百山祖常绿阔叶林优势种群的生态位
2016-05-09吴友贵叶珍林周荣飞骆争荣
吴友贵,叶珍林,周荣飞,杨 辉,骆争荣
(1.浙江凤阳山-百山祖国家级自然保护区百山祖管理处,浙江庆元323800;2.浙江农林大学林业与生物技术学院,浙江临安311300;3.丽水学院生态学院,浙江丽水323000)
百山祖常绿阔叶林优势种群的生态位
吴友贵1∗,叶珍林1,周荣飞1,杨 辉2,骆争荣3
(1.浙江凤阳山-百山祖国家级自然保护区百山祖管理处,浙江庆元323800;2.浙江农林大学林业与生物技术学院,浙江临安311300;3.丽水学院生态学院,浙江丽水323000)
为了理解百山祖中山常绿阔叶林各优势种群在群落中的关系、地位和功能,用定量分析法,对该群落中平均重要值大于1 的13个优势种群的生态位特征进行了研究。结果表明:多脉青冈、麂角杜鹃和窄基红褐柃的生态位宽度较大,它们的Hurlbert生态位宽度(Ba)分别为0.878、0.825、0.806;而厚叶红淡比、亮叶水青冈和水丝梨的生态位宽度较小,它们的Ba分别为0.341、0.320、0.272。物种的生态位宽度与其重要值大小无关,但与其在各样方中的重要值均匀程度呈极显著的线性正相关。生态位相似性比例(Cih)最大的是多脉青冈与尖连蕊茶(0.832),最小的是凸脉冬青和水丝梨(0.248)。对生境需求相似而生态位宽度较小的厚叶红淡比与亮叶水青冈的Cih高达0.758。同属植物多脉青冈与褐叶青冈、台湾冬青与凸脉冬青之间并不具有较大的Cih值。生态位宽度较大的物种与其他物种的生态位重叠值一般较大;生态位相似性比例大的物种的生态位重叠值一般较大。多脉青冈与褐叶青冈、台湾冬青与凸脉冬青的生态位重叠值并不大。总体来看,群落优势种群之间的生态位重叠程度较低,说明该群落主要优势种群之间竞争不激烈,群落稳定。该研究揭示了群落优势种群的生态位特征和结构稳定性,为生物多样性保护、群落演替和种群进化等研究提供了理论基础,也为自然保护区管理机构制定和完善保护措施提供参考依据。
常绿阔叶林,优势种群,生态位宽度,生态位相似性比例,生态位重叠,百山祖
∗通讯作者
生态位是物种在某一生态环境中入侵、定居、适应、繁衍、扩散以至衰退、消亡等历程的所有生态学过程中拥有的地位和功能,不仅反映该物种在某一时期某一生态环境里所占据的空间位置,而且反映物种在该生态环境中的土壤、水分等生态因子梯度上的位置,还可反映物种在整个生态系统的物质流、能量流和信息流中所扮演的角色(张光明和谢寿昌,1997)。生态位研究是种群生态学研究的重要方法之一,在群落演替、物种维护机制、生物多样性保护、植被恢复与森林培育、引种栽培以及人类行为学研究等领域得到广泛运用(王运生等,2008;朱春全和雷静品,1997;何中声等,2012;张大勇和姜新华,1997;程中秋等,2010;Paoli et al,2006;Thomas&Robea,2003)。
