干旱胁迫对不同抗旱性小麦种子萌发及幼苗根芽生物量分配的影响
2016-05-27施成晓王昌江秦晓梁廖允成
施成晓,陈 婷,王昌江,秦晓梁,廖允成
(西北农林科技大学农学院,陕西杨陵 712100)
干旱胁迫对不同抗旱性小麦种子萌发及幼苗根芽生物量分配的影响
施成晓,陈 婷,王昌江,秦晓梁,廖允成
(西北农林科技大学农学院,陕西杨陵 712100)
摘要:为探讨冬小麦种子萌发期抗旱性与幼苗生物量分配的关系,以小麦抗旱型品种长旱58和干旱敏感型品种漯麦18为材料,采用聚乙二醇(PEG-6000)模拟干旱胁迫的方法,测定和分析了干旱胁迫后小麦种子发芽率、幼苗根冠比、叶绿素含量等16个指标的变化。结果表明,干旱胁迫后,长旱58种子发芽率无明显变化,漯麦18种子发芽率显著下降。干旱胁迫下长旱58种子发芽势、发芽指数、活力指数和干物质转移量显著大于漯麦18,平均发芽时间显著小于漯麦18,长旱58幼苗根、芽的长度和干重及总干重均显著高于漯麦18。干旱胁迫显著增加漯麦18幼苗根冠比,对长旱58萌发第五和第七天的幼苗根冠比无显著影响。干旱胁迫显著降低漯麦18幼苗叶绿素含量,对长旱58无显著影响。由于小麦萌发期幼苗干物质累积取决于种子中营养物质的转移和呼吸消耗,因此推测种子营养物质的快速转移和低呼吸消耗有助于提高小麦萌发期的抗旱能力,促进干旱胁迫下种子萌发和幼苗生长。
关键词:小麦;干旱胁迫;种子萌发;生物量分配;根冠比;干物质转移
小麦作为我国第三大粮食作物,其产量的高低和稳定性直接影响粮食安全,决定着国计民生[1]。随着全球性气候变化和生态破坏的加剧,水资源短缺已成为阻碍农作物正常生长发育的主要因素之一。种子萌发和幼苗生长是小麦生育期中最易受到逆境胁迫影响的两个生长阶段[2]。逆境条件下小麦种子发芽率高低、幼苗生长状况好坏会影响中后期植株生长发育[3]及群体建成和质量[4-5]。在土壤墒情不足条件下,小麦种子萌发及出苗会受到严重抑制,导致田间出现缺苗断垄,直接影响基本苗数,造成群体数量下降,不利于小麦高产。
小麦根系性状与植株抗旱性存在密切关系,干旱胁迫下小麦根系发育迟缓,次生根数量减少[6]。干旱逆境下小麦抗旱品种根系发达,根生物量和长度大,根冠比高[7]。干旱胁迫会影响小麦种子发芽率和发芽速率,其中对发芽速率的影响更突出[8]。在种子萌发到幼苗自身合成糖类物质之前,幼苗正常生长主要依赖于种子贮藏的营养物质供应,同时萌发的种子和幼苗通过呼吸作用会消耗部分能量,导致幼苗生物量低于种子干物质量,因此种子干物质的转移量和转移效率会影响幼苗根和芽的生长状况[9-10]。然而,关于干旱胁迫对小麦种子萌发影响的报道大多集中于不同品种萌发特性或幼苗生长[4-10],很少涉及干旱胁迫下小麦幼苗根芽生物量分配特点。在我国西北地区,干旱频繁发生,小麦播种期常会出现土壤水分不足,导致小麦播种后苗少、苗弱[11]。本实验在已有研究[1-11,12-23]的基础上,选取抗旱品种长旱58和干旱敏感型品种漯麦18为材料,分析了干旱胁迫下小麦种子萌发特性、幼苗生物学特征和种子干物质转移状况,以进一步探讨冬小麦种子萌发期抗旱性与幼苗生物量分配的关系,以期为抗旱育种提供一定的理论依据。
