极小种群野生植物馨香木兰的种群结构及动态
2016-05-07徐加汉司马永康徐涛猫玉白余鸿
徐加汉,司马永康,徐涛,猫玉白,余鸿
(1.云南省林业科学院 云南省森林植物培育与开发利用重点实验室/国家林业局重点开放性实验室
云南珍稀濒特森林植物保护和繁育实验室,云南 昆明650201;2.云南省文山州林木种苗工作站,云南 文山663000;
3.云南大学生命科学学院,云南 昆明650091;4.昆明生物资源与多样性研究中心,云南 昆明650504;
5.云南省环境工程评估中心,云南 昆明650034)
极小种群野生植物馨香木兰的种群结构及动态
徐加汉1,2,司马永康1,徐涛3,4,猫玉白5,余鸿4
(1.云南省林业科学院 云南省森林植物培育与开发利用重点实验室/国家林业局重点开放性实验室
云南珍稀濒特森林植物保护和繁育实验室,云南昆明650201;2.云南省文山州林木种苗工作站,云南文山663000;
3.云南大学生命科学学院,云南昆明650091;4.昆明生物资源与多样性研究中心,云南昆明650504;
5.云南省环境工程评估中心,云南昆明650034)
摘要:为了更好地掌握极小种群野生植物馨香木兰的种群结构及动态情况,通过对云南省西畴县馨香木兰群落24个样方的调查,分析了馨香木兰的高度结构、年龄结构、空间分布格局,编制了特定时间生命表,绘制了生存曲线。结果表明,馨香木兰高度结构完整,种群个体高度发育是连续的,在群落中属连续型种群;其相对年龄结构不完整,呈现明显的菱形,幼苗缺乏;其空间分布格局表现为集群分布,但到IV级立木阶段就已变成随机分布;其成长可分为幼苗、幼树、成树和老树等4个时期,幼树和成树时期死亡率最低,老树时期死亡率最高;其种群生存曲线接近于Deevey I型,种群正处于衰退状态。
关键词:极小种群野生植物;馨香木兰;种群结构动态;生命表;生存曲线
野生植物资源是人类赖以生存的物质基础。随着近几十年来人类社会经济的迅猛发展,资源的过度开发利用、生境的破坏等已致使许多野生物种严重濒危,以至灭绝[1~3]。保护濒危物种已成为一个紧迫的保护生物学课题[4]。其中,研究并掌握某个濒危植物物种的种群结构及动态,不仅有助于了解该种濒危植物种群的生存现状,为揭示濒危物种种群生活史中的薄弱环节、分析致濒的原因提供基础资料,而且还有助于该种群合理的保护和增殖方案的制定,为进一步的深入研究提供科学依据[5~6]。
馨香木兰(Liriantheodoratissma(Y.W.Law et R.Z.Zhou) N.H.Xia et C.Y.Wu)又称馨香玉兰、董马木兰、调羹花或夜来香,属木兰科(Magnoliaceae)喙木兰属常绿小乔木,仅分布于中国云南省东南部和越南北部[7~12],通常生长于海拔1 100~1 600m的常绿阔叶林中[7~8,13~14]。馨香木兰分布区域狭窄,加之可供观赏和能提取高品质香精[15~18],野生资源开发利用过度,个体数量极少[8,19],属极小种群野生物种[8,20],已被列为国家和云南省II级重点保护野生植物[21~22]。