百山祖位于浙江省庆元县境内,地带性植被为常绿阔叶林,由于保护严格,在其主峰以下海拔1 250~1 750 m区域内分布着面积较大的常绿阔叶林,植被保存较为完好,在生物多样性保护、群落生态学研究等方面具有重要的价值(陈小荣等,2013;徐敏等,2007)。植物群落中优势种群的生态位特征是植物种群生态学研究的热点内容之一,研究该群落优势种群的生态位特征具有特殊的意义,通过研究可以理解群落内各种群对环境资源的利用情况以及物种之间的关系(史小华等,2007;刘金福和洪伟,1999;苏志尧等,2003;张国斌和李秀芹,2007;高俊香等,2010),可以了解各优势种群在群落中所处的地位、对其他种群的影响以及群落结构的稳定性,从而对群落演替和种群进化进行预测,为自然保护区管理机构及时制定和完善保护措施提供参考依据。
1 材料与方法
1.1 研究区概况
百山祖位于浙江百山祖国家级自然保护区境内,主峰海拔1 856.7 m。百山祖自然保护区位于Ⅱ-Ⅴ地带群落交错区(程秋波等,1996),植物物种丰富,珍稀特有物种较多(金孝锋等,2004),是全国17个具有全球意义的生物多样性保护关键区域之一——“浙、闽、赣交界山地”的重要组成部分(中国生物多样性国情研究组,1998)。以侵蚀地貌为主,土壤以黄壤为主,山地坡度大多在30°以上。属亚热带湿润季风气候,主要特点是温暖湿润多雨。百山祖气象观测点(海拔1 100 m)的观测资料(1982-2001)显示:年均气温12.8℃,最高气温32℃,最低气温-13℃,年均降水量2 341.8 mm,相对湿度84 ,无霜期187 d(余久华等,2003;金孝锋等,2004)。

图1 样方在样地中所处位置示意图Fig.1 Diagram of the sample plots in the large scale permanent plot
1.2 研究方法
百山祖自然保护区管理处于2003年在百山祖北坡常绿阔叶林中建成面积为5 hm2(250 m×200 m)的生物多样性固定监测样地,整个样地参照CTFS(Center for Tropical Forest Science)样地建设方案进行建设(徐敏等,2007),分成2 000个5 m×5 m的小样方,并于2003年8月完成第一次普查。调查样地中所有乔木及胸径1.0 cm以上的灌木的胸径DBH(实测)、树高(估)、冠幅(估),以及到最近两个顶点的水平距离(实测)(王伟等,2011;徐敏等,2007)。分别于2008年8月、2013年8月进行了复查。复查所有在上次调查中被记录的乔灌木个体,测量其胸径、生活状况。胸径进阶达到1.0 cm的个体,记录其种名、胸径、冠幅、树高及坐标。在大样地的各顶点、各边线的中点、中心位置附近共选取了9 个20 m×20 m的样方(样方在大样地中的相对位置如图1,各样方的基本概况见表1),提取2013年调查的相关数据进行统计处理。
1.3 统计方法
以所选取的样方为资源状态,以物种重要值为指标,对群落优势种群的生态位特征进行分析。物种的Hurlbert生态位宽度、生态位相似性比例、生态位重叠的计算方法参照胡正华等(2009)。
2 结果与分析
2.1 群落重要值整理