1材料与方法
1.1实验材料
供试材料为抗旱型品种长旱58(国审麦2004015)和干旱敏感型品种漯麦18(国审麦2012011)。
1.2实验设计
1.2.1发芽实验
实验采用培养皿滤纸上发芽的方法,培养皿直径为9 cm,设置15%-PEG6000溶液模拟干旱胁迫和蒸馏水(对照)2个处理。每个品种挑选籽粒均匀饱满的种子,用10%次氯酸钠消毒10 min后,蒸馏水清洗3遍,并用洁净的滤纸将水分吸干后,在铺有2层滤纸的培养皿中摆放50粒种子,每个品种各处理设置3个重复,每天每个培养皿中用注射器注入5 mL PEG溶液或蒸馏水,置于光照培养箱中进行萌发,温度设置为25 ℃,昼夜长设置为16 h/8 h。每24 h记录一次发芽情况(当芽长达到种子长度一半时视为发芽),第3天计算发芽势,第7天分别计算发芽率、发芽指数、活力指数及平均发芽时间:
发芽势= (第3天时发芽数/供试种子数) × 100%;
发芽率= (第7天时全部正常发芽的种子数/供试种子数) × 100%;
发芽指数=∑Gt/Dt,∑Gt为t天内种子萌发总数,Dt为相对应的种子发芽天数;
活力指数=发芽指数×L,L为种子根长和芽长之和;
平均萌发时间=∑(Gi×i)/∑Gi,i代表从萌发试验开始后的萌发天数,Gi代表第i天的萌发个数。
1.2.2幼苗生长实验
实验采用的培养皿直径为16 cm,设置的处理同发芽实验。每个小麦品种挑选籽粒均匀饱满的种子后,每个品种取3份200粒种子置于80 ℃烘箱中24 h,称量种子干重;其余种子用10%次氯酸钠消毒10 min后,蒸馏水清洗3遍,用洁净的滤纸将水分吸干,在铺有2层滤纸的培养皿中摆放100粒种子,每个品种每个处理设置10个重复,每天每个培养皿中用注射器注入10 mL PEG溶液或蒸馏水。置于光照培养箱中进行萌发,温度设置为25 ℃,昼夜长设置为16 h/8 h。萌发3 d、5 d和7 d时,每个品种每个处理取10株幼苗,用剪刀剪下根、芽,测量根长和芽长后,置于80℃烘箱内烘干至恒重,称量芽、根和种子干重。根据马守才[24]和Soltani[25]的方法计算根冠比、干物质转移量、干物质转移率、干物质转化效率和呼吸消耗干物质量。
根冠比= 根干重/芽干重;
干物质转运量= 发芽前种子干重-发芽后种子残留物干重;
干物质转移率= (种子贮藏干物质转移量/发芽前种子干重) × 100%;
干物质转化效率= (幼苗干重/种子贮藏干物质转移量) × 100%;
呼吸消耗量= 干物质转移量-幼苗干重。
1.3叶绿素含量和可溶性总糖含量的测定
采用蒽酮比色法测定可溶性总糖含量,采用80%丙酮比色法测定叶绿素含量[26]。
1.4数据处理
用Microsoft Excel 2010对数据进行整理, 用SPSS 22.0统计软件进行方差分析,以Sigmaplot 12.5作图。
2结果与分析
2.115%-PEG胁迫下不同抗旱性小麦品种的萌发特性
在对照条件下,两个小麦品种种子发芽率没有显著差异;干旱胁迫对长旱58种子发芽率无明显影响,但显著降低了漯麦18种子发芽率(图1)。在对照和干旱胁迫条件下,长旱58的发芽势、发芽指数和活力指数均显著大于漯麦18,平均发芽时间显著小于漯麦18。干旱胁迫显著降低了两个品种种子发芽势、发芽指数和活力指数,同时显著延长发芽时间(表1)。