目前,有关馨香木兰的分类学与地理分布[7~12,23~26]﹑形态解剖学[27]、细胞学[28~29]、植物化学[15~16,30]﹑分子系统学[31~32]、群落生态学[13~14,33~34]、种群生态学[13,35]、遗传育种学[36~38]、土壤学[39]、引种驯化与保护[17~18,20,40~50]等研究已有很多报道,但有关馨香木兰种群结构及动态的研究鲜见报道。本文分析探讨了云南省西畴县新马街乡现存馨香木兰种群的结构及动态,以期为馨香木兰的濒危机制分析及其今后的保护和利用等提供可靠的基础资料[51]。
1研究地自然概况
研究地处在云南省西畴县新马街乡小坪坝村,23°12′13″~44″N,104°31′11″~30″E,隶属湿润温暖的暖亚热带季风气候。年平均气温15.3~16.1℃,≥10℃年活动积温4 830.7℃,最热月平均气温21.4℃,极端最高气温33.2℃,最冷月平均气温7.6℃,极端最低气温-7.2℃;全年霜期9.3d;年降水量为1 266~1 290mm,年平均相对湿度83%;日照时数1 473h,日照百分率为34[52]。在此,馨香木兰主要生长于石灰岩山地季风常绿阔叶林中,海拔在1 410~1 570m之间,坡度为20°~53°,坡型为直线坡和凹形坡,土壤为石灰岩发育而成的黄红壤。群落高7~13m不等,乔木层特别发达,层盖度在80%~98%之间;灌木层不发达到发达,层盖度为20%~70%;草本层不发达到发达,层盖度为20%~80%。伴生树种主要有清香木(PistaciaweinmanniifoliaJ.Poisson ex Franch.)、滇南青冈(CyclobalanopsisaustroglaucaY.T.Chang ex Y.C.Hsu & H.W.Jen)、云桂鹅耳枥(CarpinuspubescensBurkill)、红梗润楠(MachilusrufipesH.W.Li)、香叶树(LinderacommunisHemsl.)、紫弹树(CeltisbiondiiPamp.)、密花树(MyrsineseguiniiH.Lévl.)、圆果化香树(PlatycaryalongipesY.C.Wu)、白蜡树(FraxinuschinensisRoxb.)、黄连木(PistaciachinensisBunge)、假桂皮树(Cinnamomumtonkinense(Lecomte) A.Chev.)等。人为干扰主要是频繁的砍伐攀折,偶尔有践踏和放牧。
2研究方法
2.1样方设置和调查
采用典型抽样的方法,在有馨香木兰出现的群落内,设置10m×10m的样方24个,调查总面积达2 400m2,对各个样方的坡位、坡度、坡向、海拔和土壤类型等生境因子,群落高、层盖度以及人为干扰方式和强度进行调查和记录。同时,对样方内物种逐株测量记录高度和胸径等。
2.2数据统计和分析方法
2.2.1高度结构
根据上限排外法,以2.6m为一个高度级对各高度级内的株数进行统计,确定高度结构,制作种群高度级结构图[5~6,51]。
2.2.2年龄结构
根据1930年H.J.Lutz于提出的方法,以立木大小级作为相对年龄来代替不同的年龄,并将立木划分为4级:I级为高度<33cm;II级为高度≥33cm,胸径<2.5cm;III级为2.5cm≤胸径<7.5cm;IV级为7.5cm≤胸径<22.5cm。分级统计株数,制作种群相对年龄结构图[5~6,51]。