对9个样方进行重要值统计,胸径≥1 cm的所有木本植物共有65种,选取平均重要值大于1(即总和大于9 )的13个树种(占总树种的20 )进行生态位指标测定,它们在各样地的重要值见表2,这些树种的重要值总和平均占样方的78.80 。其中窄基红褐柃(Eurya rubiginosa var.attenuata)和尖连蕊茶(Camellia cuspidata)为灌木,麂角杜鹃(Rhododendron latoucheae)为小乔木,其余树种为乔木;多脉青冈(Cyclobalanopsis multinervis)与褐叶青冈(C.stewardiana)同为青冈属植物,而凸脉冬青(Ilex editicostata)与台湾冬青(I.formosana)同为冬青属植物。
2.2 生态位宽度分析
从表2可以看出,优势种群的生态位宽度(Ba)较大的是多脉青冈、麂角杜鹃和窄基红褐柃,它们的Ba值分别为0.878、0.825、0.806;生态位宽度较小的是厚叶红淡比、亮叶水青冈和水丝梨,它们的Ba值分别为0.341、0.320、0.272。多脉青冈、麂角杜鹃具有较大的重要值和生态位宽度,利用环境资源的能力较强,对于群落的结构组成、多样性维护、演替进程等功能方面都起着主导作用。作为小乔木的麂角杜鹃虽然占据林冠第二层,但在群落内分布广,在每个资源位中都有分布,数量众多,是样地内个体数量最多的物种,故其生态位宽度较大。窄基红褐柃、尖连蕊茶和浙闽新木姜子作为耐荫树种,在较密的林冠下能完成幼苗更新,故具有较大的生态位宽度。凸脉冬青、台湾冬青、厚叶红淡比、亮叶水青冈、水丝梨在部分资源位中没出现,分布不均匀,导致它们的生态位宽度较小,表明它们利用环境资源的利用能力较弱。在同属植物中,多脉青冈与褐叶青冈的生态位宽度差别较大,台湾冬青与凸脉冬青的生态位宽度差别不大。
另外发现,物种的生态位宽度与其重要值之间没有明显的相关性,如短尾柯重要值较大而生态位宽度中等;重要值中等的亮叶水青冈、厚叶红淡比和水丝梨,它们的生态位宽度却较小;而重要值小的窄基红褐柃和浙闽新木姜子,它们的生态位宽度却较大;褐叶青冈重要值较小而生态位宽度中等。Pij(即种i对第j个资源的利用占它对全部资源利用的频度)的标准差可以表示物种在各样方中的重要值的均匀程度,标准差越小表示越均匀,标准差越大表示越不均匀。从表2可以看出,Ba的变化趋势正好与Pij的标准差相反,即随着Pij标准差增大而Ba减小,它们之间存在着负相关性。将Pij标准差与Ba进行回归,拟合方程为Ba=1.0802-0.0550d(d表示Pij标准差,r=-0.9970,P<0.001),线性回归关系极显著。

表1 各样方的自然概况Table 1 Background of the evergreen broad-leaved forest plots
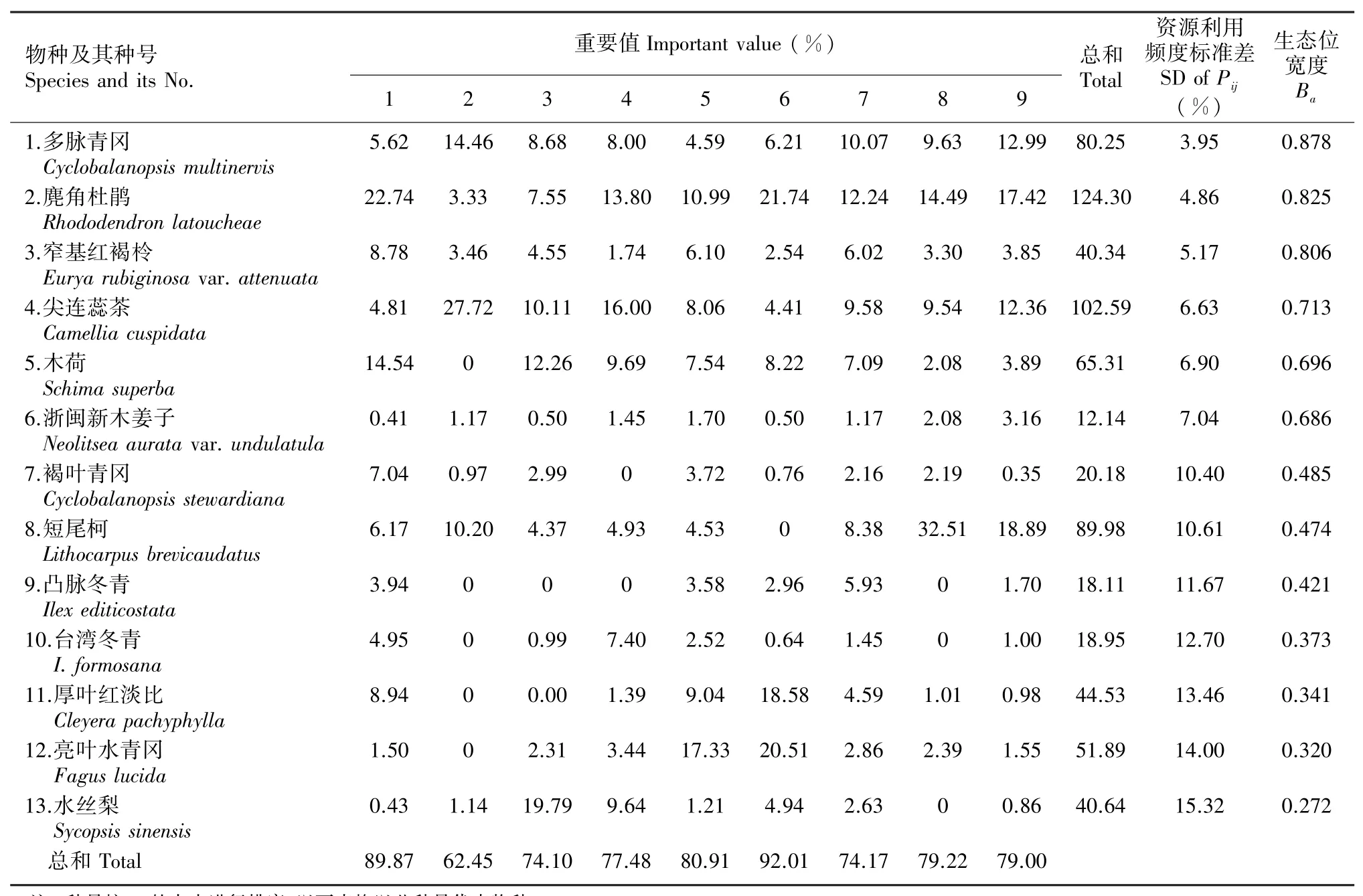
表2 优势种群的重要值和生态位宽度Table 2 Importance values and niche breadths of dominant tree species
2.3 生态位相似性比例分析
