含量反而相对降低,由于已种植多年的柏木林分郁闭度高,相应的枯枝落叶层较厚,分解周期较长,土壤腐殖质聚积,而采伐柏木改种核桃后,生长初期的核桃林分暂未郁闭,植物残体等有机物稀少,也可能因为核桃正处于生长初期,自身需要吸收大量的营养成分,造成土壤原本含有的有机质、氮含量逐步降低。此外,种植牧草的核桃林地土壤有机质及氮含量相对无植草核桃林地高,说明在核桃林地种植牧草(特别是豆科牧草)具有固氮功能,且对土壤有机质具有累积作用。相对而言,光照、水分条件好的未郁闭核桃林比较适合土壤微生物的生长及促进酶活性的提高,而生长周期更长,林下覆盖物较多的柏木林,其土壤有机质及氮含量更丰富。此外,土壤pH也相应地降低,可能因为植物根系分泌物中的有机酸在一定的程度上降低了土壤pH,也可通过对一些阳离子的络合减少其对磷酸盐的固定,从而提高有效磷的含量[21]。从长远来看,一般针叶树种的落叶量较小,针叶所含灰分也较缺乏,并且分解比较困难[22],而阔叶树种往往能发挥相反的作用,因此替换阔叶树种可能更能维持和提高地力。
4.4土壤微生物、酶活性及养分相关性
对5种模式的土壤微生物、酶活性及养分指标间的相关性分析发现,土壤微生物与酶活性具有一定的相关性,一般认为土壤酶主要来源于土壤微生物代谢过程及植物根系分泌物,而土壤微生物是土壤生态系统中土壤酶的重要来源。因此,在一定的条件下,土壤酶活性会随着微生物数量的变化而发生一定的变化,并且两者与土壤磷、钾养分也具有一定的相关性,说明微生物数量及酶活性的增加,有助于提高土壤磷、钾的供应能力。诸多研究结果表明,土壤微生物、酶活性与养分含量相关性明显,土壤酶活性可以作为土壤养分评价的一个重要指标[23-24],但本研究得出土壤微生物数量、酶活性与土壤有机质、氮含量及pH呈负相关。不同的研究者就同一种土壤微生物、同一种土壤酶与同一种土壤养分的相关性得出不同的结论[8,25-27],彼此间的关系未达成共识,且由于研究对象的不同,得出三者之间相关性的结论也不一致。总而言之,本试验结果表明三者之间的相关程度不同,造成该种关系可能与研究对象及土质状况、气候条件或者各指标本身的性质有关。与前人研究的不一致性也体现了土壤微生物、酶及养分之间关系的复杂性,说明它们三者之间的关系需要进一步研究。
4.5不同模式土壤肥力评估
通过土壤肥力综合指数的分析,各个改造模式均能有效改善土壤质量,并且改善土壤状况效果有所不同,其中,核桃+菊苣模式改造效果最佳,反应了各种牧草对土壤肥力的影响不同。董智等[28]在黄泛平原风沙化土地种植牧草改良土壤效果研究中也认为不同牧草改良土壤的效果不同。本研究采用林草结合,一方面,能够避免杂草的生长及核桃林下土地未被利用而闲置、造成土地资源浪费;另一方面,林草复合模式在解决农林“争地”矛盾的同时,还能有效地控制土壤侵蚀,减少养分亏损。而且在营造人工牧草的过程中,理应施入适量的有机肥、适当将部分牧草归还林地,使得草体中的氮、磷、钾等元素被释放于土壤中,并与土壤黏粒结合,以增加土壤养分的人为输入。同时,种植牧草能增加土壤水分,避免果树根系生长受高温季节的影响。曾艳琼等[29]通过研究认为林草复合系统对于改善土壤理化性质,提高土壤水分含量和土壤肥力具有良好的效果。姚月锋等[30]研究得出果草套种模式对南方丘陵山地表层土壤肥力条件的改善作用较为明显。林地生草是一种天然绿肥,日本、韩国及美国等国家都大力发展林地绿肥产业[31-32]。本研究还需进行长期的观察研究,以掌握林草复合系统中土壤肥力的变化规律,为林草复合的最适栽培模式提供一定的理论依据。
参考文献
[1]杨玉波,李承彪.四川森林[M].北京:中国林业出版社,1992.
[2]杨玉坡.长江上游(川江)防护林研究[M].北京:科学出版社,1993.
[3]冯文贵,刘洋.四川盆地低山丘陵区柏木低效林改造效应研究[J].安徽农业科学,2008,36(15):6296-6297,6303.
[4]徐东,胡玉芳,刘光兰.柏木长防低效林的形成原因及改造措施探讨[J].重庆林业科技,2007(1):35-36.
[5]骆宗诗,侯波,向成华,等.四川盆地低山丘陵区柏木低效防护林的改造[J].中南林业科技大学学报,2009,29(6):82-87.
