耐热普鲁兰酶CBM68结构域中关键位点对其酶学性质的影响
2016-05-09申莹莹郑宏臣李树芳付晓平徐健勇宋诙
申莹莹,郑宏臣,李树芳,付晓平,徐健勇,宋诙*
1(天津科技大学 生物工程学院,天津, 300457) 2(中国科学院 天津工业生物技术研究所,工业酶国家工程实验室,天津, 300308) 3(中国科学院 天津工业生物技术研究所,天津工业生物系统与过程重点实验室,天津, 300308)
耐热普鲁兰酶CBM68结构域中关键位点对其酶学性质的影响
申莹莹1,2,3,郑宏臣2,3,李树芳2,3,付晓平2,3,徐健勇2,3,宋诙2,3*
1(天津科技大学 生物工程学院,天津, 300457)2(中国科学院 天津工业生物技术研究所,工业酶国家工程实验室,天津, 300308)3(中国科学院 天津工业生物技术研究所,天津工业生物系统与过程重点实验室,天津, 300308)
摘要通过对Anoxybacillus sp. LM18-11来源的耐热普鲁兰酶(PulA)与麦芽三糖共结晶的晶体结构分析,在新命名的底物结合域CBM68中预测出4个与底物结合有关的氨基酸位点(Y14、D16、K62和R96),分别将4个氨基酸位点突变为丙氨酸,得到突变体PulA(Y14A)、PulA(D16A)、PulA(K62A)和PulA(R96A)。其中,突变酶PulA(Y14A) 和PulA(R96A)的底物结合力及催化效率与野生酶PulA相比均有明显降低,其Km值分别比PulA提高了4.2倍和2.5倍,k(cat)/Km 分别为PulA的29%和37%。同时,PulA(Y14A) 和PulA(R96A)的最适作用温度均降低了10 ℃,PulA(R96A)的最适作用pH也降低了0.5个单位。说明Y14和R96是CBM68结构域中的关键氨基酸位点,这两个位点的发现为进一步探究CBM68结构域的作用机制奠定了基础。
关键词耐热普鲁兰酶;CBM68;定点突变;底物亲和力;催化效率
普鲁兰酶是淀粉水解过程中一个重要的脱支酶,能够专一性地水解支链淀粉分支点中的α-1, 6糖苷键[1]。与α-淀粉酶、β-淀粉酶或葡糖淀粉酶共同作用,可使淀粉充分水解为小分子糖,提高淀粉原料的利用率,广泛应用于以淀粉为原料的食品深加工和新生物质能源等工业领域[2-3]。普鲁兰酶属于糖苷水解酶类,其分子结构一般由多个结构域组成,除了具有催化作用的催化结构域外,还包含功能各异的其他非催化结构域。其中,糖基结合模块 (CBM) 是广泛存在于糖基水解酶中的非催化性结构域,该结构域对糖苷水解酶的底物特异性、底物结合能力及结构稳定性起到重要作用[4]。目前,在普鲁兰酶分子中出现频率较高的CBM结构域家族包括CBM41、CBM48和CBM68等[5-6]。
近年来对CBM结构域的分子结构与功能的研究越来越深入,也取得了一些成果,发现不同的CBM结构域对酶的表达、酶学性质、动力学参数以及极端酶(高温酶、耐酸酶和耐碱酶等)的催化性能等显示了不同程度的影响。