群落的生态位相似性比例(Cih)见表3,Cih值最大的是多脉青冈和尖连蕊茶(0.832),最小的是凸脉冬青和水丝梨(0.248)。Cih值介于0.7至0.8的13 对(占16.67 ),介于0.5至0.7的有30对(占 38.46 ),介于0.3至0.5的有30对(占38.46 ),介于0.2至0.3的有4对(占5.13 )。生态位宽度较大的物种之间的生态位相似性比例一般较高(胡正华等,2009;刘金福和洪伟,1999),如生态位宽度最大的3个物种为多脉青冈、麂角杜鹃、窄基红褐柃,它们之间的Cih分别达到0.747、0.730和0.741。将某一树种与其它树种之间的Cih进行平均,Cih平均值较大的是窄基红褐柃、麂角杜鹃、木荷、多脉青冈,综合表明这些物种与其它物种之间对资源的利用有着更大的相似性。从表3还可看出,生态位宽度较小的厚叶红淡比与亮叶水青冈的Cih高达0.758,表明这两物种对环境资源的利用较为相似,这或许与两物种的生物学、生态学特性有密切关系。在同属植物中,多脉青冈与褐叶青冈、台湾冬青与凸脉冬青并不具有较大Cih值,而是处于中等水平(分别为0.554、0.514)。该群落中生态位相似性比例大于0.5的有44对,占总对的56.41 ,所有Cih的平均值为0.538,表明群落优势种群利用环境资源的相似程度较高,体现了亚热带常绿阔叶林具有较高的生态位相似性比例的特点。

表3 优势种群的生态位相似性比例Table 3 Niche similarities of dominant tree species

表4 优势树种的生态位重叠值Table 4 Niche overlaps of dominant tree species
2.4 生态位重叠分析
群落13个主要种群间的生态位重叠情况如表4所示。由于种号顺序是按Ba的大小进行排序的,而B(L)的大小顺序与Ba一致,即对于每一对物种来说,B(L)i>B(L)h,所以表中生态位重叠值Lih大于相应的Lhi。Lih值最大的是窄基红褐柃与褐叶青冈,为0.127,Lhi值最大的是窄基红褐柃与麂角杜鹃,为0.096;Lih值和Lhi值最小的都是台湾冬青与水丝梨,分别为0.025和0.018。Lih大于0.1的有19个(占总数的12.18 ),Lhi都小于0.1。生态位宽度较大的物种与其他物种的生态位重叠值较大,如生态位宽度排在前5位的物种的Lih大于0.1的有17个(占大于0.1个数的89.47 )。物种之间的生态位相似性比例大,则它们的生态位重叠值一般较大,如Cih大于0.7的14对中有9对的Lih大于0.1,其余5对接近0.1;又如厚叶红淡比与亮叶水青冈的Lih/Lhi值为0.103/0.098。总体来看,群落主要优势种群之间的生态位重叠程度很低,在0.018~0.127之间波动,所有的Lih值和Lhi值平均为0.062。在同属植物中,多脉青冈与褐叶青冈、台湾冬青与凸脉冬青的生态位重叠值较小,Lih/Lhi值分别0.082/0.044、0.058/0.053。一般认为,同属物种亲缘关系较近,它们的生物学、生态学特性比较相似,对环境的需求也较为相似。在本研究中,多脉青冈与褐叶青冈、台湾冬青与凸脉冬青的生态位相似性比例中等,而生态位重叠值均较小。群落主要优势种群之间较低的生态位重叠程度和同属植物的生态位特征,表明百山祖中山常绿阔叶林群落稳定性较大,处于或接近于地带性顶极群落。
3 讨论
