生境异质性对鼎湖山常绿阔叶林群落功能多样性的影响
2016-05-09周光霞黄立新臧晓蔚叶万辉
周光霞,黄立新,臧晓蔚,韦 霄,叶万辉,沈 浩∗
(1.中国科学院退化生态系统植被恢复与管理重点实验室,中国科学院华南植物园,广州510650;2.中国科学院大学,北京100049;3.广西壮族自治区广西植物研究所,广西桂林541006)中国科学院
生境异质性对鼎湖山常绿阔叶林群落功能多样性的影响
周光霞1,2,黄立新1,2,臧晓蔚1,2,韦 霄3,叶万辉1,沈 浩1∗
(1.中国科学院退化生态系统植被恢复与管理重点实验室,中国科学院华南植物园,广州510650;2.中国科学院大学,北京100049;3.广西壮族自治区广西植物研究所,广西桂林541006)
中国科学院
物种共存机制一直以来是群落生态学的研究热点。为了探讨异质生境条件下鼎湖山常绿阔叶林群落功能多样性变化,找到其变化的主要环境驱动因子,该研究利用位于鼎湖山20 hm2监测样地第2次群落调查数据并选择代表不同生境(海拔和地形)的27个样方(20 m×20 m),于2013年夏季在样地内所选样方中测定所有胸径≥1 cm树种的叶片功能性状。所测性状包括形态学性状(比叶面积、叶片干物质含量、叶面积以及叶片长宽比)和化学计量学性状(叶片碳、氮、磷的含量),结合地形和土壤数据并通过分析功能多样性随环境梯度的变化,探讨了环境过滤和竞争在鼎湖山群落物种共存中的相对重要性。结果表明:功能分歧度和群落权重平均值与环境因素关系密切,尤其是海拔、凹凸度和土壤养分。环境条件较好区域(微尺度高海拔、高凹凸度和土壤养分含量)的植物采取统一的养分有效保存(低SLA,高LDMC)的适应策略(功能分歧度低),环境过滤所起作用更强;植物在相反的环境条件下,采取快速生长策略(高SLA,低LDMC),能够更好地适应环境的变化,且性状变化是多样的(功能分歧度高),在该条件下竞争作用更为显著。叶面积和叶片氮含量的分歧度在环境条件较好的区域增大,这与其他功能性状不一致,说明不同生态位轴(环境因素)影响不同性状的分歧度变化,并且在局域尺度上植物为了更好地适应环境变化采取了多样的适应策略。
群落生态学,功能多样性,生境异质性,群落共存机制,适应性策略
群落构建机制(community assembly mechanisms)一直以来是生态学研究的热点问题,其中环境过滤(environmental filtering)和竞争(competitive interac-tions)是影响群落构建最主要的两种生态学过程(Cornwell et al,2006;Kraft et al,2008;Siefert et al,2013)。一般认为,环境过滤会导致物种的适应性策略趋同,而竞争则导致物种的适应性策略趋异(Spasojevic&Suding,2012;Chalmandrier et al,2013;Purschke et al,2013)。功能多样性(functional diversity)是指功能性状在某个特定群落内的值、范围和相对多度(Villeger et al,2008;Car-reno-Rocabado et al,2012)。功能多样性与群落构建机制和生态系统过程都有着密切的关联(Mason et al,2012;Conti&Diaz,2013;Mason et al,2013;Roscher et al,2013)。环境过滤和竞争的相对强度通常导致不同的群落构建方式(Spasojevic&Suding,2012),即功能趋同或者趋异(Kraft et al,2008),两者的相对强度可能取决于群落的环境因素,并随环境梯度而变化。目前针对功能多样性与环境因子的相关性研究,包括功能性状与大尺度条件下土壤和气候的变异有显著相关性(Siefert et al, 2013;Fortunel et al,2014);随着群落演替的进行,功能多样性会显著升高(Janecek et al,2013;Pur-schke et al,2013)。然而,小尺度下功能多样性随环境梯度变化尚不清楚(Lohbeck et al,2012)。
植物功能多样性的测量方法多样,但各个指标都是在植物功能性状和物种丰富度的基础上进行的,采用较多的有Rao指数(Rao,1982)、功能丰富度(Mason et al,2005)、功能均匀度(Mouillot et al,2005)、群落权重平均值(Lavorel et al,2008)等。其中受到关注较多的是功能离散度(functional disper-sion,Fdis)和群落权重平均值(communtiy weighted mean,CWM),群落离散度是指随机挑选的物种间某一相同性状值之间的差异,通常被用来表征生态位互补性(Petchey&Gaston,2006)。群落权重平均值是指物种的功能性状值加权相对多度后表征群落中该性状的平均值,通常被用来评估群落的动态以及生态系统特性(Enoki&Abe,2004;Louault et al,2005)。从功能性状的群落权重平均值和功能离散度的双重角度出发,能够更加准确地判断随着环境变化,群落水平的功能性状以及性状离散度水平的变化趋势。