例如,VENDITTO 等人将嗜碱芽孢杆菌Bh Cel5B的内切葡聚糖酶C端的CBM46结构域去除后,发现CBM46缺失酶的底物亲和力明显下降[7];STROBEL 等人探究了里氏木霉Cel7A的CBM结构域对木质素的亲和力,通过定点突变将找到的31个氨基酸位点统一突变为丙氨酸,证明了提高酶对底物的选择性是增加酶催化能力的必要条件[8];此外,日本科研人员对几丁质酶A1(ChiA1) CBM12结构域进行了结合活性的研究,采用定点突变的方法对选择的14个氨基酸同样突变为丙氨酸,除了687位的芳香族氨基酸与结合活性有关外,非保守性的679位谷氨酰胺的突变也导致了结合活性的显著降低,但核磁共振结果显示该位点突变并没有改变蛋白的整体结构[9];江南大学生物技术重点实验室研究了分别截掉普鲁兰酶N端CBM41结构域、CBM41+X25结构域和CBM41+X45+X25结构域,所得到的突变酶在酶学性质及分别表达的变化,从而发现CBM41和 X25对该酶的热稳定性有一定的不利影响,X45对该酶活性的保持是必须的[1, 10]。目前对于CBM结构域的关键结合位点及其的催化功能的影响机制还有待进一步研究。
本实验室在前期的研究中获得了1株耐热普鲁兰酶产生菌Anoxybacillussp.LM18-11[11],并从中获得了耐热I型普鲁兰酶PluA,晶体结构分析显示PluA具有4个相对独立的结构域 (N1、N2、A、C),N末端存在两个连续的CBM,N2 区属于CBM48,而N1区不属于已知的任何CBM家族,被CAZy首次认定为一个新的CBM家族,即CBM68[12]。前期对CBM68结构域的缺失实验表明突变体的底物亲和力、最适温度以及热稳定性均较野生型有明显降低[11],可见CBM68对PulA的催化性能起到了重要作用,因此本论文在PulA与小分子麦芽三糖共结晶的基础上,根据序列比对及酶与小分子相互作用的结构分析,在CBM68结构域中选取4个可能与底物结合相关的氨基酸位点(Y14、D16、K62和R96),并对其分别进行定点突变研究,揭示CBM68中的关键作用位点及其对普鲁兰酶催化性质的影响,为进一步探究新结构域CBM68的作用机制奠定一定理论基础。
1材料与方法
1.1材料
1.1.1菌种和质粒
大肠杆菌DH5α和BL21trxB(DE3)、载体pET32a为本实验室收藏。
1.1.2酶和主要试剂
Phusion热启动II高保真DNA聚合酶,ThermoFisher公司; 各种抗生素、普鲁兰糖、IPTG,Sigma公司;蛋白质纯化用HisTrap柱、XK16/70凝胶柱,GE公司;引物由中国科学院天津工业生物技术研究所合成。
1.1.3培养基
LB培养基(g/L):酵母粉 5,胰蛋白胨 10,NaCl10;添加 2% 琼脂为固体培养基。
1.1.4定点突变引物

引物名称引物序列(5’-3’)Y14AFGTCGTTTGAAGCTGCATTAGATGAAT-TGACRTGCAGCTTCAAACGACTGTTGTTTT-GD16AFGAAGCTTATTTAGCGGAATTGA-CAATGRCGCTAAATAAGCTTCAAACGACTGT-TGK62AFCATCATTCATTTATAGCGTATGAGT-GTCTCRCGCTATAAATGAATGATGCTTCT-CACTTGAR96AFGTAGGAGCTGTGATGGC-CACGAAAGCGTTTRGGCCATCACAGCTCCTACTTGCAATG-
1.2方法
1.2.1突变反应