生态位宽度是指一个种群在群落中所利用的各种不同资源的总和,在可利用资源较少的条件下,生态位宽度往往会变宽,才能使种群得到充足的资源,而在可利用资源相对充裕的条件下,会导致选择性地利用资源,从而使生态位宽度变窄;这种利用资源的能力从种群在群落中的分布范围和生物量的大小上得以体现(张金屯,2011)。一般地,在天然林中,一个种群必须具有较大的生态位宽度,较好地利用有限的资源,才能成为群落的优势种群。本研究发现,物种的生态位宽度与其重要值大小无关,但与其在各样方中的重要值均匀程度存在极显著的相关性,因而物种的生态位宽度实质反映的是该物种利用资源的均匀程度(即分布范围)。在判定某一物种对资源的利用情况时,不仅要看它的生态位宽度,还要考虑其实际的重要值大小(即生物量)。
大多数植物种群生态位研究的样方面积为20 m×20 m或者更大,边长20 m的正方形的对角线长为28.28 m,在这样的尺度下,两个物种即使同时出现在一个样方内,也未必会因为利用相同的资源而发生竞争。生态位重叠只是种间竞争发生的必要条件,而非充分条件,生态位重叠不一定就会发生种间竞争,当环境资源条件丰富时,它们可以共同享用资源,除非共享资源供应不足(张金屯,2011;金俊彦等,2013)。它们可通过其它方式如营养成份的选择性吸收、根系深浅的不同、相互错开物候期等,以此来减少种间竞争;在生态位分化的群落内,各种群在空间、时间和其它生态因子的利用等方面多趋向于互相弥补、互相配合,而不是直接竞争(杨持,2008)。生态位重叠值小可以表明竞争不激烈,但生态位重叠值较大并不一定能够表明竞争激烈。
致谢 丽水学院张超蓝等学生参与样地的外业调查,张莉芗等参与样地数据资料的整理与录入,温州大学丁炳扬教授在样地复查方面付出了大量的辛劳,在此一并致谢。
China National Biodiversity Research Group,1998.Chinese Nation-al Report of Diversity[M].Beijing:China Environment Science Press:147-154. [中国生物多样性国情研究组,1998.中国生物多样性国情研究报告[M].北京:中国环境科学出版社: 147-154.]
CHEN XR,CHEN YY,LUO ZR,et al,2013.A 5-year mid-mountain subtropical evergreen broadleaved forest study in Bais-hanzu,east China[J].J Zhejiang A&F Univ,30(6):821-829. [陈小荣,陈园园,骆争荣,等,2013.百山祖中山中亚热带常绿阔叶林群落5年动态特征[J].浙江农林大学学报,30(6):821-829.]
CHENG QB,WU MX,CHENG HT,1996.Comprehensive obser-vations report on Fengyangshan-Baizushan Nature Reserves of Zhejiang[J].J Zhejiang For Sci Technol,16(6):1-7. [程秋波,吴鸣翔,陈豪庭,1996.浙江凤阳山一百山祖自然保护区综合考察报告[J].浙江林业科技,16(6):1-7.]