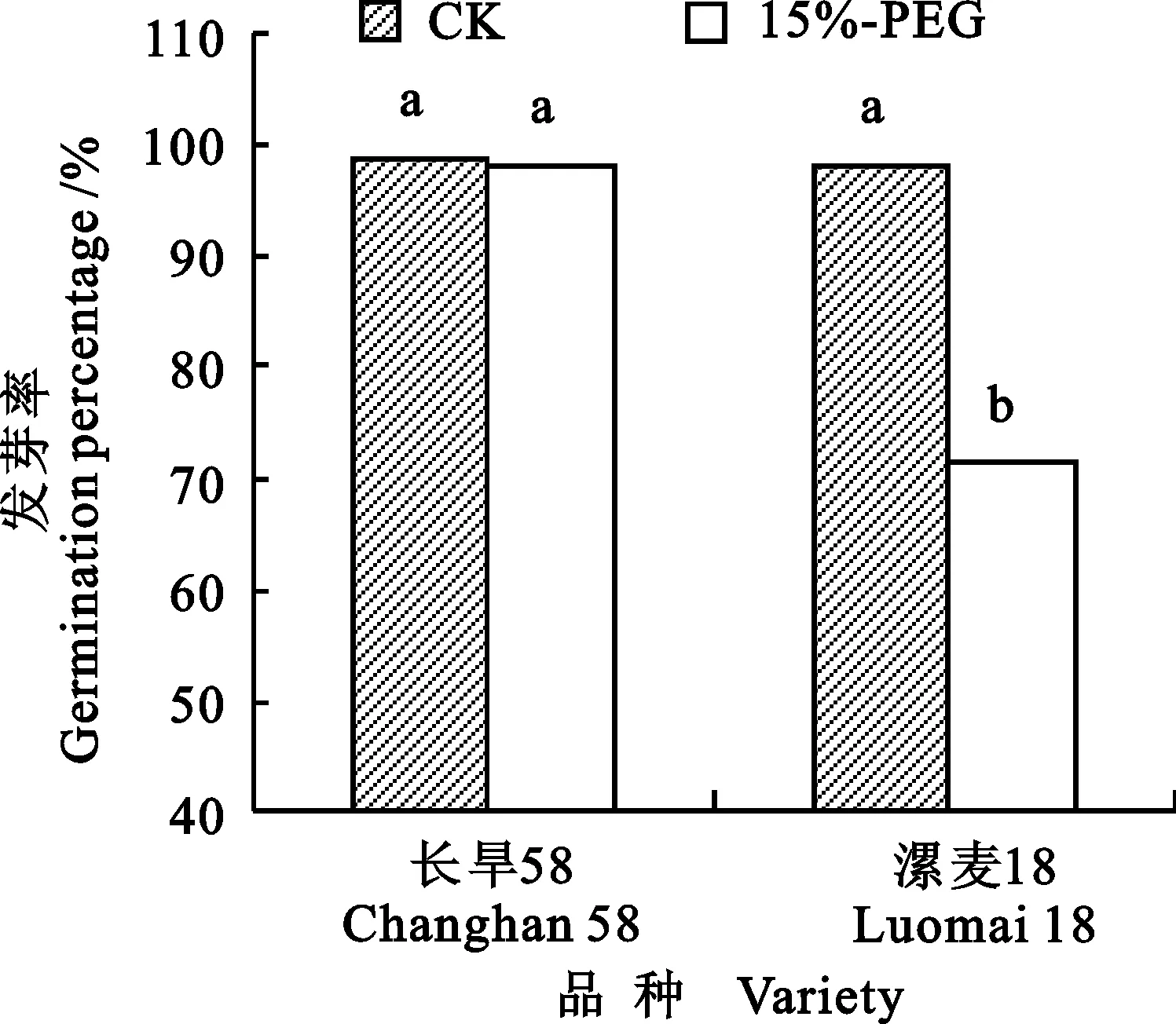
图柱上不同字母表示处理间差异显著(P<0.05)。图3和图4同
The different letters mean significant difference among the different treatments at 0.05 level. The same as in Fig.3 and Fig.4

图1 对照和干旱胁迫下小麦的种子发芽率
表中小写字母表示相同处理下不同品种间差异显著(P<0.05),大写字母表示同一品种不同处理间差异显著(P<0.05)。下表同
The lower-case letters mean significant difference between different varieties under a same treatment (P<0.05);the capital letters mean significant difference between different treatments for a same variety (P<0.05).The same as in other tables
2.2干旱胁迫下不同抗旱性小麦品种幼苗生长状况的比较
干旱胁迫下两个小麦品种幼苗根长、芽长、根干重和芽干重在整个萌发过程中均显著小于对照;除萌发3 d时品种间芽干重没有差异外,长旱58幼苗根长、芽长、根干重和芽干重均显著大于漯麦18。此外,干旱胁迫对芽长、芽生物量的抑制作用大于根系(表2)。以上结果表明,长旱58幼苗生长比漯麦18易于适应干旱环境。
无论在对照还是在干旱胁迫条件下,随着萌发时间的延长,两个小麦品种幼苗根冠比均呈减小趋势。与对照相比,干旱胁迫使根冠比增大,其中长旱58在萌发3 d时根冠比增加显著,而漯麦18整个萌发过程根冠比增加均达到显著水平(图2)。以上结果表明,小麦种子萌发时分配到芽的生物量多于分配到根的生物量;干旱胁迫下,长旱58幼苗生物量的分配比漯麦18更合理,因此更有利于根系与茎叶协调生长,以适应干旱胁迫。

表2 对照和干旱胁迫下小麦幼苗的根长、芽长、根干重和芽干重
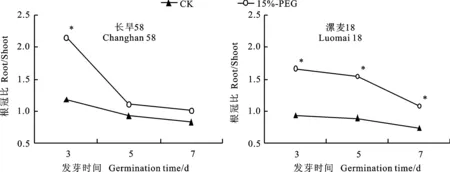
*表示不同处理间差异显著(P<0.05)
* means significant difference between two treatments at 0.05 level
图2对照和干旱胁迫下小麦幼苗的根冠比
Fig.2Ratio of root to shoot of wheat under the treatments of CK and 15% PEG
2.3干旱胁迫下不同抗旱性小麦品种种子干物质转移的差异