2.2.3空间分布格局
采用方差均值比率法判定种群水平空间分布格局,即在泊松分布中的方差(S2)与均值(M)之比等于或接近于1,为随机分布;如果S2/M的值小于1,则为均匀分布;S2/M的值大于1,则为集群分布。同时,采用t检验来确定实测值与预期值的偏离程度[5~6,51]。
2.2.4生命表
根据特定时间生命表的编制原理和方法[53],利用立木相对年龄大小级来代替绝对年龄级,计算各参数,制作出馨香木兰种群特定时间生命表。具体计算参数和方法参考郝佳波等[5]的方法。
2.2.5生存曲线
在种群生存曲线的制作中,以立木相对年龄大小级为横轴,以ln(lx)为纵轴,以种群特定时间生命表数据为依据。
3结果与分析
3.1高度结构
根据高度级结构(图1),馨香木兰种群的高度级可分为5个,其比例各为38.78%、29.59%、20.41%、9.69%和1.53%。馨香木兰株数随高度级的增加而递减,减速在8.16%~10.72%之间,递减速度相对均匀。其中,除由5.2~7.8m向7.8~10.4m间的递减最快外,其余各级间的递减变化则较为缓慢。由此说明,馨香木兰种群具有完整的高度结构,种群个体高度发育连续,为连续型种群。馨香木兰种群高度结构呈金字塔形,顶部较狭窄,基部最宽,储备相对幼树个体较丰富,对在群落中下一层个体向上一层补充有利。
3.2年龄结构
根据大小级结构(图2),馨香木兰种群的立木大小级可分为4级,其比例各为0.00%、46.94%、46.94%和6.12%。馨香木兰种群株数在第I级向第II级发展时急剧递增,在第III级向第IV级发展时迅速递减,而在第II级向第III级发展时则无增减。由此说明,馨香木兰种群具有不完整的年龄结构,I级幼苗缺乏,大小级结构呈明显的菱形,两头狭窄,中间宽,储备幼树和成树个体特别丰富,老树个体占少数,幼苗个体缺乏。由此可知,虽然馨香木兰幼树和成树的个体较多,但是种子萌发受阻,自然更新能力下降。如果此后更新状况仍得不到改善,没有幼苗向幼树层补充,则本研究区内的馨香木兰将发展成为衰退种群。
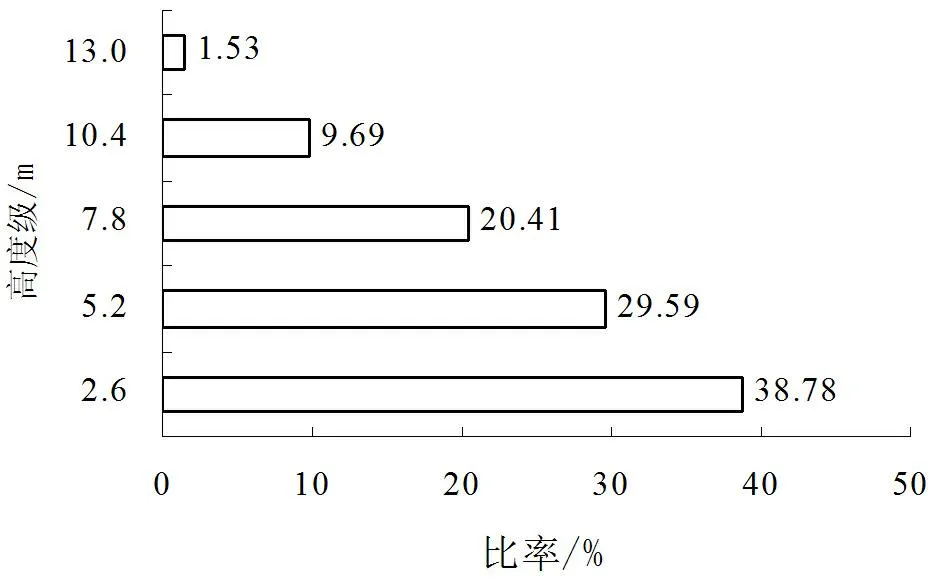
图1 馨香木兰种群高度级结构
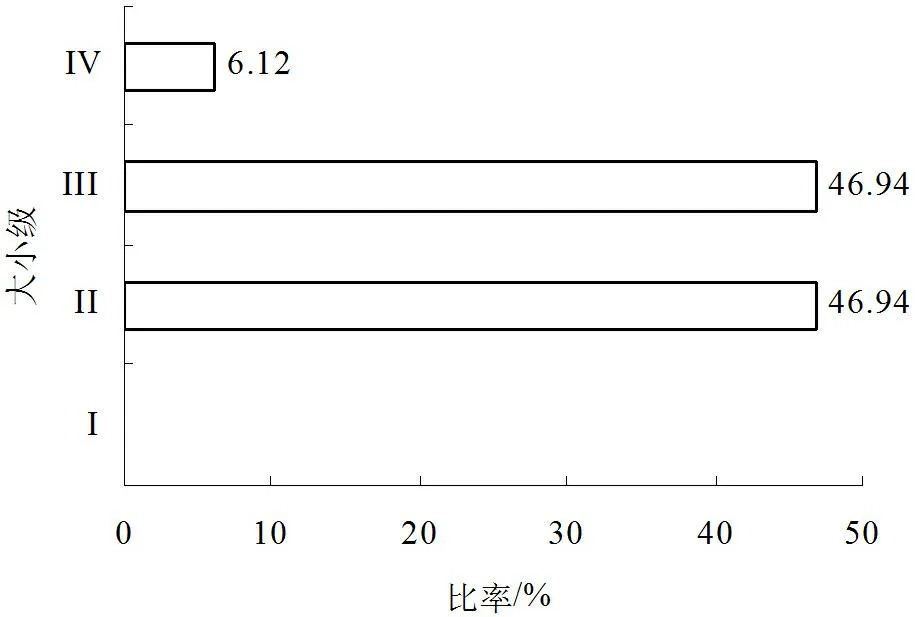
图2 馨香木兰种群大小级结构
3.3空间分布格局
根据空间分布格局情况(表1),馨香木兰种群总体为集群分布。而根据相对年龄划分,馨香木兰种群除II级至III级均为集群分布外,到IV级时就变为随机分布。显然,馨香木兰种群个体的集群强度随着种群的不断发育和年龄增加而快速减小,最后解体转化为随机分布。
3.4生命表
根据馨香木兰种群特定时间生命表数据(表2),馨香木兰种群由幼苗生长到死亡(qx)可大体分为4个阶段。
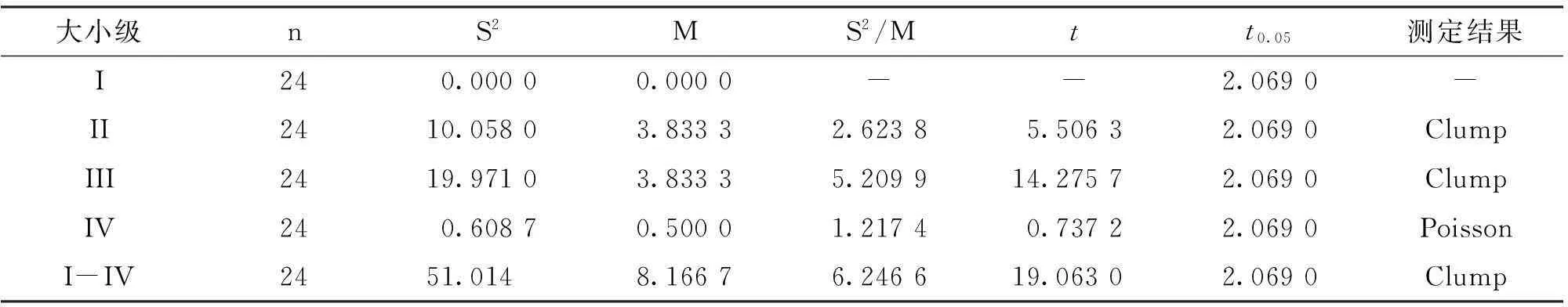
表1 馨香木兰种群空间分布格局
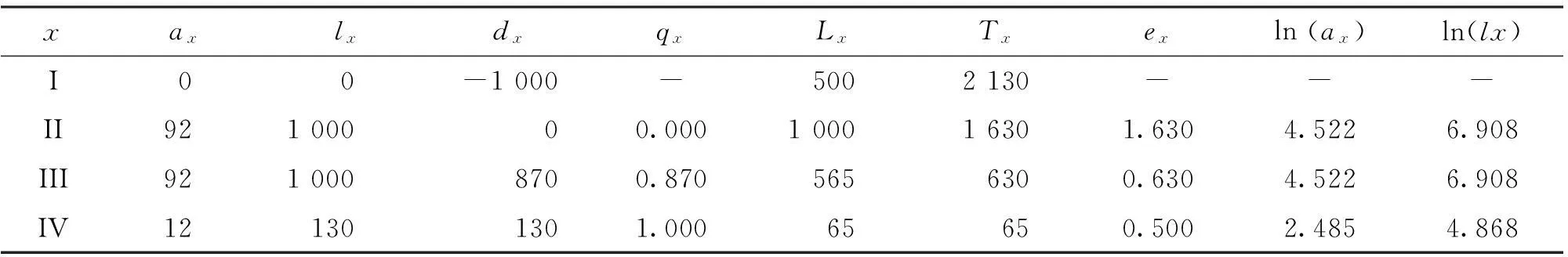