[6]黎燕琼,龚固堂,郑绍伟,等.低效柏木纯林不同改造措施对水土保持功能的影响[J].生态学报,2013,33(3):934-943.
[7]ROBERTSM R. Response of the herbaceous layer to natural disturbance in North American forests[J]. Canadian Journal of Botany,2004,82(9):1273-1283.
[8]杨涛,徐慧,李慧,等.樟子松人工林土壤养分、微生物及酶活性的研究[J].水土保持学报,2005,19(3):50-53.
[9]李振高,骆永明,滕应.土壤与环境微生物研究法[M].北京:科学出版社,2008.
[10]关松荫.土壤酶及其研究法[M].北京:中国农业出版社,1986:274-339.
[11]中国科学院南京土壤研究所.土壤理化分析[M].上海:上海科学技术出版社,1978:62-153.
[12]骆伯胜,钟继洪,陈俊坚.土壤肥力数值化综合评价研究[J].土壤,2004,36(1):104-106.
[13]庞学勇.川西亚高山针叶林不同演替阶段土壤特性比较研究[D].雅安:四川农业大学,2002.
[14]张猛,张健.林地土壤微生物、酶活性研究进展[J].四川农业大学学报,2003,21(4):347-351.
[15]BADIANE N N Y, CHOTTE J L, PATE E, et al. Use of soil enzyme activities to monitor soil quality in natural and improved fallows insemi-arid tropical regions[J]. Applied Soil Ecology,2001,18(3):229-238.
[16]陶勇,陈防,万开元,等.江汉平原杨树林下土壤酶及微生物的年动态变化[J].福建林业科技,2009,36(4):90-97.
[17]MARSCHNER P, SOLAIMAN Z, RENGEL Z. Rhizosphere properties of poaceae genotypes under p-limiting conditions[J]. Plant and Soil,2006,283(1):11-24.
[18]韩东苗,张林成,冯茂松,等.柏木低效林不同改造模式中土壤微生物及酶活性的研究[J].华南农业大学学报,2016,37(1):96-103.
[19]孙霞,柴仲平,蒋平安,等.生草对新疆南部干旱区苹果园土壤肥力的影响[J].中国农学通报,2011,27(7):177-181.
[20]林兴生,林占熺,林冬梅,等.荒坡地种植巨菌草对土壤微生物群落功能多样性及土壤肥力的影响[J].生态学报,2014,34(15):4304-4312.
[21]赵小亮,刘新虎,贺江舟,等.棉花根系分泌物对土壤速效养分和酶活性及微生物数量的影响[J].西北植物学报,2009,29(7):1426-1431.
[22]毕英杰.对营造混交林优越性的研究[J].林业勘查设计,2004(2):36-37.
[23]宋海燕,李传荣,许景伟,等.滨海盐碱地枣园土壤酶活性与土壤养分、微生物的关系[J].林业科学,2007,43(增刊1):28-32.
[24]郝艳,刘君昂,周国英,等.不同抚育措施对油茶林土壤养分、微生物及酶活性的影响[J].林业资源管理,2008(6):97-101.
[25]齐文娟.黄河源区高寒草地土壤微生物数量动态及其与土壤养分关系研究[D].兰州:甘肃农业大学,2007.
[26]李春艳,李传荣,许景伟,等.泥质海岸防护林土壤微生物、酶与土壤养分的研究[J].水土保持学报,2007,21(1):156-159,200.
[27]蒋智林,刘万学,万方浩,等.紫茎泽兰与非洲狗尾草单、混种群落土壤酶活性和土壤养分的比较[J].植物生态学报,2008,32(4):900-907.
[28]董智,李红丽,任国勇,等.黄泛平原风沙化土地种植牧草改良土壤效果研究[J].中国草地学报,2008,30(3):84-87.
[29]曾艳琼.花椒林-牧草间作对牧草生长、光合特性及土壤理化性质的影响[D].北京:北京林业大学,2008.
[30]姚月锋,徐广平,黄玉清,等.果草套种对南方丘陵山地表层土壤微生物量碳氮及酶活性的影响[J].水土保持通报,2015,35(6):32-37.
[31]ASAGI N, UENO H. Nitrogen dynamics in paddy soil applied with various15 N-labdlled green manures[J]. Plant and Soil,2009,322(1):251-262.
[32]KIM D J, CHUNG D S, BAI S C C, et al. Effects of soil selenium supplementation level on selenium contents of green tea leaves and milk vetch[J]. Journal of Food Science and Nutrition,2007,12(1):35-39.
收稿日期:2015年11月21日。
第一作者简介:韩东苗,女,1991年1月生,四川农业大学林学院,硕士研究生。E-mail:32686632@qq.com。通信作者:冯茂松,四川农业大学林学院,副教授。E-mail:705592631@qq.com。
1)国家科技支撑计划项目(2011BAC09B05);四川省高等学校重点实验室建设项目。
责任编辑:程红。