以pET32a-pulA为模板,采用Phusion热启动II高保真DNA聚合酶进行PCR突变反应。反应体系(50 μL):10 μL 5× Phusion HF缓冲液,4 μL dNTPs(各2.5 mmol/L),1 μL 模板DNA,1.5 μL 正向引物,1.5 μL 反向引物,1.5 μL DMSO,0.5 μL Phusion热启动II高保真DNA聚合酶(2 U/μL),补加双蒸水至50 μL。反应条件:98 ℃预变性 3 min;98 ℃变性 10 s,55 ℃退火 30 s,72 ℃延伸 4 min,共30个循环;72 ℃再延伸10 min。PCR产物经1%琼脂糖凝胶电泳进行鉴定。
1.2.2筛选阳性克隆
电泳目的条带位置正确的PCR产物在37 ℃经Dpn I消化模板质粒1 h后,转DH5α感受态,37 ℃培养12 h,挑选单克隆,提质粒,经1%琼脂糖凝胶电泳进行鉴定,将电泳目的条带位置正确的质粒送北京六合华大基因科技股份有限公司测序。
1.2.3蛋白表达与纯化
基因位点突变正确的重组质粒转化到大肠杆菌BL21trxB(DE3),将含有突变重组质粒的大肠杆菌BL21trxB(DE3)单菌落接种于4 mL液体LB(卡那霉素50 μg/mL、氨苄青霉素100 μg/mL)培养基中,于37 ℃、220 r/min下振荡培养12 h,然后以1%的接种量转接到200 mL含卡那霉素50 μg/mL和氨苄青霉素100 μg/mL的液体LB培养基中,37 ℃、220 r/min培养至OD600=0.6~0.8,加IPTG终浓度至0.05 mmol/L,在16 ℃、160 r/min下进行蛋白诱导表达,16 h后于4 ℃、5 000 r/min离心20 min收集菌体,高压均质破碎仪破碎细胞,蛋白经His-Trap(5 mL)柱、XK16/70凝胶柱纯化,纯化后的蛋白经适当稀释用SDS-PAGE进行纯度分析。
1.2.4普鲁兰酶酶活力测定
DNS方法[13]:在相应的温度下,取450 μL反应缓冲液 (5%的普鲁兰糖溶液和相应的pH缓冲液以1∶8的比例混合均匀),加入50 μL适当稀释的普鲁兰酶酶液混合均匀,反应30 min,加入500 μL DNS溶液终止反应,100 ℃水浴10 min,测定OD540值。
酶活力单位定义为:在特定的反应条件下,每分钟水解普鲁兰糖产生相当于1 μmol葡萄糖的还原糖,所需的酶量为1个活力单位(U)。
1.2.5突变酶最适pH值及pH稳定性测定
采用pH 4.0~5.8乙酸-乙酸钠缓冲液和pH 5.8~9.0磷酸氢二钠-磷酸二氢钠缓冲液分别配制pH 4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0缓冲液,取450 μL反应缓冲液(5%的普鲁兰糖溶液和相应的pH值缓冲液以1∶8的比例混合均匀),加入50 μL适当稀释的酶液,混合均匀后55 ℃保温30 min,然后加入500 μL DNS溶液终止反应,100 ℃水浴10 min,测定OD540值。将普鲁兰酶液在以上不同pH值缓冲液中于4 ℃处理4 h,在最适条件下按以上方法测定残余酶活力。
1.2.6突变酶最适温度及耐热稳定性测