表2 馨香木兰种群特定时间生命表
(1)幼苗阶段(I~II级立木前期阶段)该阶段因无I级幼苗,无法统计死亡率。
(2)幼树阶段(II~III级立木前期阶段)该阶段的死亡率最低,为0。该阶段处在幼树向青年期转变的早期,馨香木兰多已处于灌木层向乔木层林木靠近的空间,由于灌木层不发达至发达,相对喜欢荫湿的馨香木兰处于较强的竞争优势,个体间的分化小,导致较低的死亡率。
(3)成树阶段(III~IV级立木前期阶段)该阶段的死亡率为0.870,立木已处在成年期,作为乔木层亚优势植物,馨香木兰进一步向乔木上层发展,与其他林木的竞争进一步加强,在乔木层种内种间竞争的抑制下,自疏和他疏作用激烈增强,分化在个体间非常严重,因而死亡率极高。
(4)老树阶段(IV级立木阶段)该阶段的死亡率为1.000,达到最高水平,馨香木兰立木已处在老年期,已明显出现生理功能的衰退,馨香木兰生命历程的最终死亡高峰形成。总之,III级立木阶段大约是馨香木兰种群的生理年龄。
x龄级内个体的平均生存能力是由平均生命期望(ex)反映的。馨香木兰种群在II~III级立木前期阶段时ex值达到最大。作为乔木层亚优势种的馨香木兰个体处于竞争优势,竞争压力小,具有最高的生存质量和旺盛生理活动;在III~IV级立木阶段时,馨香木兰种群的平均生命期望急减,竟不足II~III级立木阶段的1/2,馨香木兰种群生理衰退明显。
3.5生存曲线
制作馨香木兰种群的生存曲线(图3)时,采用相对年龄大小级为横轴和ln(lx)为纵轴。参照Deevey生存曲线[54]与馨香木兰种群的生存曲线比较发现,馨香木兰种群的生存曲线介于Deevey I和II型曲线之间,靠近Deevey I型,呈凸型明显。说明馨香木兰种群在接近生理寿命之前只有少数个体死亡。
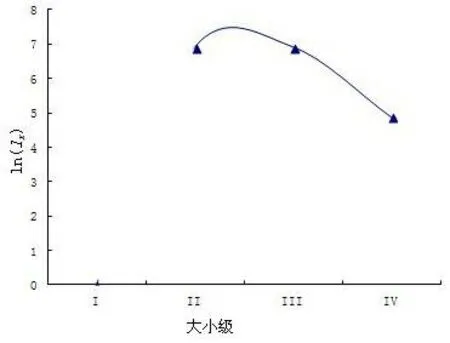
图3 馨香木兰种群的生存曲线
4结论与讨论
种群结构是种群的基本特征,包括种群的数量、年龄、性比、密度、高度等结构要素[6,51,53,55],其高度结构和年龄结构在很大程度上不仅预示了种群的数量变化和未来的动态发展趋势,而且反映了种群当时的发育阶段和状况[6]。本研究结果表明,馨香木兰种群的高度结构完整,呈现明显的金字塔形,基部最宽,顶部较狭窄,储备相对幼树个体较丰富,对在群落中下一层个体向上一层补充有利,而各高度级阶梯中均有馨香木兰的个体分布,表明种群个体高度发育是连续的,在群落中属连续型种群。这与馨香木兰在群落中为亚优势种群的事实一致[13~14]。馨香木兰作为群落的亚优势种,不受群落乔木上层其他树种因竞争关系而对其高度的抑制,在践踏和放牧等人为干扰少而轻的情况下,才在高度结构中形成如此自幼树向大树个体数量逐级递减的现象。