干旱胁迫导致两个小麦品种幼苗生物量均显著降低,其中漯麦18的降幅显著大于长旱58(表3)。在对照条件下,两个小麦品种种子干物质转移量差异在萌发3 d和5 d时达到显著水平;干旱胁迫显著减少种子干物质的转移量,其中长旱58减少量显著低于漯麦18。在对照条件下, 萌发5 d时长旱58种子干物质转移率显著大于漯麦18,但在萌发7 d时显著低于漯麦18;干旱胁迫使两个品种种子干物质转移率显著减小,其中萌发3 d和5 d时长旱58的减少量显著低于漯麦18。干旱胁迫显著降低了长旱58在萌发3 d时种子干物质转化效率,对漯麦18种子干物质转化效率没有显著影响。在对照条件下,长旱58呼吸消耗的干物质量在萌发5 d时显著大于漯麦18;干旱胁迫在萌发5 d和7 d时均能显著减少呼吸消耗的干物质。以上结果表明,干旱胁迫下冬小麦种苗转化过程中幼苗生物量降低的主要原因是种子干物质转移率降低,同时降低了干物质的转化效率。长旱58在干旱胁迫下幼苗干物质积累和种子干物质转化均显著大于漯麦18,因而更有利于幼苗生长发育和适应干旱环境。
2.4干旱胁迫对小麦种子萌发期间叶绿素和可溶性糖含量的影响
2.4.1对叶绿素含量的影响
无论在对照还是干旱胁迫条件下,两个小麦品种幼苗叶绿素含量均随时间逐渐增加。在干旱胁迫条件下,漯麦18幼苗叶绿素含量在整个萌发生长过程中均显著低于对照,而长旱58幼苗叶绿素含量与对照无显著差异(图3)。说明干旱条件下抗旱品种长旱58可以维持较高的叶绿素含量,避免光合作用受到明显抑制,从而减轻干旱对幼苗物质生产的影响。

表3 对照和干旱胁迫下小麦种子贮藏物质转运的比较
SW:幼苗干重;DA:干物质转移量;DR:干物质转移率;DE:干物质转化效率;BD:呼吸消耗干物质量
SW:Dry weight of seedling;DA:Transferred amount of dry matter;DR:Transferred rate of dry matter;DE:Transferred efficiency of dry matter;BD:Breathing consumption of dry matter
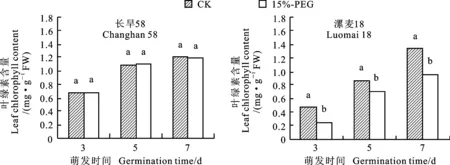
图3 对照和干旱胁迫下小麦幼苗的叶绿素含量
2.4.2对可溶性糖含量的影响
无论对照还是干旱胁迫条件下,两个小麦品种幼苗可溶性糖含量随萌发时间的延长而逐渐减少,萌发3 d时干旱胁迫下芽可溶性糖含量显著高于对照,萌发5 d和7 d时干旱胁迫降低了芽可溶性糖含量,其中漯麦18减少的幅度(31.2%和47.8%)大于长旱58(4.5%和9.3%)。在对照和干旱胁迫条件下,两个小麦品种幼苗根系可溶性糖含量均随萌发时间的延长呈先升后降的趋势,干旱胁迫降低了小麦幼苗根系可溶性糖含量(图4),其中在萌发5和7 d时,漯麦18的降幅(62.5%和60.5%)显著大于长旱58(14.7%和14.4%)。以上结果说明,干旱胁迫对长旱58幼苗可溶性糖含量的影响小于漯麦18。
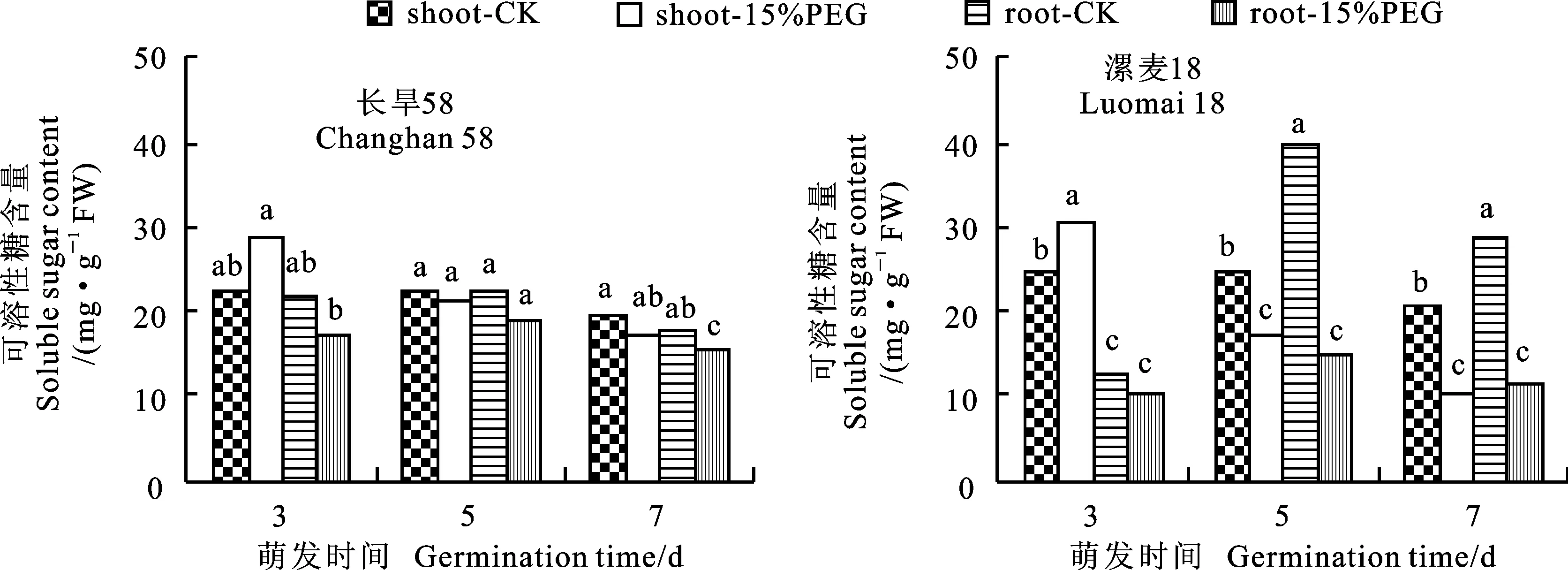
图4 对照和干旱胁迫下小麦幼苗的可溶性糖含量
3讨 论