同一群落中从性状的不同角度出发会出现一系列不同的功能多样性指标(Ricotta,2005;Petchey &Gaston,2006)。由于不同功能性状可能与不同生态过程相关,当不同的单一性状与生态位轴的关系相反时,这些相反的关系有可能造成在多元功能多样性的计算中相互抵消,从而掩饰多元功能多样性与环境的关联。某些群落的多元功能多样性与环境梯度之间无显著关联(Cornwell et al,2006),这会极大地影响群落构建机制的判断(Cornwell&Ack-erly,2009;Spasojevic&Suding,2012)。所以本文从植物叶片形态学性状和理化性质出发,选择具有代表性的功能性状来探索功能多样性的变化。
森林群落中,生境异质性通常是小尺度变异引起的(如地形和土壤的变化) (Harms et al,2001;John et al,2007),生境异质性导致样地能量、水分和养分的可利用性产生差异(Bellingham et al,2000),从而对群落中物种的分布产生直接或间接影响。鼎湖山大样地作为中国森林生物多样性监测网络(Chinese Forest Biodiversity Monitoring Network)和美国Smithsonian研究院CTFS(Center for Tropical Forest Science)最重要的大型监测样地之一,建成于2005年(叶万辉等,2008),2010年完成了首次复查(Shen et al,2013)。它作为南亚热带常绿阔叶林的典型代表,海拔230~470 m,地形复杂,存在较强的环境异质性(叶万辉等,2008)。关于鼎湖山大样地群落构建机制,已在群落(Li et al,2009;Wang et al,2009)、种群遗传(Wang et al,2012)、进化(Pei et al,2011)等方面开展了研究。生境异质性、种子性状个体间的相互作用等对物种分布格局的形成以及生物多样性的维持起着重要作用,而功能多样性的变化正是植物对生境异质性的适应性体现,在群落构建中起到非常重要的作用。因此,本研究从海拔和地形的角度,探讨了鼎湖山样地异质生境条件下群落功能多样性的变化,为物种共存的重要生态学过程提供理论支持,同时也为该地区植被恢复以及生态系统的保护提供理论依据。
1 材料与方法
1.1 研究区自然概况
鼎湖山国家级自然保护区位于广东省肇庆市,112°30′39″~112°33′41″E,23°09′21″~23°11′30″N,地处亚热带季风气候区南缘,北回归线附近,形成并保存了较完好的地带性植被类型——南亚热带常绿阔叶林,是我国植物种类最丰富的植被类型之一,具有热带向亚热带的过渡性质,群落结构复杂,物种组成丰富。年平均温度为20.9℃,年平均降雨量为1 927 mm,4-9月是主要的降雨季节,也是植物生长的主要季节,鼎湖山的地带性土壤为赤红壤。
鼎湖山大样地位于鼎湖山保护区的核心区,地形复杂,海拔为230~470 m,坡度为30°~50°,地形起伏较大,水热条件良好,相对湿度较大。植被类型为南亚热带常绿阔叶林,群落的发育和保存良好(叶万辉等,2008)。
1.2 研究方法
本研究中,生境异质性以海拔和地形因子加以表征。在鼎湖山大样地中选择具有代表性的3条山体,从同一山体的不同海拔高度(308.42~413.4 m)、不同凹凸度(-6.16~6.71 m)出发,于每条山体不同海拔(低海拔、中海拔、高海拔)和不同凹凸度的样方(山谷、山坡、山脊)中选取Jacard相似度最高的样方(20 m×20 m)作为本次研究对象,每条山脉同一海拔同一凹凸度选择3个样方,共包括27个样方,代表9种生境类型(表1)。
1.2.1 环境因子测定 地形参数选取海拔高度、凹凸度、坡度和坡向;土壤参数选取土壤含水量、容重、pH、土壤有机质以及土壤中钾、磷、氮的含量,数据来源于鼎湖山20 hm2样地的测定数据。地形因子的测定是测量每个20 m×20 m样方4个角的海拔均值为此样方的海拔;凹凸度为某个样方的海拔减去相邻8个样方的海拔平均值;坡度为样方的任意3个角形成4个三角形平面偏离水平面角度的均值。土壤因子测定时在20 hm2样地内每隔30 m系统随机布设土壤采样点,在每个采样点中,随机选择8个方向中的一个方向,在2、5或15 m三个距离中随机增加2个采样点。在鼎湖山共布设了714个采样点(Lin et al,2013)。取表层0~10 cm的表土用于测定土壤指标。据此,本研究每个样方涉及到的样点为2~3个,共统计了64个样点的土壤数据。