在本研究区域内,馨香木兰种群具有不完整的年龄结构,I级幼苗缺乏,大小级结构呈明显的菱形,两头狭窄,中间宽,储备幼树和成树个体特别丰富,老树个体占少数,幼苗个体缺乏。由此而论,馨香木兰种群很可能遭遇到了人为干扰,致使种子不能萌发,很难完成自然更新。如果此后几年自然更新仍不能顺利完成的话,本地馨香木兰种群将发展成为衰退种群。众所周知,频繁而严重的放牧和践踏对种子和幼苗的破坏是十分严重的,对幼苗的形成和生长极其不利。而就其调查样地内人为干扰以砍伐攀折为主兼有少量的放牧和践踏等而言,显然,现存馨香木兰种群生长于陡峭的石灰岩小山包上,不存在放牧和践踏的严重影响。而就现有的馨香木兰生存环境看,已多为无法开垦或开发为农用地或居民地的陡峭的石灰岩小山包上,在影像图上看似农用地和居民地中深色的“孤岛”。“孤岛”生境的湿润程度明显比未开垦或开发前干燥很多。根据观察,馨香木兰的种子在坚硬的种皮外具有一层鲜艳红色或桔红色的假种皮,对种子的萌发具有一定的化学抑制作用,通常只有在较湿润和湿润的生境中,通过淋溶作用和腐化作用才能解除这种化学抑制和坚硬种皮的物理抑制而萌发[51]。当然,也可通过鲜艳的假种皮色彩吸引鸟类取食,经鸟类消化腔解除这种化学抑制和物理抑制,并随鸟类粪便下种萌发[51]。众所周知,鸟类对馨香木兰种子的取食,其主要作用在于对种子的传播和扩散。就当地而言,较湿润和湿润的生境,才是决定种子是否萌发为幼苗的必备条件。因此,“孤岛”生境的干燥化可能是造成馨香木兰种群缺乏幼苗的直接原因。馨香木兰种群的“孤岛”生境明显是由近一二十年来人为干扰造成的,归根结底,过度的人为干扰才极有可能是形成馨香木兰种群年龄结构缺乏幼苗而为衰退种群的真正或间接原因。
种群空间分布格局通常反映着一定环境因子对个体行为、生存和生长的影响,是种群对环境长期适应和选择的结果[53]。馨香木兰的种群空间分布格局为集群分布,与馨香木兰种子的传播途径和生物学特性等有关。馨香木兰种子除偶尔被鸟类取食外,都会落下,通常在母树周围集中散布,并在湿润、土壤肥厚的条件下萌发,以集群分布格局局限分布在水肥条件相对丰富的直线坡和凹形坡中,偶然成为群落的亚优势种群之一。但随着种群的不断发育,种内种间的竞争加剧,导致种群个体的集群强度随着年龄的增加而快速减小,最后集群解体转化为随机分布[6]。馨香木兰种群在II级和III级立木阶段时均表现为集群分布,但到IV级立木阶段时就转化为随机分布。种群在幼年阶段集群强度高有利于生存和发挥群体效应,到成年时个体由于变大,拥挤效应和自疏作用增强[56],减小集群强度有利于获得足够的环境资源[6]。总之,种群集群强度随年龄的增减而变化是种群的一种适应机制或生存策略[6]。
致谢:衷心感谢西畴县林木种苗工作站杨耀海高工和西畴县新马街乡小坪坝村用天国先生在野外调查上的尽心帮助。
参考文献:
[1]Diamond J.Overview of recent extinctions [A].Western D,Pearl MC.Conservation for the twenty-first century [M].Oxford:Oxford University Press,1989.
[2]钱迎倩,马克平.生物多样性研究的原理与方法[M].北京:中国科学技术出版社,1994.
[3]蒋志刚,马克平,韩兴国.保护生物学[M].杭州:浙江科学技术出版社,1997.
[4]Lindborg R,Ehrlen J.Evaluating the extinction risk of a perennial herb:demographic data versus historical records [J].Conservation biology,2002,16:683-690.