在35~80 ℃范围内测定突变酶最适温度,取450 μL反应缓冲液(5%的普鲁兰糖溶液和相应的pH 6.0磷酸缓冲液以1∶8的比例混合均匀),加入50 μL适当稀释后的酶液,混合均匀后分别在35、40、45、50、55、60、65、70、75、80 ℃下保温反应30 min,然后加入500 μL DNS溶液终止反应,100 ℃水浴10 min,测定OD540值。另外,取适当稀释的突变酶500 μL加入10 μL石蜡油覆盖酶液,置于60 ℃水浴锅温育不同时间,在最适条件下按以上方法测定残余酶活力。
1.2.7突变酶底物特异性分析[14-15]
取50 μL适当稀释的酶液与400 μL磷酸缓冲液(20 mmol/L 、pH 6.0)混合,分别以5%的普鲁兰糖、可溶性淀粉、支链淀粉、糖原做反应底物,反应体系底物终浓度为5 mg/mL,55 ℃保温30 min,然后加入500 μL DNS溶液终止反应,100 ℃水浴10 min,测定OD540值。以最高酶活力为100%,计算不同底物的相对酶活力。
1.2. 8突变酶动力学分析
取450 μL用pH 6.0磷酸缓冲液配制的不同浓度的普鲁兰糖溶液(反应缓冲液中普鲁兰糖的终质量浓度为1.0,1.25,1.5,2.0,2.5,3和5.0 mg/mL),在55 ℃预热5 min后加入50 μL适当稀释的突变蛋白酶液,混合均匀后继续保温10 min,然后加入500 μL DNS溶液终止反应,100 ℃水浴10 min,测定OD540值。建立1/V-1/[S]的双倒数曲线,计算其Km值、Vmax值、kcat值及kcat/Km值。
2结果与分析
2.1普鲁兰酶突变体表达菌株的构建
以pET32a-PulA质粒为模板,应用1.1.4所示的定点突变引物,经Phusion热启动II高保真DNA聚合酶进行PCR突变反应,得到相应的突变质粒,质粒全长8kb左右 (图1)。经基因测序,得到突变正确的重组质粒分别命名为pET32a-PulAY14A、pET32a-PulAD16A、pET32a -PulAK62A和pET32a-PulAR96A(图2)。然后将测序正确的突变质粒分别转化到E.coliBL21trixB(DE3)中,成功构建了突变酶重组表达菌株E.coliBL21trixB(pET32a-PulAY14A)、E.coliBL21trixB(pET32a-PulAD16A)、E.coliBL21trixB(pET32a-PulAK62A)和E.coliBL21trixB(pET32a-PulAR96A)。
M1、M2、M3、M4: DNA marker 1: pET32a-PulAY14A 2: pET32a-Pu-lAD16A3: pET32a-PulAK62A 4: pET32a- PulAR96A图1 Phusion DNA聚合酶定点突变PCR反应结果Fig.1 Gel electrophoresis of mutant products using Phusion DNA polymerase

图2 阳性克隆突变位点的测序图 Fig.2 The DNA sequences of pullulanase mutants surrounding mutagenesis position图示为正义链序列, 箭头所指为定点突变核苷酸;A:pET32a-PulAY14A中PulA的14位酪氨酸TAT突变为丙氨酸GCA;B:pET32a-PulAD16A中PulA的16位天冬氨酸GAT突变为丙氨酸GCG;C:pET32a-PulAK62A中PulA的62位赖氨酸AAA突变为丙氨酸GCG;D:pET32a-PulAR96A中PulA的96位精氨酸CGC突变为丙氨酸GCC。