1.2.2 功能性状的测定 对选定样方中所有可以采到的DBH≥1 cm植株,多于10株的物种随机采10株作为代表,不足10株物种则全部采集。每植株分别选取4~6片向阳成熟的叶片,先称量叶片鲜重(fresh weight,FW),再用螺旋测微器测量植物的叶片厚度(leaf thickness,LT),用LI-3000C便携式叶面积仪(Li-Cor,NE,USA)测量叶片面积(leaf area,LA)以及叶片长度(leaf length,LL)、叶片宽度(leaf width,LW),最后将叶片放入70℃的烘箱中烘干至恒重,并称量其干重(dry weight,DW)。比叶面积(specific leaf area,SLA)为叶面积与其干重的比值;叶片干物质含量(leaf dry matter content,LDMC)为叶片干重与鲜重的比值。样品烘干后磨碎过100目筛,用钼抗比色法测定单位质量叶片磷含量;碳氮含量用元素分析仪(Elementar Analysensysteme GmbH,Hanau,Germany)测定。
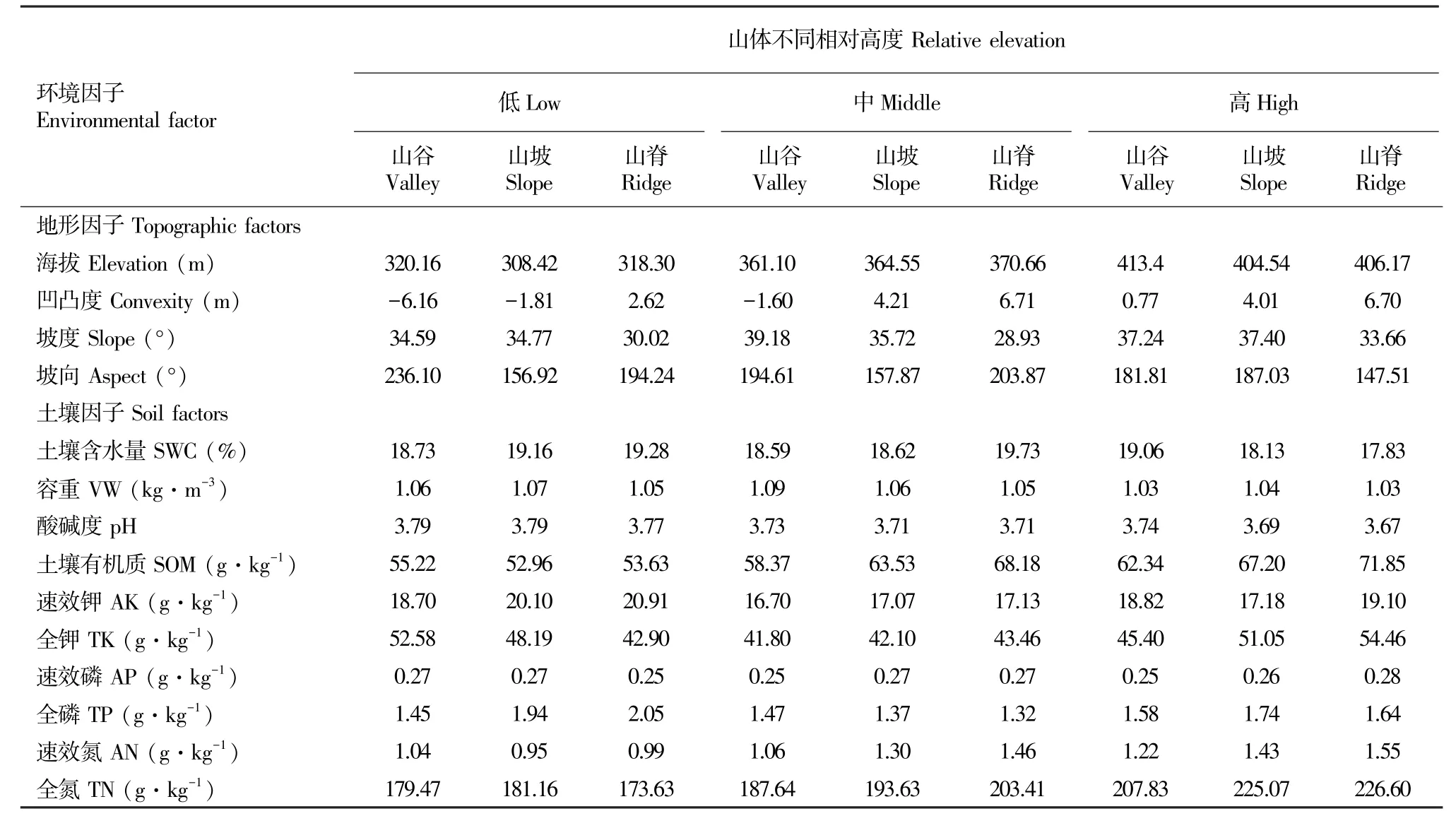
表1 鼎湖山样地异质生境条件下环境因子指标Table 1 Environmental factors of heterogeneous habitats in Dinghu Mountain plot
本文选取功能离散度(Functional dispersion,FDis)和群落的权重平均值(Communtiy-Weighted Mean,CWM)这两种功能多样性来解释群落构建机制,其计算方法参考Spasojevic et al(2012)。
1.3 数据分析
所有相关性分析均采用软件SPSS 18.0(SPSS Inc.Chicago,IL,USA)进行统计分析,当P<0.05则表示有统计学意义。功能离散度(Fdis)和群落权重平均值(CWM)等功能多样性参数的计算采用R语言(3.0.1)(Team,2012),调用R语言包“FD”完成(Laliberte&Legendre,2010)。采用CANOCO 4.5 对Fdis和CWM与环境因子的关系进行多元典范冗余分析(Redundancy analysis,RDA)。
2 结果与分析
2.1 异质生境中的环境因子
在鼎湖山样地中,从所选取的样方的环境因子中可以看出,低海拔、中海拔与高海拔之间均有50 m的微尺度高度差,凹凸度(-6.16~6.71 m),分别代表了山谷、山坡和山脊的生境。样地的坡度无明显变化,所选样地均向南(表1)。在土壤因子中,总磷和总氮以及土壤有机质含量变化较大,其他土壤因子变幅不大。表2表明,与土壤因子显著相关的是海拔高度和凹凸度,高海拔和凹凸度较大的样方中,其土壤有机质、有效氮和总氮含量也越高,而土壤pH值越低,高海拔地区的土壤养分更为充足。
2.2 样地群落功能分歧度(FDis)
不同性状在异质生境中分歧度的变异趋势是不同的,其中叶片厚度,比叶面积,叶片磷含量以及叶片中的碳磷比和碳氮比在同一海拔下的变化趋势都是山谷>山坡>山脊;而叶片碳含量,叶片磷含量以及叶片氮磷比在同一海拔的不同生境下都是先上升后下降的趋势。不同海拔同一地形下性状的多样性无明显差异(表3)。

表2 鼎湖山样地27个样方地形因子与土壤因子之间的相关性分析Table 2 Correlation coefficients between soil characteristics and topological factors in the 27 sampled quadrats in Dinghu Mountain plot
2.3 样地地形和土壤因子与功能分歧度
鼎湖山样地群落功能分歧度与地形因子以及土壤因子的相关性分析(表4)结果显示,随着海拔升高,叶片干物质含量的分歧度变化降低,性状趋于一致,凹凸度的增加也会引起比叶面积、叶片干物质含量、叶片磷含量的分歧度显著降低,坡向与叶面积的分歧度变化是正相关。
土壤因子与不同功能性状分歧度变化的相关性结果显示,土壤pH的升高会引起叶片干物质含量及叶片磷含量的分歧度增加;土壤有机质含量的增加则会引起叶片干物质含量的分歧度变化降低;土壤中有效氮和总氮含量增加,叶片干物质含量的分歧度变化降低,但叶片氮含量的分歧度变化增加。
对群落功能分歧度与地形因子以及土壤因子的RDA约束排序结果见图1。凹凸度对功能分歧度的解释力最大(图1:A),与叶片干物质含量以及叶片磷含量的分歧度变化呈负相关,海拔与叶片氮含量的分歧度变化呈正相关;从图1:B看出,土壤pH、容重、有机质含量和氮素含量对功能分歧度的影响较大。土壤pH、容重与有机质、有效氮和总氮含量与不同功能分歧度的变化相反,前者与叶片厚度,叶片干物质含量以及比叶面积的分歧度变化都呈正相关,而后者与这些性状的分歧度变化为负相关。
2.4 样地叶片功能性状群落权重平均值(CWM)