CHENG ZQ,ZHANG KB,CHANG J,et al,2010.Vegetable niche of different enclosure measures in Yanchi County,Ningxia [J].Ecol Environ Sci,19(7):1 537-1 542. [程中秋,张克斌,常进,等,2010.宁夏盐池不同封育措施下的植物生态位研究[J].生态环境学报,19(7):1 537-1 542.]
DING S,OUYANG J,LU D,et al,2014.Niche characteristics of dominant species populations in Castanopsis fargesii forest in Jinggang Mountain National Nature Reserve[J].J Centr S Univ For&Technol,34(7):99-103.[丁松,欧阳杰,吕丹,等,2014.井冈山国家自然保护区栲树林优势种群生态位特征[J].中南林业科技大学学报,34(7):99-103.]
DOLéDEC S,CHESSEL D,GIMARET-CARPENTIER C,2000. Niche separation in community analsis:a new method[J].Ecol-ogy,81(10):2 914-2 927.
GAO JX,LU XZ,MA L,et al,2010.Study on the niche of the dominant tree species in the evergreen broad-leave forest in Fengyangshan[J].J Nanjing For Univ:Nat Sci Ed,34(4): 157-160.[高俊香,鲁小珍,马力,等.2010.凤阳山常绿阔叶林乔木层优势种群生态位分析[J].南京林业大学学报ű自然科学版,34(4):157-160.]
HE ZS,LIU JF,ZHU DH,et al,2012.Effects of forest gaps on species niche in regeneration layer of Castanopsis kawakamii nat-ural forest[J].Guihaia,32(5):624-629. [何中声,刘金福,朱德煌,等,2012.林窗对格氏栲天然林更新层物种生态位的影响[J].广西植物,32(5):624-629.]
HU ZH,QING HY,YU MJ,2009.The niche of dominant species populations in Castanopsis eyrei forest in Gutian Mountain National Nature Reserve[J].Acta Ecol Sin,29(7):3 670-3 677. [胡正华,钱海源,于明坚,2009.古田山国家级自然保护区甜槠林优势种群生态位[J].生态学报,29(7):3 670-3 677.]
HU YN,WANG DX,HUANG QP,et al,2012.Species composi-tion and population niche of natural forests in Zhouzhi Laox-iancheng Natural Reserve of Shaanxi[J].Acta Bot Boreal-Occi-dent Sin,32(6):1 238-1 244. [胡有宁,王得祥,黄青平,等,2012.陕西周至老县城自然保护区天然林物种组成及主要种群生态位[J].西北植物学报,32(6):1 238-1 244.]
JIN JY,QIN WG,LUO LJ,et al,2013.Research on the niche of dominant species population of endangered plants Kmeria septen-trionalis community[J].Guihaia,33(3):300-305. [金俊彦,覃文更,罗柳娟,等,2013.濒危植物单性木兰群落优势种群生态位研究[J].广西植物,33(3):300-305.]
JIN XF,DING BY,ZHENG CZ,et al,2004.The floristic analysis of seed plants in Baishanzu Nature Reserve from Zhejiang Prov-ince[J].Acta Bot Yunnan,26(6):605-618. [金孝锋,丁炳扬,郑朝宗,等,2004.浙江百山祖自然保护区种子植物区系分析[J].云南植物研究,26(6):605-618.]
LIU JF,HONG W,1999.A study on the community ecology of Castanopsis kawakamii:Study on the niche of the main tree pop-ulation Castanopsis kawakamii community[J].Acta Ecol Sin,19(3):347-352. [刘金福,洪伟,1999.格氏栲群落生态学研究——格氏栲林主要种群生态位的研究[J].生态学报,19(3):347-352.]
LIU W,CAO W,2011.Niche characteristics of main plant species in spruce-fir forests in Changbai Mountains[J].Chin J Ecol,30 (8):1 766-1 774. [刘巍,曹伟,2011.长白山云冷杉群落主要种群生态位特征[J].生态学杂志,30(8):1 766-1 774.]