种子萌发和幼苗生长是小麦生长发育的开始,其发芽率高低、幼苗生长状况好坏、萌发生理过程等均会决定后续的生育状况[3],同时影响小麦群体的建成[4-5]。作物群体密度对产量和品质具有重要影响,播量和发芽率共同决定密度,研究不同抗旱性品种的发芽率对旱区作物的适宜播量确定至关重要。小麦种子的发芽率、发芽势、发芽时间、发芽指数、活力指数等萌发指标可以反映小麦种子萌发期抗旱性的强弱[12],因此可以通过比较不同品种在干旱胁迫和对照间的萌发指标来明晰不同品种的抗旱性差异。本研究结果表明,15%-PEG胁迫显著降低漯麦18种子发芽率、发芽势、发芽指数和活力指数,同时显著抑制发芽速度,该结果与于军等[13]对矮沙冬青种子萌发、原小燕等[14]对不同抗旱性油菜种子萌发和王宏征等[15]对不同倍性小麦种子萌发对干旱胁迫响应的研究结果一致。
小麦幼苗生长过程中地上部和地下部生物量分配比例是受自身遗传因素和生长环境因素耦合作用而表现出的综合结果[17]。当植株地上部和地下部生长均匀、根冠比协调时,对资源的利用效率将达到最大化[18]。本研究表明,干旱胁迫显著降低小麦幼苗根长、芽长、根生物量和芽生物量,干旱敏感型小麦品种漯麦18受到的影响显著大于抗旱型品种长旱58。此外,干旱胁迫显著增加了漯麦18幼苗根冠比,表明干旱胁迫对漯麦18幼苗地上部生长的抑制比根系强,导致幼苗生物量优先分配到根系,芽获得生物量显著降低,使幼苗生长受到显著抑制;而长旱58幼苗根冠比只在萌发第三天显著增加,第五天和第七天与对照无显著区别,使长旱58幼苗在干旱胁迫下维持较高的地上部生物量和较佳的地上部和地下部生长协调性。
Bewley等[9]和Voigt等[10]指出小麦种苗转化过程所需的能量与营养是由种子中贮藏的有机物质提供,但因呼吸作用的存在,幼苗生物量的积累总是少于种子干物质转移量,因此种子干物质转移量和转移效率在一定程度上可能决定了幼苗生长状况。本研究结果表明,在15%-PEG胁迫下,小麦幼苗干物质积累量、种子干物质转移量、转移率和转化效率显著降低,其中干旱敏感型品种漯麦18受到的影响显著大于抗旱型品种长旱58。说明在种苗转化期间,干旱胁迫抑制幼苗物质积累的可能原因是抑制了种子干物质转移率和转化效率,因此在抗旱育种过程中提高种子干物质转移率将有利于种子在干旱环境中萌发建苗。本研究结果还显示,干旱胁迫下,抗旱型品种萌发过程中呼吸消耗的干物质量和幼苗生物量大于敏感型品种,这表明在干旱环境中种子萌发呼吸消耗干物质量的多少可能影响着该品种幼苗生物量的积累量,因此可以作为鉴定抗旱性的指标,但仍需进一步研究。
干旱胁迫会降低植物叶片叶绿素含量,叶片失水会影响叶绿素合成,同时会导致已合成的叶绿素快速分解,造成叶片变黄。近年来,叶绿素荧光与植物抗逆性之间的关系成为了逆境研究的热点之一[19]。张永强等[20]研究指出,中度以上的干旱胁迫会明显抑制植物光合作用,导致光合作用受抑制的主要原因是干旱胁迫阻碍了叶绿体的功能;杨晓青等[21]发现干旱条件下冬小麦抗旱性与叶绿素荧光参数变化有显著关系。本研究表明,在15%-PEG胁迫下,干旱敏感型品种漯麦18叶绿素含量显著降低,而抗旱型品种长旱58叶绿素含量不受影响,这与杨晓青等[21]的研究结果一致,说明干旱条件下抗旱型品种可以维持较高的叶绿素含量,可以避免光合作用受到明显抑制,从而减轻干旱危害。可溶性糖含量的变化可以体现植物对干旱胁迫的适应性反应。孙 萍等[22]发现可溶性糖的积累可以通过渗透调节来减轻干旱胁迫对长春花的伤害;王 川等[23]指出在干旱胁迫下,抗旱小麦植株中可溶性糖积累量比非抗旱品种小麦多。本研究结果显示,长旱58在干旱胁迫下根和芽的可溶性糖含量与对照没有显著区别,在萌发的第五天和第七天漯麦18在干旱胁迫下根和芽可溶性糖含量显著低于对照,这与前人研究结果一致,说明可溶性糖含量是可以作为衡量植物抗旱性强弱的指标。