群落权重平均值的结果综合分析了植物的功能性状及其在该群落中的多度,能充分表明性状在群落间的变化。从鼎湖山各群落权重平均值的结果中看出,随着海拔升高,叶片厚度、比叶面积以及叶面积的群落权重平均值均减小,叶片干物质含量,叶片碳含量的群落权重平均值均增加,而同一海拔不同地形下不同性状之间几乎无差异(表5)。
2.5 样地地形和土壤因子与群落权重平均值的关系
CWM与环境因子的相关性分析结果显示(表6),影响群落中不同性状的CWM主要是海拔和凹凸度,且海拔和凹凸度对不同性状的群落权重平均值的影响是一致的,随着海拔和凹凸度的增加,叶片厚度、比叶面积、叶面积的群落权重平均值均降低,而叶片干物质含量、叶片的长宽比以及叶片碳、氮含量的群落权重平均值均增加。影响群落权重平均值的主要土壤因子是土壤容重、pH、有机质以及有效氮和总氮含量。其中土壤容重和pH对群落权重平均值的影响是一致的,随着土壤容重和pH的增加,比叶面积和叶面积的群落权重平均值均增加,而叶片干物质含量以及叶片碳含量的群落权重平均值均降低;土壤有机质含量和土壤有效氮以及总氮含量对性状群落权重平均值的影响与前述的土壤容重和pH则相反,随着土壤有机质含量和土壤有效氮以及总氮含量的增加,比叶面积、叶面积的群落权重平均值均降低,而叶片干物质含量和叶片碳含量的群落权重平均值均增加。
群落权重平均值与地形因子以及土壤因子的RDA约束排序结果见图2。从图2:A可以看出,海拔高度和凹凸度与叶面积、叶片厚度以及比叶面积的群落权重平均值是负相关的,与叶片干物质含量以及叶片碳含量的群落权重平均值则是正相关的。图2:B中,土壤容重和pH对性状权重平均值的影响与土壤有机质和土壤氮含量的影响恰恰是相反的,土壤容重和pH与叶面积以及比叶面积的群落权重平均值是正相关的,土壤有机质和土壤氮含量与其则为负相关;而土壤有机质和土壤氮含量与叶片干物质含量以及叶片碳含量的群落权重平均值是正相关的,则土壤容重和pH与其是负相关的。
3 讨论
功能多样性越来越多地被用来检验环境过滤和竞争排斥导致的群落物种共存(Mouchet et al,2010;Mason et al,2013;Siefert et al,2013)。一般认为,在环境过滤起主要作用的条件下功能分歧度会减小,而在竞争排斥较为明显的条件下功能分歧度会增加(Cornwell&Ackerly,2009)。本研究结果表明,除叶面积和叶片氮含量外,其他功能性状在共存的物种中呈现出在高海拔,凹凸度较大以及土壤养分较高的环境条件下表现出功能趋同,环境过滤的作用更明显,且在该资源相对丰富的条件下群落权重平均值中出现了低SLA,高LDMC,这是一种有效的养分保存策略。鼎湖山资源相对匮乏的条件下(低海拔,凹凸度较小,土壤养分较低),由于竞争作用加强,功能分歧度增加,且在该条件下快速生长(高SLA,低LDMC)的生存策略更容易在竞争中取胜。本研究还发现,叶面积和叶片氮含量的变化与其他性状是不一致的,在Spasojevic&Suding (2012)的研究中也出现了两种性状(叶面积和树高)与其他性状(比叶面积、气孔导度、叶绿素含量)的分歧度变化不一致的结果,说明植物在局域环境下采取了多样的适应策略,且不同性状的变化有差异。

表3 鼎湖山样地异质生境下各群落功能分歧度指标Table 3 Functional dispersion of different communities in heterogeneous environments in Dinghu Mountain plot
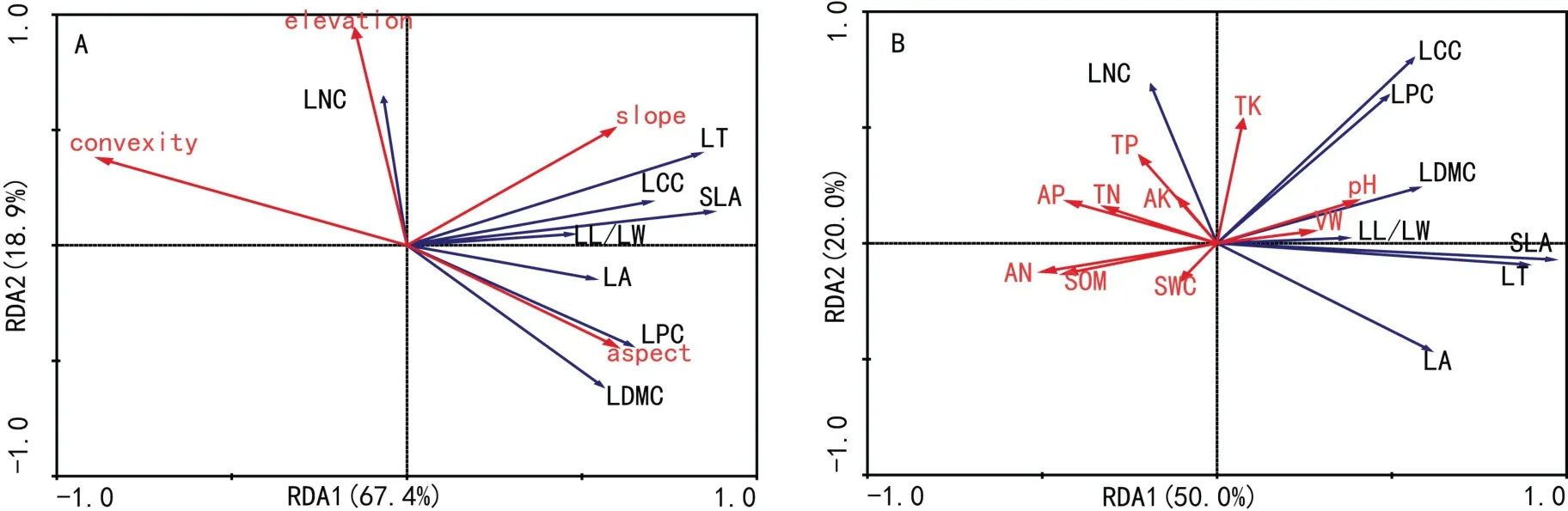
图1 鼎湖山样地环境因子对群落叶片功能分歧度影响的冗余分析Fig.1 RDA analysis on the relationship between environment factors and leaf functional dispersion in Dinghu Mountain plot

图2 鼎湖山样地环境因子对群落权重平均值影响的冗余分析Fig.2 RDA analysis on the relationship between environment factors and community-weighted mean in Dinghu Mountain plot
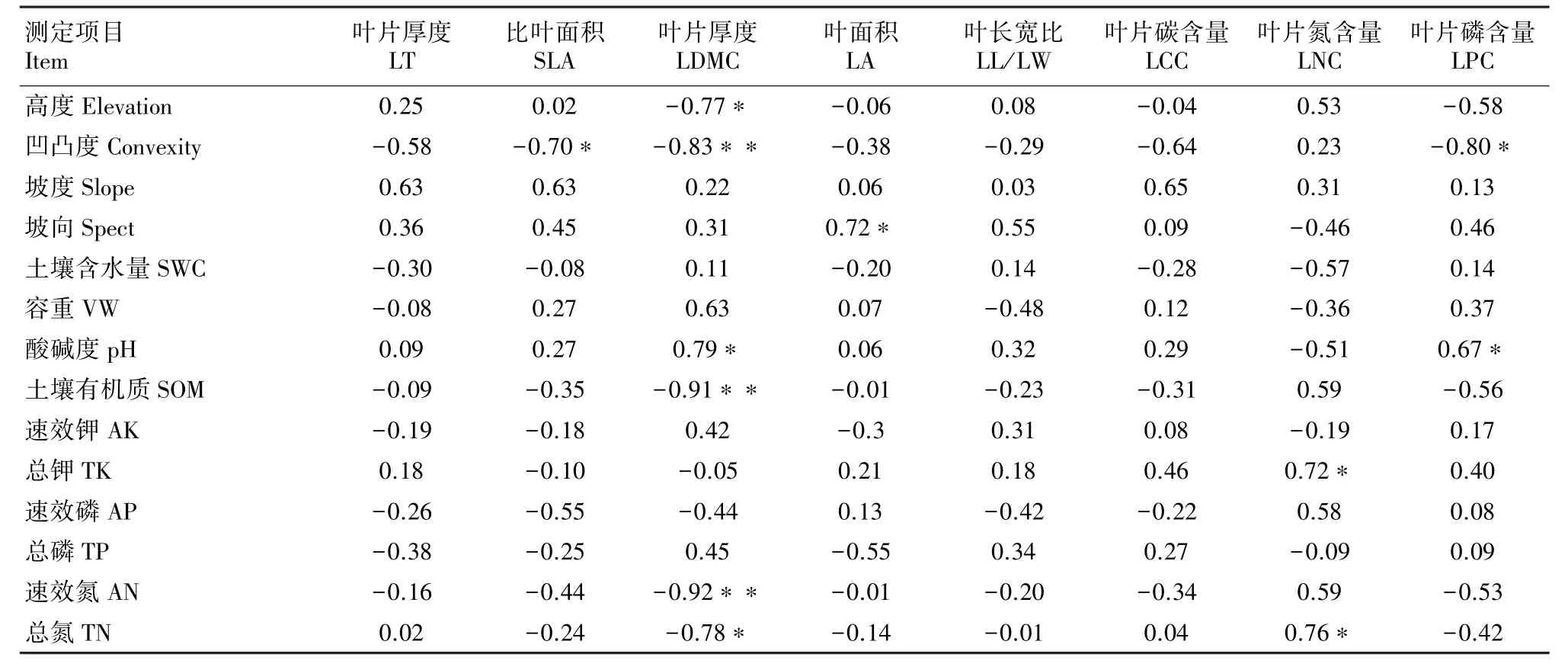
表4 鼎湖山样地群落功能分歧度与环境因子之间的相关系数Table 4 Correlation coefficients between plant functional dispersion and environmental factors in Dinghu Mountain plot