2.2普鲁兰酶PulA及其突变酶的诱导表达与纯化
将得到的大肠杆菌重组工程菌分别进行摇瓶培养,应用IPTG诱导重组普鲁兰酶的表达,然后应用Ni2+亲和柱、XK16/70凝胶柱对重组普鲁兰酶进行纯化。纯化后的蛋白经SDS-PAGE蛋白电泳,结果如图3所示,PulA与各点突变普鲁兰酶均得到了较好的纯化,其蛋白分子量均为100 kDa左右。将点突变后的普鲁兰酶分别命名为PulAY14A、PulAD16A、PulAK62A和PulAR96A。
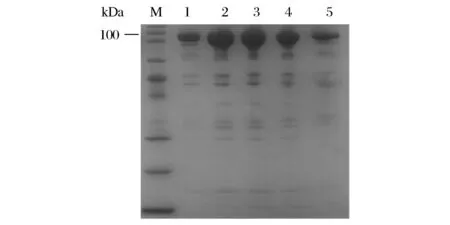
M-protein marker;1-PulA;2-PulAY14A;3-PulAD16A;4-PulAK62A;5-PulAR96A图3 纯化后普鲁兰酶PulA及其突变体的SDS-PAGE分析Fig.3 The SDS-PAGE analysis of PulA and its mutants
2.3氨基酸突变位点对普鲁兰酶最适pH值及其pH稳定性的影响
将纯化后的普鲁兰酶PulA和4个点突变的普鲁兰酶分别在不同温度下测定酶活,结果如图4a所示。突变酶PulAY14A、PulAD16A和PulAK62A在pH 4.5~8.0范围内显示了与PulA相似的趋势曲线,最适作用pH均为6.0,而突变酶PulAR96A的最适作用pH为5.5,并且在pH 5.5条件下,4个突变酶的相对酶活均高于野生型普鲁兰酶PulA。同时,4个突变普鲁兰酶在不同pH下的稳定性曲线均与野生型普鲁兰酶PulA相似 (图4b)。可见,除了R96A突变位点外,其余3个氨基酸位点的突变对PulA的最适pH及pH稳定性的影响并不明显,而R96A突变使得PulA的最适作用pH向酸性偏移了0.5个单位,说明R96位点对PulA的酸碱催化过程具有重要影响。
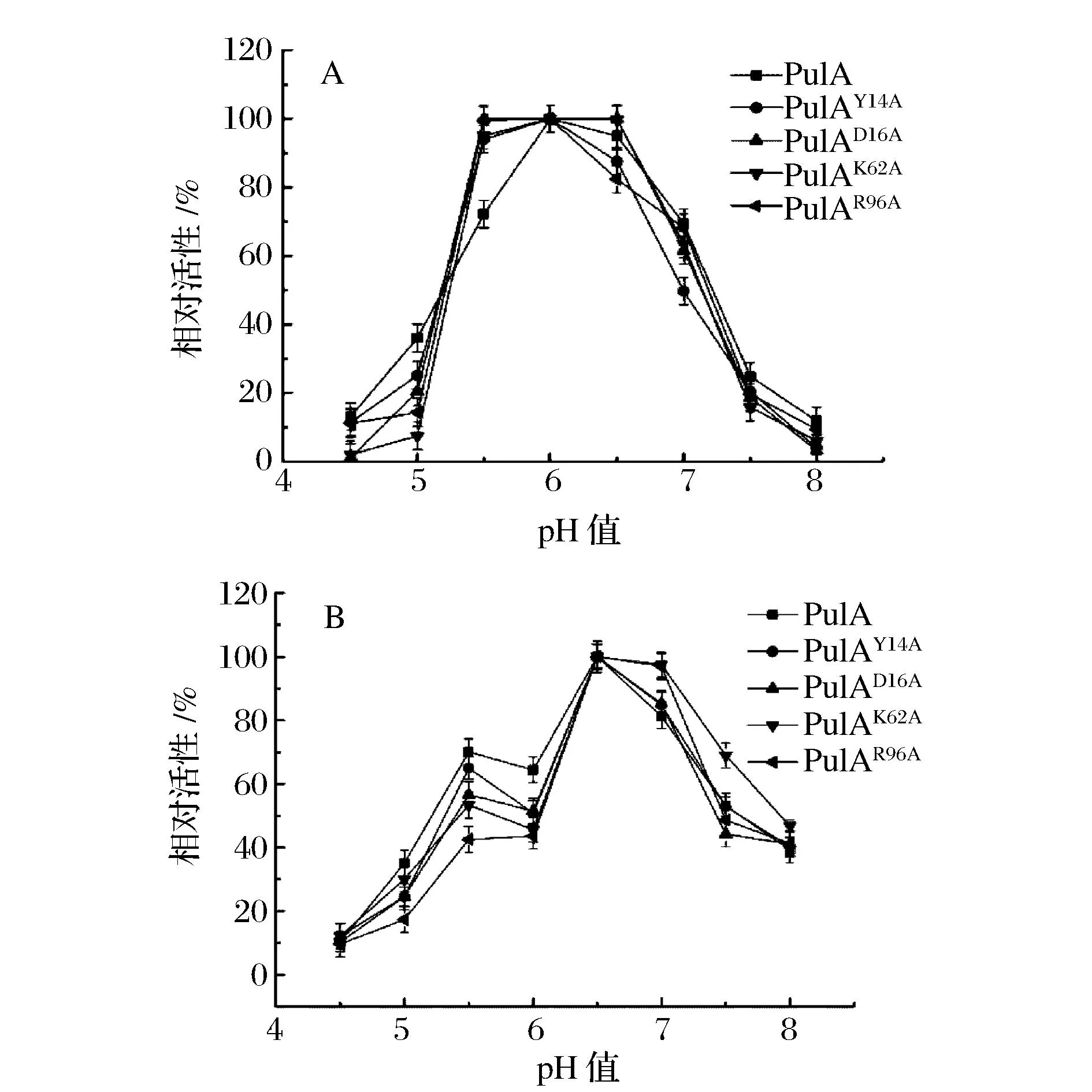
图4 重组普鲁兰酶最适作用pH及pH稳定性Fig.4 The optimum pH and pH stability of the recombinant pullulanases
2.4氨基酸突变位点对普鲁兰酶最适温度及温度稳定性的影响
重组普鲁兰酶在不同温度下测定酶活力,结果如图5a所示,PulAD16A和PulAK62A的最适作用温度为55~60 ℃,和PulA的最适作用温度相同,而PulAY14A和PulAR96A的最适作用温度为50 ℃,并且PulAR96A在60 ℃时相对酶活降到50%。另外,PulAR96A在60 ℃的半衰期为8 h,比PulA的半衰期48 h降低了5倍 (图5b)。当保温时间到达25 h时,PulAR96A几乎完全失活。可见,Y14A和R96A的点突变可使PulA的耐热性降低,尤其是R96A的作用最为明显。经分析,将PulA96位侧链为长链的精氨酸突变为短链的丙氨酸,可能改变了蛋白的结构柔性,使其在高温下维持蛋白三级结构的能力降低,蛋白分子更容易变性,从而导致该突变酶的最适温度和热稳定性都显著降低[16]。
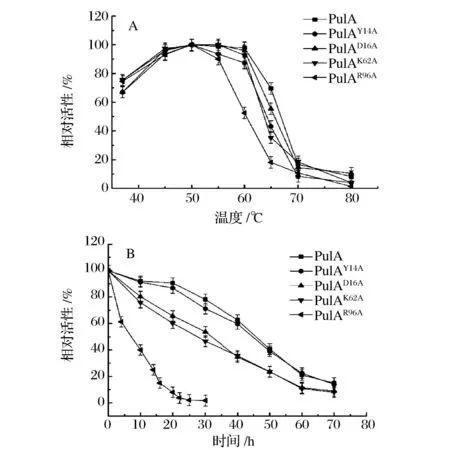