本研究中,干旱胁迫影响小麦种子萌发的主要原因在于前期种子贮藏物质的转移受到抑制,后期幼苗叶绿素合成和可溶性糖含量的积累受到抑制。不同抗旱性小麦植株可溶性糖含量与叶绿素含量不同,在干旱条件下,叶绿素含量影响着植株光合作用,从而影响着可溶性糖含量的变化及分配,但其相互调节的作用机制仍需进一步研究。此外,与漯麦18相比,干旱胁迫下长旱58抗旱优势具体体现在萌发期间具有较高的发芽势、发芽指数、活力指数,生长过程中有较长的根长和芽长、较高根芽生物量、较大的根冠比和较稳定的叶绿素、可溶性糖含量。漯麦18和长旱58可以作为理想的模式品种用于小麦抗旱育种研究,但由于小麦抗旱性与整个生育期生理、形态、分子等多项指标有关,故仍需进行更多的深入研究。
参考文献:
[1]中华人民共和国农业部.中国农业统计资料(2012) [M].中国农业出版社,2013.
The Ministry of Agriculture of the People’s Republic of China.China’s Agricultural Statistical Data(2012) [M].China Agriculture Press,2013.
[2]Omami E N,Haigh A M,Med R W,etal.Changes in germination ability,dormancy and viability ofAmaranthusretroflexusas affected by depth and duration of burial [J].WeedResearch,1999,39:345-354.
[3]蔺吉祥,李晓宇,唐佳红,等.盐碱胁迫对小麦种子萌发、早期幼苗生长及Na+、K+代谢的影响[J].麦类作物学报,2011,31(6):1148-1152.
Lin J X,Li X Y,Tang J H,etal.Effects of salt and alkali stresses on seed germination,early seedling growth and the metabolize of Na+and K+in shoots of wheat [J].JournalofTriticeaeCrops,2011,31(6):1148-1152.
[4]Deng X P,Shan L,Inanaga S.Sensitivity and resistance of seedling establishment to water stress in spring wheat [J].CerealResearchCommunications,2002,30:125-132.