表5 鼎湖山样地异质生境下各群落权重平均值指标Table 5 Community-weighted mean of different communities in heterogeneous habitats in Dinghu Mountain plot
3.1 功能分歧度与环境因子的相关性
目前,在大尺度条件下,环境因素与功能分歧度之间存在密切关联(Cornwell&Ackerly,2009;Spa-sojevic et al,2012;Siefert et al,2013;Fortunel et al,2014),但在局域尺度下是未知的。
通过鼎湖山地形与土壤因子的相关性分析可以看出,海拔和凹凸度与土壤有机质,有效氮含量和总氮含量是负相关的,一些研究发现凹凸度与土壤养分有负相关关系(Noguchi et al,2007;Lan et al,2011;McEwan et al,2011),因而可以对植物性状起到影响作用。在鼎湖山样地中海拔高差不很明显,但高凹凸度与高光照相关联(Enoki&Abe,2004;Tsujino et al,2006)。此外,从低海拔到高海拔,人类活动强度会减弱(del Moral&Watson,1978),加上微生物活动会增加土壤有机质含量(隋跃宇等,2009)。因此,在鼎湖山高海拔、凹凸度较大的样地中,土壤养分较低海拔、凹凸度较小的样地更为丰富。

表6 鼎湖山样地群落权重平均值与环境因子之间的相关系数Table 6 Correlation coefficients between community-weighted mean and environmental factors in Dinghu Mountain plot
在热带森林中,植物对土壤养分和光照的竞争较强,生长在高凹凸度区域的个体能够获得更多的光照(Enoki&Abe,2004;Tsujino et al,2006)。鼎湖山样地植物功能性状中叶面积和叶片氮含量的分歧度是增加的,表明随着样地的海拔和凹凸度增加,光照强度将发生变化(Song et al,2006;Wang et al,2008;Chu et al,2009;Mason et al,2011),植物通过形成不同的叶面积来适应对光照的充分利用,而叶片氮含量的多样性变化是因为植物在条件较好的情况下竞争地下资源(Spasojevic&Suding,2012),所以在鼎湖山高海拔叶面积和叶片氮含量的分歧度都是增加的。而其他性状的分歧度变化在条件较好的条件下(高海拔、凹凸度较大)是降低的,即植物采取了趋同的性状来适应环境的变化,这是环境过滤作用更为明显的结果。这些性状在资源相对匮乏的条件下(低海拔、凹凸度较小)分歧度是增加的,这是由于在环境压力较大的条件下,竞争排斥更为显著(Tilman,1982),所以出现了性状分歧度在贫瘠的条件下增加。
功能分歧度的格局与土壤因子也有明显的关联,土壤肥力(如土壤K和P的含量)能够影响功能分歧度(Mason et al,2012),单一性状功能分歧度比如树高、比叶面积和叶绿素含量也与土壤含水量和N含量有显著的关联(Spasojevic&Suding,2012)。在鼎湖山样地中,叶片干物质含量的分歧度变化受到土壤因子的影响较大,土壤中有机质、有效氮和总氮含量越高的样地其pH低,这种情况下,叶片干物质含量的分歧度变化降低,植物采取了统一的养分保存的适应策略。
3.2 CWM与环境因子的相关性
CWM随着环境梯度发生变化是环境过滤在起作用,但这种过滤作用是植物对环境的抵抗能力,另一种适应方式是植物通过竞争资源,结果是适合的性状或性状组合得以保留(Cornwell&Ackerly,2009;Lebrija-Trejos et al,2010)。
从本研究分析结果来看,高海拔和高凹凸度条件下,即土壤中有机质含量和有效氮以及总氮含量较高,土壤容重和pH较低的条件下,比叶面积和叶面积的群落权重平均值都是降低的,而叶片干物质含量和叶片碳含量的群落权重平均值都是增加的。高的凹凸度和土壤肥力更适合有大的叶面积和比叶面积,这与物种具有更高的生长速率和碳同化速率(Poorter et al,2008;Carreno-Rocabado et al,2012),但鼎湖山与这些研究的结果是不一致的,在高凹凸度和高土壤肥力条件下,比叶面积和叶面积都是减小的,而叶片干物质含量和叶片碳含量都是增加的。SLA和LDMC反映了植物性状中生物量的快速生产(高SLA,低LDMC物种)和养分的有效保存(低SLA,高LDMC物种)的一个基本权衡(Garnier,2001)。在资源丰富的条件下,由于环境过滤的作用,植物统一采取了快速生长的策略(树高,叶面积和叶绿素含量都是增大的)(Spasojevic et al,2012),而在鼎湖山样地中出现了叶片干物质含量和叶片碳含量增加,这说明植物采取了养分的有效保存。