图5 重组普鲁兰酶的最适作用温度及耐热稳定性Fig.5 The optimum temperature and thermostability of the recombinant pullulanases
2.5氨基酸突变位点对普鲁兰酶底物特异性的影响
分别选取普鲁兰糖、支链淀粉、可溶性淀粉以及糖原为底物在普鲁兰酶的最适作用条件下测定酶活 ,结果如图6所示,4个突变普鲁兰酶与PulA一样,均对普鲁兰糖显示了最高催化活性,对支链淀粉和可溶性淀粉催化活性相对较低,对糖原几乎没有催化作用。其中,突变体PulAY14A对支链淀粉和可溶性淀粉的催化活性完全丧失,可见14位上这种高保守型的芳香族氨基酸(Tyr)对底物特异性具有重要影响。另外,PulAD16A与PulA相比,其对支链淀粉的催化活性几乎降低为零,而对可溶性淀粉的催化作用却明显增强了。而PulAK62A和PulAR96A对支链淀粉的催化活性完全丧失,对可溶性淀粉的催化作用影响不大。综上,Y14A和R96A定点突变不但影响该酶的作用温度及pH,对其底物特异性也具有显著影响,而D16A和K62A定点突变虽然促使该酶的底物特性发生了一定变化,但对该酶其他性质影响不大,可见,4个位点的突变对该酶酶学性质和底物特异性的影响机制可能是不同的。
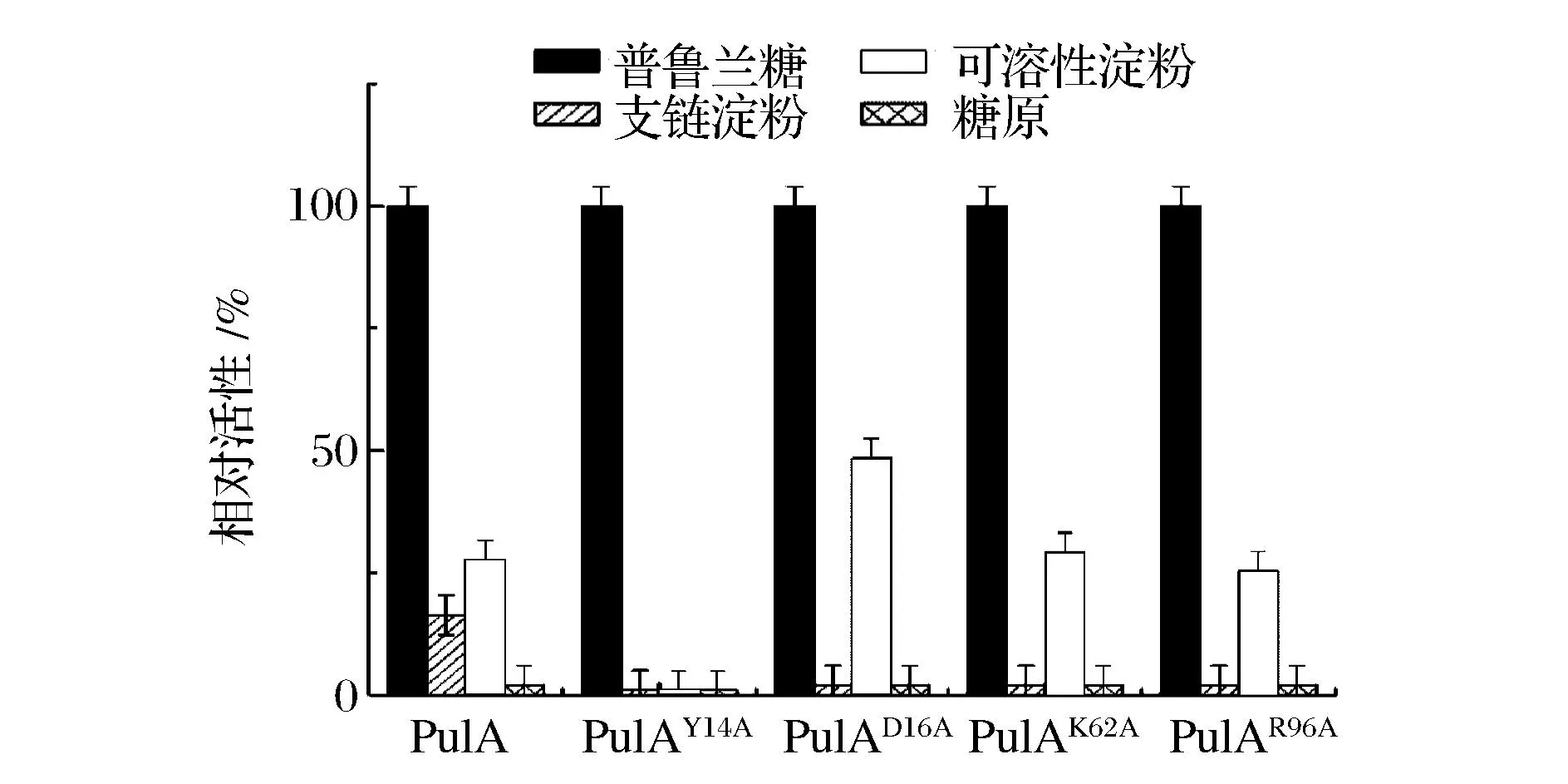
图6 普鲁兰酶PulA及其突变体的底物特异性Fig.6 Substrate specificity of PulA and its mutants
2.6氨基酸突变位点对普鲁兰酶动力学参数的影响
以普鲁兰糖为底物分别对重组普鲁兰酶动力学参数进行测定和比较,结果如表1所示,PulAD16A和PulAK62A的动力学参数与PulA非常接近,说明D16A和 K62A突变对PulA底物亲和力及催化效率没有影响。而PulAY14A和PulAR96A的Km值分别比PulA提高了4.2倍和2.5倍,kcat/Km分别为PulA的29%和37%。可见,Y14A和R96A突变严重降低了PulA的底物亲和力及催化效率。说明Y14和R96是PulA中CBM68结构域的关键氨基酸位点。将CBM68结构域的保守芳香族氨基酸Y突变成A,对底物的亲和力性影响较大,Y的苯环与环状糖分子之间靠范德华力形成堆叠作用,可能决定酶与底物结合的方式及亲和力[16]。而PulAR96A亲和力显著降低可能是因氨基酸侧链变短,致使蛋白结构柔性增大[10],从而降低其底物亲和力。至于这两个突变体催化效率的明显降低,很可能是亲和力的降低促使底物被运送到活性中心的速率降低所致。

表1 普鲁兰酶PulA及其突变体动力学参数
3结论
本论文为研究耐热普鲁兰酶PulA中新鉴定的底物结合结构域CBM68的作用机制,成功构建了4个单点突变的普鲁兰酶突变体PulAY14A、PulAD16A、PulAK62A和PulAR96A,经底物特异性、动力学参数及其他酶学性质的测定与对比分析,发现Y14和R96是CBM68中的两个关键氨基酸位点,对这两个关键氨基酸位点的突变不仅显著影响普鲁兰酶PulA的底物亲和力及催化效率,也对其作用温度、pH及稳定性具有一定的影响,然而具体的影响机制仍需进一步揭示。
参考文献