[5]郝佳波,司马永康,陈少瑜,等.麻栗坡大果木莲的居群结构及动态研究[J].西部林业科学,2012,41(3):84-88.
[6]付玉嫔,司马永康,方波,等.篦子三尖杉的居群结构与动态研究[J].广东农业科学,2015,42(11):48-54.
[7]司马永康,余鸿.木兰属一新命名[J].云南林业科技,2003(1):39.
[8]司马永康.馨香木兰[A].见:李玉媛,郭立群,胡志浩.云南国家重点保护野生植物[M].昆明:云南科学技术出版社,2005:211-212.
[9]Xia N-H,Liu Y-H(Law Y-W),Nooteboom HP.Magnoliaceae [A].In:Wu Z-Y,Raven PH,Hong D-Y.Flora of China,7[M].Beijing:Science Press; St.Louis:Missouri Botanical Garden Press,2008:48-91.
[10]Sima Y-K,Lu S-G.2009.Magnoliaceae [A].In:Shui Y-M,Sima Y-K,Wen J,etal. Vouchered flora of Southeast Yunnan,1 [M].Kunming:Yunnan Publishing Group Corporation; Yunnan Science & Technology Press,2009:16-67.
[11]司马永康.中国木兰科植物的分类学修订[D].昆明:云南大学,2011.
[12]Vu QN,Xia N-H.Taxonomy and biogeography of the family Magnoliaceae from Vietnam [A].In: Xia N-H,Zeng Q-W,Xu F-X,etal. Proceedings of the Second International Symposium on the Family Magnoliaceae [M].Wuhan:Huazhong University Science & Technology Press,2012:95-115.
[13]柴勇,孟广涛,武力,等.馨香木兰所处植物群落的物种多样性及其种群结构特征研究[J].西部林业科学,2008,37(3):46-50.
[14]董莉,柳小强,余德恒,等.云南省极危种馨香木兰的群落特征初步研究[J].安徽农业科学,2010,38(22):12068-12069,12297.
[15]芮和恺,季伟良,李达孝,等.馨香木兰叶的精油成份分析[J].云南林业科技,1992(2):64-66.
[16]李玉媛,李达孝,毛云玲,等.云南木兰科香料植物浸膏、精油提取及化学成分研究[J].香料香精化妆品,1996(3):2-8.
[17]李玉媛,李达孝.云南木兰科植物的保护价值与开发前景[J].北京林业大学学报,1999,21(3):29-35.