结合其他性状的变化如叶片碳含量增大、叶面积减小可以看出,在鼎湖山生长环境优越条件下,植物是通过延长其生命、提高叶片碳含量,来为提高其生命周期打下基础,而不是通过短期生物量的快速生产策略,物种采取这种生活方式也是为了适应小尺度环境的不断变化,这也是与许多发现不一致的地方(Carreno-Rocabado,2012)。主要是因为鼎湖山环境变化多样,采取有效的养分保存策略会提高其存活的几率。所以在鼎湖山样地中条件较为优越的条件下,CWM的变化是环境过滤的作用,即保留有效保存养分的植物。在资源较为贫瘠的条件下,植物所需资源紧缺,竞争作用明显(Cornwell et al,2009),能够快速生长的植物被保留下来,结合分歧度的变化来看,这种快速生长的生活策略的分歧度变化相对营养丰富的条件更高,但统一是高SLA,低LDMC的适应策略。
4 结论
一般认为,环境压力较大的环境生境过滤作用更明显,功能趋同(Cornwell et al,2006),而在资源丰富的环境条件下,竞争排斥作用会引起功能趋异(Chesson,2000)。但本研究结果恰恰相反,在微尺度低海拔条件下,资源相对微尺度高海拔减少,但是植物为了生存,竞争作用明显,功能多样性增大,结合CWM的结果来看,这些竞争力强的植物都是能够快速生长(高SLA,低LDMC)的植物。而在高海拔条件相对优越的环境条件下,植物采取了统一的养分有效保存的生存策略,但叶面积和叶片氮含量的多样性变化由于光照和土壤养分的作用,在高海拔的分歧度变化是增大的。所以说,植物在局域尺度下,为了更好地适应环境的变化采取了多样的适应策略,且不同性状的变化有差异,进一步论证了采取多元功能多样性可能存在的问题。
局域尺度上物种共存不同于大尺度上的一般规律,理解局域尺度上物种的共存机制较大尺度更为复杂。鼎湖山样地特殊的异质生境变化为物种共存的探讨提供了一个崭新的平台,同时也为局域尺度下功能多样性变化提供理论基础。
BELLINGHAM PJ,TANNER EVJ,2000.The influence of topogra-phy on tree growth,mortality,and recruitment in a tropical montane forest[J].Biotropica,32(3):378-384.
CARRENO-ROCABADO G,PENA-CLAROS M,BONGERS F,et al,2012.Effects of disturbance intensity on species and functional diversity in a tropical forest[J].J Ecol,100(6): 1 453-1 463.
CHALMANDRIER L,MUNKEMULLER T,GALLIEN L,et al,2013.A family of null models to distinguish between environ-mental filtering and biotic interactions in functional diversity pat-terns[J].J Veg Sci,24(5):853-864.
CHESSON P,2000.Mechanisms of maintenance of species diversity [J].Ann Rev Ecol Syst,31:343-366.
CHU CJ,WANG YS,LI Q,et al,2009.Effects of traits,species identity and local environmental conditions on the assessment of interactions:insights from an alpine meadow community[J].J Plant Ecol,2:135-141.
CONTI G,DIAZ S,2013.Plant functional diversity and carbon storage-an empirical test in semi-arid forest ecosystems[J].J Ecol,101(1):18-28.
CORNWELL WK,ACKERLY DD,2009.Community assembly and shifts in plant trait distributions across an environmental gradient in coastal California[J].Ecol Monogr,79(1):109-126.
CORNWELL WK,SCHWILK DW,ACKERLY DD,2006.A trait-based test for habitat filtering:convex hull volume[J]. Ecology,87(6):1 465-1 471.