[5]Okcu G,Kaya M D,Atak M.Effects of salt and drought stresses on germination and seedling growth of pea(PisumsativumL.) [J].TurkishJournalofAgricultureandForestry,2005,29:237-242.
[6]Ekanayake I,Garrity D,etal.Inheritance of root characters and their relations to drought resistance in rice [J].CropScience,1985,25(6):927-933.
[7]周桂莲,杨慧霞.小麦抗旱性鉴定的生理生化指标及其分析评价[J].干旱地区农业研究,1996,14(2):65-71.
Zhou G L,Yang H X.Analysis and review of physiological and biochemical indexes about wheat drought resistance identification [J].AgriculturalResearchintheAridAreas,1996,14(2):65-71.
[8]刘世鹏,叶 飞,曹娟云,等.水分胁迫对红小豆和绿豆发芽的影响[J].北方园艺,2011(15):38-41.
Liu S P,Ye F,Cao J Y,etal.Effects of water stress on germination of adzuki bean and mung bean [J].NorthernHorticulture,2011(15):38-41.
[9]Bewley J D.Seed germination and dormancy [J].Plant Cell,1997,9:1055-1066.
[10]Voigt E L,Almeida T D,Chagas R M,etal.Source-sink regulation of cotyledonary reserve mobilization during cashew(Anacardiumoccidentale)seedling establishment under NaCl salinity [J].PlantPhysioogyl,2009,166:80-89.
[11]山 仑,邓西平,苏 佩,等.挖掘作物抗旱节水潜力-作物对多变低水环境的适应与调节[J].中国农业科技导报,2000,2(2):66-70.
Shan L,Deng X P,Su P,etal.Exploitation of crop drought resistance and water-saving potentials-adaptability of the crops to the low and variable water conditions [J].ReviewofChinaAgriculturalScienceandTechnology,2000,2(2):66-70.
[12]Kumar A,Bahadur B,Sharma B K.Influence of drought on the germination and seedling growth ofHordeumvulgareL.[J].AnnalAridZone,1988,27:65-66.
[13]于 军,焦培培.聚乙二醇(PEG-6000)模拟干旱胁迫抑制矮沙冬青种子的萌发[J].基因组学与应用生物学,2010,29(2):355-360.
Yu J,Jiao P P.Inhibition of seed germination ofAmmmopiptanthusnanusCheng f.under simulated drought stress with polythylene glycol(PEG-6000) [J].GenomicsandAppliedBiology,2010,29(2):355-360.
[14]原小燕,符明联,何晓莹.不同抗旱性油菜种子萌发期抗旱指标比较研究[J].干旱地区农业研究,2012,30(5):77-81.
Yuan X Y,Fu M L,He X Y.The comparative study on drought resistance index of rape with different drought resistance in germination [J].AgriculturalResearchintheAridAreas,2012,35(5):77-81.
[15]王征宏,赵 威,郭秀璞,等.不同倍性小麦种子萌发特征及其对水分胁迫敏感性的差异[J].植物生理学报,2013,49(8):817-823.
Wang Z H,Zhao W,Guo X P,etal.Seed germination characteristics and their differences in sensitivity to water stress in wheat with different ploidy levels [J].PlantPhysiologalJournal,2013,49(8):817-823.
[16]张盼盼,冯佰利,王鹏科,等.干旱条件下糜子叶片衰老与保护酶活性变化[J].干旱地区农业研究,2010,28(2):99-103.
Zhang P P,Feng B L,Wang P K,etal.Leaf senescence and protective enzyme of broom corm millet under drought condition [J].AgriculturalResearchintheAridAreas,2010,28(2):99-103.
[17]马富举,李丹丹,蔡 剑,等.干旱胁迫对小麦幼苗根系生长和叶片光合作用的影响[J].应用生态学报,2012,23(3):724-730.
Ma F J,Li D D,Cai J,etal.Responses of wheat seedlings root growth and leaf photosynthesis to drought stress [J].ChineseJournalofAppliedEcology,2012,23(3):724-730.
[18]Magnani F,Mencuccini M,Grace J.Age-related decline in stand productivity:The role of structural acclimation under hydraulic constraints [J].PlantCellandEnvironment,2000,23:251-263.
[19]殷桂香,王 瑾,徐惠君,等.几个小麦基因型苗期抗旱性鉴定及相关生理指标分析[J].麦类作物学报,2009 ,29(2):319 -323.
Yin G X,Wang J,Xu H J,etal.Drought tolerance test and related physiological indexes analysis at seedling stage of several wheat genotypes [J].JournalofTriticeaeCrops,2009,29(2):319-323.