PAOLI GD,CURRAN LM,ZAK DR,2006.Soil nutrients and beta diversity in the Bornean Dipterocarpaceae:evidence for niche par-titioning by tropical rain forest trees[J].J Ecol,94(1):157-170.
SHI XH,XU XB,ZHANG WH,2007.Study on the niche of the main populations in Abies chensiens community[J].Bull Bot Res,27(3):345-349. [史小华,许晓波,张文辉,2007.秦岭冷杉群落主要种群生态位研究[J].植物研究,27(3):345-349.]
SU ZY,WU DR,CHEN BG,2003.Niche characteristics of domi-nant populations in natural forest in North Guangdong[J].Chin J Appl Ecol,14(1):25-29. [苏志尧,吴大荣,陈北光,2003.粤北天然林优势种群生态位研究[J].应用生态学报,14(1):25-29.]
THOMAS MG,ROBEA KS,2003.Body size,niche breadth,eco-logically scaled responses to habitat fragmentation:mammalian predators in an agricultural landscape[J].Biol Cons,109(2): 283-295.
WANG W,LUO ZR,ZHOU RF,et al,2011.Habitat associations of woody plant species in Baishanzu subtropical broad-leaved ev-ergreen forest[J].Biodivers Sci,19(2):134-142. [王伟,骆争荣,周荣飞,等,2011.百山祖常绿阔叶林木本植物的生境相关性分析[J].生物多样性,19(2):134-142.]
WANG YS,XIE BY,WAN FH,et al,2008.Application of eco-logic niche models in explanation of niche shift of invasive alien species[J].Acta Ecol Sin,28(10):4 974-4 981. [王运生,谢丙炎,万方浩,等,2008.应用生态位模型研究外来入侵物种生态位漂移[J].生态学报,28(10):4 974-4 981.]
XU M,LUO ZR,YU MJ,et al,2007.Floristic composition and com-munity structure of mid-montane evergreen broad-leaved forest in north slope of Baishanzu Mountain[J].J Zhejiang Univ:Agric Life Sci Ed,33(4):450-457. [徐敏,骆争荣,于明坚,等,2007.百山祖北坡中山常绿阔叶林的物种组成和群落结构[J].浙江大学学报ű农业与生命科学版,33(4):450-457.]
YANG C,2008.Ecology[M].2nd Ed.Beijing:Higher Education Press:104-108. [杨持,2008.生态学[M].第2版.北京:高等教育出版社:104-108.]
YU JH,YAO FP,CHEN XR,et al,2003.An introduction to main vegetation types in the Baishanzu National Nature Reserve[J].J Trop Subtrop Bot,11(2):93-98. [余久华,姚丰平,陈小荣,等,2003.百山祖自然保护区主要植被类型概述[J].热带亚热带植物学报,11(2):93-98.]
ZHANG DY,JIANG XH,1997.A hypothesis for the origin and maintenance of within-community species diversity[J].Chin Biodivers,5(3):161-167. [张大勇,姜新华,1997.群落内物种多样性发生与维持的一个假说[J].生物多样性,5(3): 161-167.]
ZHANG GB,LI XQ,2007.Study on the niche of dominant species in evergreen broadleaved forest in Linnan Nature Reserve[J].J Nanjing For Univ:Nat Sci Ed,31(4):46-50. [张国斌,李秀芹,2007.岭南自然保护区常绿阔叶林优势树种的生态位研究[J].南京林业大学学报ű自然科学版,31(4):46-50.]
ZHANG GM,XIE SC,1997.Developement of niche concept and its perspectives:a review[J].Chin J Ecol,16(6):46-51.[张光明,谢寿昌,1997.生态位概念演变与展望[J].生志学杂志,16(6):46-51.]
ZHANG JT,2011.Quantitative Ecology[M].2nd Ed.Beijing: Science Press:113-115. [张金屯,2011.数量生态学[M].第2版.北京:科学出版社:113-115.]
ZHENG YR,1999.Main woody species niche of plant community in Daqinggou[J].J Plant Ecol,23(5):475-479. [郑元润,1999.大青沟森林植物群落主要木本植物生态位研究[J].植物生态学报,23(5):475-479.]