DEL MORAL R,WATSON AF,1978.Gradient structure of forest vegetation in the central Washington Cascades[J].Vegetatio,38(1):29-48.
ENOKI T,ABE A,2004.Saplings distribution in relation to topog-raphy and canopy openness in an evergreen broad-leaved forest [J].Plant Ecol,173(2):283-291.
FORTUNEL C,PAINE CET,FINE PVA,et al,2014.Environ-mental factors predict community functional composition in Ama-zonian forests[J].J Ecol,102(1):145-155.
GARNIER E,SHIPLEY B,ROUMET C,et al,2001.A standard-ized protocol for the determination of specific leaf area and leaf dry matter content[J].Funct Ecol,15(5):688-695.
HARMS KE,CONDIT R,HUBBELL SP,et al,2001.Habitat as-sociations of trees and shrubs in a 50 ha neotropical forest plot [J].J Ecol,89(6):947-959.
JOHN R,DALLING JW,HARMS KE,et al,2007.Soil nutrients influence spatial distributions of tropical tree species[J]. PNAS,104(3):864-869.
JANECEK S,DE BELLO F,HORNIK J,et al,2013.Effects of land-use changes on plant functional and taxonomic diversity along a productivity gradient in wet meadows[J].J Veg Sci,24 (5):898-909.
KRAFT NJB,VALENCIA R,ACKERLY DD,2008.Functional traits and niche-based tree community assembly in an amazonian forest[J].Science,322(5901):580-582.
LALIBERTE E,LEGENDRE P,2010.A distance-based framework for measuring functional diversity from multiple traits[J].Ecol-ogy,91(1):299-305.
LAN GY,HU YH,CAO M,et al,2011.Topography related spatial distribution of dominant tree species in a tropical seasonal rain forest in China[J].For Ecol Manag,262(8):1 507-1 513.
LAVOREL S,GRIGULIS K,MCINTYRE S,et al,2008.Assessing functional diversity in the field-methodology matters! [J].Funct Ecol,22(1):134-147.
LEBRIJA-TREJOS E,PEREZ-GARCIA EA,MEAVE JA,et al,2010.Functional traits and environmental filtering drive commu-nity assembly in a species-rich tropical system[J].Ecology,91 (2):386-398.
LI L,HUANG ZL,YE WH,et al,2009.Spatial distributions of tree species in a subtropical forest of China[J].Oikos,118 (4):495-502.
LIN G,STRALBERG D,GONG G,et al,2013.Separating the effects of environment and space on tree species distribution: from population to community[J].PLos One,8:e56171.
LOHBECK M,POORTER L,PAZ H,et al,2012.Functional di-versity changes during tropical forest succession[J].Perspect Plant Ecol,14(2):89-96.
LOUAULT F,PILLAR VD,AUFRERE J,et al,2005.Plant traits and functional types in response to reduced disturbance in a semi-natural grassland[J].J Veg Sci,16(2):151-160.
MASON NWH,DE BELLO F,DOLEZAL J,et al,2011.Niche o-verlap reveals the effects of competition,disturbance and con-trasting assembly processes in experimental grassland communities[J].J Ecol,99(3):788-796.
MASON NWH,DE BELLO F,MOUILLOT D,et al,2013.A guide for using functional diversity indices to reveal changes in assembly processes along ecological gradients[J].J Veg Sci,24 (5):794-806.
MASON NWH,RICHARDSON SJ,PELTZER DA,et al,2012.Changes in coexistence mechanisms along a long-term soil chronosequence revealed by functional trait diversity[J].J Ecol,100(3):678-689.
MCEWAN RW,LIN YC,SUN IF,et al,2011.Topographic and bi-otic regulation of aboveground carbon storage in subtropical broad-leaved forests of Taiwan[J].For Ecol Manag,262(9): 1 817-1 825.
MOUCHET MA,VILLEGER S,MASON NWH,et al,2010.Func-tional diversity measures:an overview of their redundancy and their ability to discriminate community assembly rules[J]. Funct Ecol,24(4):867-876.
MOUILLOT D,MASON WHN,DUMAY O,et al,2005.Functional regularity:a neglected aspect of functional diversity[J].Oeco-logia,142(3):353-359.
NOGUCHI H,ITOH A,MIZUNO T,et al,2007.Habitat divergence in sympatric Fagaceae tree species of a tropical montane forest in northern Thailand[J].J Trop Ecol,23(05):549-558.
PEI NC,LIAN JY,ERICKSON DL,et al,2011.Exploring tree-habitat associations in a Chinese subtropical forest plot using a molecular phylogeny generated from DNA barcode loci[J]. PLoS One,6:e21273.
PETCHEY OL,GASTON KJ,2006.Functional diversity:back to basics and looking forward[J].Ecol Lett,9(6):741-758.
POORTER L,WRIGHT SJ,PAZ H,et al,2008.Are functional traits good predictors of demographic rates?Evidence from five Neotropical forests[J].Ecology,89(7):1 908-1 920.
PURSCHKE O,SCHMID BC,SYKES MT,et al,2013. Contrasting changes in taxonomic,phylogenetic and functional diversity during a long-term succession:insights into assembly processes[J].J Ecol,101(4):857-866.
RAO CR,1982.Diversity and dissimilarity coefficients:a unified approach[J].Theor Popul Biol,21(1):24-43.
RICOTTA C,2005.A note on functional diversity measures [J].Basic Appl Ecol,6(5):479-486.
ROSCHER C,SCHUMACHER J,LIPOWSKY A,et al,2013.A functional trait-based approach to understand community assembly and diversity-productivity relationships over 7 years in experimental grasslands[J].Persp Plant Ecol,15(3):139-149.
SHEN Y,SANTIAGO LS,MA L,et al,2013.Forest dynamics of a subtropical monsoon forest in Dinghushan,China:recruitment,mortality and the pace of community change[J].J Trop Ecol,29(2):131-145.
SIEFERT A,RAVENSCROFT C,WEISER MD,et al,2013. Functional beta-diversity patterns reveal deterministic community assembly processes in eastern North American trees[J].Glob Ecol Biogeogr,22(6):682-691.