[20]张永强,毛学森,孙宏勇,等.干旱胁迫对冬小麦叶绿素荧光的影响[J].中国生态农业学报,2002,10(4):13-15.
Zhang Y Q,Mao X S,Sun H Y,etal.Effects of drought stress on chlorophyll fluorescence of winter wheat [J].ChineseJournalofEco-Agriculture,2002,10(4):13-15.
[21]杨晓青,张岁岐,梁宗锁,等.水分胁迫对不同抗旱类型冬小麦幼苗叶绿素荧光参数的影响[J].西北植物学报,2004,24(5):812-816.
Yang X Q,Zhang S Q,Liang Z S,etal.Effects of water stress on chlorophyll fluorescence parameters of different drought resistance winter wheat cultivars seedlings [J].ActaBotanicaBoreali-OccidentaliaSinica,2004,24(5):812-816.
[22]孙 萍,段喜华.干旱胁迫对长春花光合特性及可溶性糖的影响[J].东北林业大学学报,2010,38(8):54-56.
Sun P,Duan X H.Effects of drought stress on soluble sugars and photosynthetic characteristics ofCatharanthusroseusseedlings [J].JournalofNortheastForestryUniversity,2010,38(8):54-56.
[23]王 川,谢惠民,王 娜,等.小麦品种可溶性糖和保护性酶与抗旱性关系研究[J].干旱地区农业研究,2011,29(5):94-99.
Wang C,Xie H M Wang N,etal.Study on the relation of soluble sugar,protective enzyme and drought resistance in wheat [J].AgriculturalResearchintheAridAreas,2011,29(5):94-99.
[24]马守才,张改生,王军卫,等.小麦种子活力性状的遗传变异和相关研究[J].西北植物学报,2004,24(9):1674-1679.
Ma S C,Zhang G S,Wang J W,etal.Genetic difference and interrelationship among seed vigor traits in wheat [J].ActaBotanicaBoreali-OccidentaliaSinica,2004,24(9):1674-1679.
[25]Soltani A,Gholipoor M,Zeinali E.Seed reserve utilization and seedling growth of wheat as affected by drought and salinity [J].EnvironmentalandExperimentalBotany,2006,55:195-200.
[26]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006:71-76.
Gao J F.Plant Physiology Experiment Instruction [M].Beijing:Higher Education Press,2006:71-76.
Effect of Drought Stress on Seed Germination and Biomass Allocation of Root and Shoot of Different Drought Resistant Wheat Cultivars
SHI Chengxiao,CHEN Ting,WANG Changjiang,QIN Xiaoliang,LIAO Yuncheng
(College of Agronomy,Northwest A&F University,Yangling,Shaanxi 712100,China)
Abstract:In order to explore the effects of drought stress on seed germination and seedling growth of different drought resistant wheat cultivars,an experiment was conducted to determine and analyze 16 indices,including germination rate,root and shoot dry weight,ratio of root to shoot,etc. with drought-tolerant variety Changhan 58 and drought-sensitive variety Luomai 18 under the treatments of 15% PEG-6000 and control. The results showed that drought stress had no effect on seed germination rate of Changhan 58,but seed germination rate of Luomai 18 was significantly decreased under drought stress. The seed germination vigor,germination index,vitality index and translocation of seed reserves of Changhan 58 were significantly greater than those of Luomai 18,but the average germination time was significantly lower than that of Luomai 18;root length,shoot length,root dry weight,shoot dry weight and accumulation of seedling dry matter of Changhan 58 were significantly higher than those of Luomai 18 under drought stress. In addition,the root-shoot ratio of Luomai 18 was increased significantly under drought stress,but there was no significant effect on seedling root-shoot ratio of Changhan 58 on the 5th and 7th day. The leaf chlorophy content of Luomai18 was significantly decreased under drought stress,however,there was no significant difference between 15% PEG-6000 and control for Changhan 58. From these results,it can be concluded that Luomai 18 and Changhan 58 can be selected as ideal model cultivars for research on drought resistance in future.
Key words:Wheat;Drought stress;Seed germination;Biomass allocation;Root shoot ratio;Translocation of seed reserves
中图分类号:S512.1;S311
文献标识码:A
文章编号:1009-1041(2016)04-0483-08
通讯作者:廖允成(E-mail:yunchengliao@163.com);秦晓梁(E-mail:xiaoliangqin2006@163.com)
基金项目:国家科技支撑计划项目(2015BAD22B03-05);陕西省科技统筹创新工程计划项目(2015KTZDNY01-02)
收稿日期:2015-11-16修回日期:2015-12-29
网络出版时间:2016-04-01
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160401.1533.028.html
第一作者E-mail:shichengxiao0509@163.com