ZHU CQ,LEI JP,1997.Expansion of human niche and sustainable development[J].Chin J Ecol,16(3):50-54. [朱春全,雷静品,1997.人类生态位的扩充与可持续发展[J].生志学杂志,16(3):50-54.]
Niche of dominant species populations in an evergreen broad-leaved forest in Baishanzu
WU You-Gui1∗,YE Zhen-Lin1,ZHOU Rong-Fei1,YANG Hui2,LUO Zheng-Rong3
(1.Management Office of Baishanzu,Fengyangshan-Baishanzu National Nature Reserve,Qingyuan 323800,China;2.School of Forestry and Biotechnology,Zhejiang Agriculture and Forestry University,Lin’an 311300,China;3.College of Ecology,Lishui University,Lishui 323800,China)
Dominant plant populations have great ability to adapt to the environment and use resources,and they play a decisive role in the community structure and function.What's more,the species niche characteristic reflects the utiliza-tion of environmental resources from a perspective.In order to explore the relationship,position and function of each dominant species population in an evergreen broad-leaved forest of Baishanzu,nine subplots of 20 m×20 m regularly from the 5 hm2(250 m×200 m)plot which established in 2003 were selected,where all trees with DBH≥1 cm were tagged,measured,and identified to species.The niche breadths,niche similarities and niche overlaps of 13 species (importance value larger than 1 )were analyzed quantitatively based on the data collected in 2013.The results demon-strated that the niche breadths of Cyclobalanopsis multinervis,Rhododendron latoucheae and Eurya rubiginosa var.attenu-ate were relatively larger,with Hurlbert niche breadth values(Ba)being 0.878,0.825,0.806,respectively,while the niche breadths of Cleyera pachyphylla,Fagus lucida and Sycopsis sinensis were relatively smaller,with Ba values being 0.341,0.320,0.272,respectively.The species niche breadth was not related to its importance value,but was signifi-cantly positively correlated with the evenness of importance values across different plots.The niche similarity between Cyclobalanopsis multinervis and Camellia cuspidate was the largest,being 0.832,while that between Ilex editicostata and Sycopsis sinensis was the smallest,being 0.248.Despite of small niche breadths,in addition,the niche similarity be-tween Cleyera pachyphylla and Fagus lucida was high(0.758)due to their similar habitat requirements.Low niche simi-larities were found in two pair of species in the same genus:between Cleyera multinervis and C.stewardiana,and be-tween Ilex formosana and I.editicostata.Species with wide niche breadth often had high degrees of niche overlap with other species,and large niche similarity was often associated with high degrees of niche overlap.Low degrees of niche overlap were found between Cleyera multinervis and C.stewardiana,and between Ilex formosana and I.editicostata.In a word,low degrees of niche overlap found in this study suggested weak interspecies competition among dominant species and therefore a potentially stable plant community in this forest.This paper revealed the niche characteristics of dominant species populations and structural stability of the community,and it would provide theoretical basis for the research of biodiversity conservation,succession,population evolution.And it also could provide reference for the nature reserve management institution to adjust and improve conservation measures.
evergreen broad-leaved forest,dominant species populations,niche breadth,niche similarity,niche over-lap,Baishanzu Mountain
Q948.1,S718.54
A
1000-3142(2016)02-0186-07
10.11931/guihaia.gxzw201408035
吴友贵,叶珍林,周荣飞,等.百山祖常绿阔叶林优势种群的生态位[J].广西植物,2016,36(2):186-192
WU YG,YE ZL,ZHOU RF,et al.Niche of dominant species populations in an evergreen broad-leaved forest in Baishanzu[J].Guihaia,2016,36(2):186-192
2014-08-29
2015-03-30
国家林业局林业国家级自然保护区补助资金项目(财农[2013]81号)[Supported by Grant Fund for National Natural Reserve of State Forestry Administration([2013]81)]。
吴友贵(1970-),男,浙江庆元县人,高级工程师,主要从事自然保护区生物多样性保护等研究,(E-mail)qywyg@sina.com