SONG MH,TIAN YQ,XU XL,et al,2006.Interactions between root and shoot competition among four plant species in an alpine meadow on the Tibetan Plateau[J].Acta Oecol,29 (2):214-220.
SPASOJEVIC MJ,SUDING KN,2012.Inferring community assem-bly mechanisms from functional diversity patterns:the importance of multiple assembly processes[J].J Ecol,100(3): 652-661.
SUI YY,JIAO XG,GAO CS,et al,2009.The relationship among or-ganic matter content and soil microbial biomass and soil enzyme activities[J].Chin J Soil Sci,5:1 036-1 039. [隋跃宇,焦晓光,高崇生,等,2009.土壤有机质含量与土壤微生物量及土壤酶活性关系的研究[J].土壤通报,5:1 036-1 039.]
TILMAN D,1982.Resource competition and community structure. (Mpb-17) [M].Princeton University Press.
TSUJINO R,TAKAFUMI H,AGETSUMA N,et al,2006. Variation in tree growth,mortality and recruitment among topo-graphic positions in a warm temperate forest[J].J Veg Sci,17 (3):281-290.
VILLEGER S,MASON NWH,MOUILLOT D,2008.New multidi-mensional functional diversity indices for a multifaceted framework in functional ecology[J].Ecology,89(8):2 290-2 301.
WANG YS,CHUA CJ,MAESTRE FT,et al,2008.On the rele-vance of facilitation in alpine meadow communities:an experi-mental assessment with multiple species differing in their ecolog-ical optimum[J].Acta Oecol,33(1):108-113.
WANG Z,YE WH,CAO HL,et al,2009.Species-topography as-sociation in a species-rich subtropical forest of China[J].Basic Appl Ecol,10(7):648-655.
WANG ZF,LIAN JY,HUANG GM,et al,2012.Genetic groups in the common plant species Castanopsis chinensis and their as-sociations with topographic habitats[J].Oikos,121(12): 2 044-2 051.
YE WH,CAO HL,HUANG ZL,et al,2008.Connunity structure of a 20 hm2lower subtropical evergreen broadleaved forest plot in dinghushan,China[J].Chin J Plant Ecol,32(2):274-286. [叶万辉,曹洪麟,黄忠良,等,2008.鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J].植物生态学报,32(2):274-286.]
Effects of habitat heterogeneity on community functional diversity of Dinghu Mountain evergreen broad-leaved forest
ZHOU Guang-Xia1,2,HUANG Li-Xin1,2,ZANG Xiao-Wei1,2,WEI Xiao3,YE Wan-Hui1,SHEN Hao1∗
(1.Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems,South China Botanical Garden,Chinese Academy of Sciences,Guangzhou 510650,China;2.University of Chinese Academy of Sciences,Beijing 100049,China;3.Guangxi Institute of Botany,Guangxi Zhuang Autonomous Region and Chinese Academy of Sciences,Guilin 541006,China)
Species co-existence mechanisms have long been one of the core topics in community ecology.To explore the changes in community functional diversity of Dinghushan evergreen broad-leaf forest under the heterogeneous habitats and to find out the main driving factors of the functional diversity varations,based on the second community survey data of the 20 hm2forest dynamics plot in Dinghushan National Nature Reserve,27 subplots(20 m×20 m)were selected according to the changes of relative elevation and topograph.These 27 subplots can stand for the heterogeneous habitats in Dinghushan ever-green broad-leaved forest.Leaf functional traits of all the trees with DBH(diameter at breast height)≥1 cm in the 27 sub-plots were measured to calculate the functional diversity(functional dispersion and community weighted mean are caculated as the functional diversity in this paper)during the summer of 2013.The measured traits included morphological traits(spe-cific leaf area,leaf dry matter content,leaf area and leaf lenth to leaf width ratio)and stoichiometric traits(leaf carbon,nitrogen,phosphorus content).Combining with the topographic and soil information,we analysed the main environmental factors that influence the variation of functional diversity.To elucidate the relative importance of environmental filtering and competitive interactions for species co-existence of Dinghushan evergreen broad-leaved forest,we inferred the shifts in func-tional diversity patterns across environmental gradients.The results indicated that functional divergence and community-weighted mean were in close relation with environmental factors,especially elevation,convexity and soil fertility.In the op-timal environmental conditions(high relative elevation,high convexity and higher soil nutrient content),plants adopt the similar(functional divergence decreased)adaptive strategy of efficient conservation of nutrient(low SLA,high LDMC),which suggested that environmental filtering played a stronger role;while in the adverse environmental conditions(low rela-tive elevation,low convexity and lower soil nutrient content),the traits were various(functional divergence increased)and use fast growing strategy(high SLA,low LDMC),since competitive interactions might play a key role.The functional di-vergence of leaf area and leaf nitrogen contents increased in better environmental conditions(high relative elevation,high convexity and higher soil nutrient content),and these two traits(leaf area and leaf nitrogen content)were opposite to the others(specfic leaf area,leaf dry matter content,leaf length to leaf width ratio,leaf carbon and phosphorus content).These results indicated that different niche axes(environments)had different importances in the changes in func-tional traits dispersion,and illustrated that we could use plants diverse strategies to adapt to heterogeneous environments in local scale.
community ecology,functional diversity,habitat heterogeneity,species co-existence mechanisms,adap-tive strategy
Q948.15,Q945.79
A
1000-3142(2016)02-0127-10
10.11931/guihaia.gxzw201503015
周光霞,黄立新,臧晓蔚,等.生境异质性对鼎湖山常绿阔叶林群落功能多样性的影响[J].广西植物,2016,36(2):127-136
ZHOU GX,HUANG LX,ZANG XW,et al.Effects of habitat heterogeneity on community functional diversity of Dinghu Mountain evergreen broad-leaved forest[J].Guihaia,2016,36(2):127-136
2015-04-10
2015-06-24
国家自然科学基金(31370446)[Supported by the National Natural Science Foundation of China(31370446)]。
周光霞(1990-),女,安徽芜湖人,硕士,主要从事植物生理生态学研究,(E-mail)zhouscbg@163.com。
∗通讯作者:沈浩,博士,副研究员,主要从事保护生态学和植物生理生态学研究,(E-mail)shenhao@scbg.ac.cn。