[18]杨耀海.馨香木兰扦插繁殖试验[J].林业科技开发,2009,23(2):103-105.
[19]顾云春.中国国家重点保护野生植物现状[J].中南林业调查规划,2003,22(4):1-7.
[20]郑进烜,华朝朗,陶晶,等.云南省极小种群野生植物拯救保护现状与对策研究[J].林业调查规划,2013,38(4):61-66.
[21]于永福.中国野生植物保护工作的里程碑——国家重点保护野生植物名录(第一批)出台[J].植物杂志,1999(5):5-11.
[22]周彬.云南省第一批省级重点保护野生植物名录修订[J].云南植物研究,2010,32(3):221-226.
[23]刘玉壶,周仁章.广东,云南木兰科一新种[J].植物研究,1986,6(2):139-142.
[24]刘玉壶,吴容芬.木兰科[A].见:中国科学院中国植物志编辑委员会.中国植物志,[M].北京:科学出版社,1996:82-198.
[25]刘玉壶,曾庆文,周仁章,等.中国木兰[M].北京:北京科学技术出版社,2004.
[26]夏念和.国产木兰科的分类修订[D].昆明:中国科学院昆明植物研究所,2007.
[27]吴涛,韩明跃,司马永康,等.木兰科6属27种植物的叶片比较解剖学研究[J].西部林业科学,2013,42(6):25-37.
[28]王亚玲,张寿洲,李勇,等.木兰科13个分类群和12个杂交组合的染色体数目[J].植物分类学报,2005,43(6):545-551.
[29]赵强民,何子灿,王亚玲.木兰科5个种间杂种减数分裂行为观察[J].亚热带植物科学,2014,43(2):119-122.
[30]石文华,银建中,徐巧莲.鲜花精油和浸膏的超临界CO2萃取进展[J].精细化工,2004,21(增刊):103-107.
[31]王亚玲,李勇,张寿洲,等.用matK序列分析探讨木兰属植物的系统发育关系[J].植物分类学报,2006,44(2):135-147.
[32]Nie Z-L,Wen J,Azuma H,etal. Phylogenetic and biogeographic complexity of Magnoliaceae in the Northern Hemisphere inferred from three nuclear data sets [J].Molecular Phylogenetics and Evolution,2008,48:1027-1040.
[33]柴勇,孟广涛,武力,等.馨香玉兰所在群落主要树种的种间联结性[J].西北林学院学报,2009,24(5):31-35.
[34]邵金平,李贵祥,柴勇,等.馨香玉兰及其伴生树种的生态位研究[J].西部林业科学,2015,44(1):64-68.
[35]柴勇,祁荣频,孟广涛,等.云南特有种馨香木兰种群格局的分形特征[J].东北林业大学学报,2013,41(5):24-27.
[36]王亚玲.玉兰亚属的研究[D].杨凌:西北农林科技大学,2003.
[37]王亚玲,李勇,张寿洲,等.木兰科植物的人工杂交[J].武汉植物学研究,2003,21(6):508-514.
[38]金蕊,石雨鑫,徐涛,等.云南特有濒危植物馨香木兰的遗传多样性研究[J].西部林业科学,2014,43(6):80-84.
[39]祁荣频,柴勇,武力,等.馨香木兰集中分布地生境的土壤理化性状研究[J].西部林业科学,2010,39(4):95-98.
[40]李达孝,杨绍诚,税希特.云南木兰科植物物种资源及其种质库的研究[J].生物多样性,1995,3(4):195-200.
[41]张茂钦.云南珍稀濒危树种生态生物学研究[M].昆明:云南大学出版社,1998.
[42]周际中,李庆华,李革,等.昆明基地珍稀濒危植物引种驯化[J].云南环境科学,2003,22(增刊1):1-9.
[][]
[43]曹基武,刘春林,祁承经,等.18个木兰科树种播种育苗技术初步研究[J].林业科技,2008,33(5):16-19.
[44]韩春艳.木兰科几种植物种子休眠和种质资源保存的研究[D].昆明:中国科学院昆明植物研究所,2008.
[45]杨祖顺.馨香木兰育苗技术[J].林业科技开发,2008,22(4):109-111.
[46]杜有新,张青松,庞宏东,等.东亚-北美间断分布植物的早期适应性[J].生态环境学报,2009,18(3):990-995.