[1]DUAN Xu-guo,WU Jin.Enhancing the secretion efficiency and thermostability of a Bacillus deramificans pullulanase mutant (D437H/D503Y) by N-terminal domain truncation[J].Appl Environ Microbiol,2015,81(6):1 926-1 931.
[2]姜楠,宋诙,王萍.普鲁兰酶及其分泌相关蛋白的研究进展[J].微生物学报,2011,51(6):725-731.
[3]SIEW Ling Hii,JOO Shun TAN,TAU Chuan Ling,et al. Pullulanase:Role in starch hydrolysis and potential industrial applications[J].Enzyme Res,2012,2012:921 362.
[4]BORASTON A B,HEALEY M,KLASSEN J,et al. A structural and functional analysis of alpha-glucan recognition by family 25 and 26 carbohydrate-binding modules reveals a conserved mode of starch recognition[J].Biol Chem, 2006,281(1):587-98.
[5]MIKAMI B,IWAMOTO H,MALLE D,et al. Crystal structure of pullulanase:evidence for parallel binding of oligosaccharides in the active site[J].Mol Biol,2006,359(3):690-707.
[6]TURKENBURG J P,BRZOZOWSKI A M,SVENDSEN A,et al. Structure of a pullulanase from Bacillus acidopullulyticus[J].Proteins,2009,76(2):516-519.
[7]VENDITTO I, NAJMUDIN S,Lus A S,et al. Family 46 carbohydrate-binding modules contribute to the enzymatic hydrolysis of xyloglucan and β-1,3-1,4-Glucans through distinct mechanisms[J]. Journal of Biological Chemistry,2015,290(17):10 572-10 586.
[8]STROBEL K L,PFEIFFER K A,HARVEY W B,et al. Structural insights into the affinity of Cel7A carbohydrate-binding module for lignin[J].Journal of Biological Chemistry,2015,290(37):22 818-22 826.
[9]HARA M,SUGIMOTO H,UEMURA M,et al. Involvement of Gln679,in addition to Trp687,in chitin-binding activity of the chitin-binding domain of chitinase A1 from Bacillus circulans WL-12[J].Journal of Biochemistry, 2013,154(2):185-193.
[10]DUAN Xu-guo,CHEN Jian,WU Jing.Improving the thermostability and catalytic efficiency of Bacillus deramificans pullulanase by site-directed mutagenesis[J].Applied and Environmental Microbiology,2013, 79(13):4 072-4 077.
[11]甄杰,胡政,李树芳,等.一个新型耐热普鲁兰酶的结构与功能[J].生物工程学报,2014,30(1):119-128.
[12]XU Jiang-yong,REN Fei-fei,HUANG Chun-hsiang,et al. Functional and structural studies of pullulanase from Anoxybacillus sp. LM18-11[J].Proteins,2013,82(9):1 685-1 693.
[13]李红,孙红军.国内外普鲁兰酶市场分析及预测报告(2003、2004年度)[R].北京:理德斯普企业管理咨询有限公司,2004.
[14]ZHOU Cheng,XUE Yan-feng,ZHANG Yue-ling,et al. Recombinant expression and characterization of Thermoanaerobacter tengcongensis MB4 thermostable α-glucosidase with regioselectivity for high yield isomaltooligosaccharides synthesis[J].Journal of Microbiology and Bchnology,2009,19(12):1 547-1 556.
[15]CHEN C,XUE Yan-fen,MA Yan-he.Enhancing the thermostability of α-glucosidase fromThermoanaerobactertengcongensisMB4 by single proline substitution[J].Bioscience and Bioengineering,2010,110(1):12-17.
[16]GUILLEN D,S SANCHEZ,RODRIGUEZ-SANOJA R.Carbohydrate-binding domains: multiplicity of biological roles[J]. Appl Microbiol Biotechnol,2010,85(5):1 241-1 249.
Key amino acid sites in the CBM68 structure of thermostable pullulanase and their effects on the enzymatic properties
SHEN Ying-ying1,2,3,ZHENG Hong-chen2,3,LI Shu-fang2,3,FU Xiao-ping2,3,XU Jian-yong2,3,SONG Hui2,3*
1(College of Biotechnology, Tianjin University of Science and Technology, Tianjin 300457, China)2(Industrial Enzymes National Engineering Laboratory, Tianjin Institute of Industrial Biotechnology, Chinese Academy of Sciences, Tianjin 300308, China)3(Key Laboratory of Tianjin Industrial Biosystem and Process Engineering, Tianjin Institute of Industrial Biotechnology, Chinese Academy of Sciences, Tianjin 300308, China)
ABSTRACTBased on the analysis of the crystalline structure of pullulananse (PulA)-maltotetraose complex, four key amino acid sites (Y14, D16, K62 and R96) in a new carbohydrate-binding module CBM68 closely related to substrate binding were discovered. The thermostable pullulanase (PulA) was derived from Anoxybacillus sp. LM18-11. Four mutants of PulA(Y14A), PulA(D16A), PulA(K62A) and PulA(R96A) were obtained respectively by site-directed mutation. Among them, PulA(Y14A) and PulA(R96A) showed lower affinity for substrate and lower catalytic efficiency compared with the wild-type pullulanase (PulA). The Km values of PulA(Y14A) and PulA(R96A) were 4.2 times and 2.5 times higher than that of PulA, respectively. The k(cat)/Km values of PulA(Y14A) and PulA(R96A) only remained 29% and 37% of the original k(cat)/Km value (PulA), respectively. Moreover, the optimum temperatures for both PulA(Y14A) and PulA(R96A) were 10 ℃ lower than that for the wild-type pullulanase (PulA). And PulA(R96A) also showed decreasing of the optimum pH by 0.5 units. All the results in this work have proved the significance of the key amino acid sites Y14 and R96 on the structure CBM68. Further study on the function of the two key amino acid sites could enhance the understanding of the functional mechanism of CBM68.
Key wordsthermostable pullulanase; CBM68; site-directed mutation; substrate affinity; catalytic effeciency
收稿日期:2015-11-06,改回日期:2015-12-04
基金项目:国家863计划(2012AA022205);天津市科学基金(13JCYBJC39500)
DOI:10.13995/j.cnki.11-1802/ts.201603003
第一作者:硕士研究生(宋诙研究员为通讯作者,E-mail:song_h@tib.cas.cn)。