[47]Cao J-W,Liu C-L,Wu Y,etal. Studies on seed germination and seedling growth mechanism of six endangered Magnoliaceous plants[A].In: Xia N-H,Zeng Q-W,Xu F-X,etal. Proceedings of the Second International Symposium on the Family Magnoliaceae[M].Wuhan:Huazhong University Science & Technology Press,2012:247-255.
[48]沐建华.文山州云南省重点保护野生植物资源及其价值[J].文山学院学报,2012,25(3):19-23.
[49]Yang Y-H.Propagation of rare and endangered plantMagnoliaodoratissima[A].In: Xia N-H,Zeng Q-W,Xu F-X,etal. Proceedings of the Second International Symposium on the Family Magnoliaceae[M].Wuhan:Huazhong University Science & Technology Press,2012:236-246.
[50]孙卫邦,韩春艳.论极小种群野生植物的研究及科学保护[J].生物多样性,2015,23(3):426-429.
[51]司马永康,刘伦辉,蒋宏.珍稀濒危植物监测与结果分析[A].见:李玉媛.菜阳河自然保护区定位监测[M].昆明:云南大学出版社,2003:93-126.
[52]云南省气象局.云南气候图册[M].昆明:云南人民出版社,1982.
[53]王伯荪,李鸣光,彭少麟.植物种群学[M].广州:广东高等教育出版社,1995.
[54]Leak WB.Age distribution in virgin red spruce and northen hardwoods[J].Ecology,1975,56:1451-1454.
[55]郑诗章,吴千红,王海波,等.普通生态学——原理、方法和应用[M].上海:复旦大学出版社,1994.
[56]Molles MC.Ecology:Concepts and applications[M].Beijing:China Science Press & McGraw-Hill Companies,Inc.,2000:176-177.
Population Structure and Dynamics of Lirianthe odoratissma(Y.W.Law et R.Z.Zhou) N.H.Xia et C.Y.Wu,a Plant Species with Extremely Small Populations
XU Jia-han1,2,SIMA Yong-kang1,XU Tao3,4,MAO Yu-bai5,YU Hong4
(1.Yunnan Provincial Key Laboratory of Cultivation and Exploitation of Forest Plants/Yunnan Laboratory for Conservation of Rare,Endangered &Endemic Forest Plants,Public Key Laboratory of the State Forestry Administration,Yunnan Academy of Forestry,Kunming Yunnan 650201,P.R.China;2.Forest Seed and Seedling Station of Wenshan Prefecture,Wenshan Yunnan 663000,P.R.China;3.School of Life Science,Yunnan University,Kunming Yunnan 650091,P.R.China;4.Kunming Center of Biological Resources & Biodiversity Researches,Kunming Yunnan 650504,P.R.China;5.Yunnan Provincial Appraisal Center for Environmental Engineering,Kunming Yunnan 650034,P.R.China)
Abstract:In order to better understand the population structure and dynamics of Lirianthe odoratissma(Y.W.Law et R.Z.Zhou) N.H.Xia et C.Y.Wu,a plant species with extremely small populations,the height and age structure and the spatial distribution of its population were analyzed,the time-specific life table was established and the survivorship curve was drawn with the data of the survey of 24 sample plots of its communities in the Xichou county of Yunnan province.The results indicated that the height structure was complete.This implies that the population is successive in the height growth of its individuals and in community.The relative age structure is uncomplete and its histogram looks like a diamond while the numbers of seedlings are lacking and old trees are too less.Its spatial distribution pattern is of clump and that of the IV-class population become of poisson.Its development can be divided into 4 phases,namely seedling,young,adult and old phases.Its mortality rate is the lowest in the young and adult phases and the highest in the old phase.The survivorship curve is close to the type of Deevey I and this reveals that the population is declining.
Key words:plant species with extremely small populations;Lirianthe odoratissma(Y.W.Law et R.Z.Zhou) N.H.Xia et C.Y.Wu; population structure dynamics;life table;survivorship curve
中图分类号:S 792.99
文献标识码:A
文章编号:1672-8246(2016)02-0035-07
通讯作者简介:司马永康(1967-),男,研究员,博士,主要从事木兰科植物分类学、保护生物学和资源植物学研究。E-mail:simayk@163.com。
第一作者简介:徐加汉(1967-),男,工程师,主要从事木兰科植物繁育栽培技术和保护研究。E-mail:1784124664@qq.com
基金项目:国家自然科学基金项目(30660154,31060096),云南省自然科学基金项目(2011FZ008),云南省中青年学术和技术带头人后备人才培养项目(2012HB025)。
*收稿日期:2015-12-19

