马尾松GGPPS基因克隆及序列分析*
2016-05-07陈博雯覃子海王鹏良贾婕陈虎刘海龙
陈博雯,覃子海,王鹏良,贾婕,陈虎,刘海龙
(广西林业科学研究院,广西优良用材林资源培育重点实验室,广西 南宁530002)
马尾松GGPPS基因克隆及序列分析*
陈博雯,覃子海,王鹏良,贾婕,陈虎,刘海龙
(广西林业科学研究院,广西优良用材林资源培育重点实验室,广西南宁530002)
摘要:牻牛儿基牻牛儿基焦磷酸合成酶(GGPPS)是萜烯类物质合成的关键酶之一,本研究通过同源序列检索分析设计引物,从马尾松幼苗针叶组织中克隆到其GGPPS基因,命名为PmaGGPPS,该基因cDNA长1 143bp,开放阅读区编码380个氨基酸。PmaGGPPS编码的蛋白质序列分析显示具有Trans IPPS结构域及FARM、SARM两个富含天冬氨酸区域,序列特点与GGPPS蛋白的酶学功能相符。同源性分析结果显示PmaGGPPS核酸序列与其他两种松属植物GGPPS基因同源性达到99%以上,PmaGGPPS编码蛋白与挪威云杉等植物中GGPPS蛋白同源性也在69%以上。对PmaGGPPS编码蛋白进行理化性质及结构的生物信息学分析,结果显示该蛋白为一种无信号肽、无跨膜区的碱性蛋白质,二级结构由16段α-螺旋组成,三级结构同源建模显示与欧薄荷的异聚牻牛儿基焦磷酸合酶大亚基同源性最高。系统进化树分析显示该蛋白与红豆杉、银杏亲缘关系较近。本研究中克隆得到PmaGGPPS基因为深入研究其在松脂合成的影响奠定了基础,同时根据GGPPS基因设计荧光定量引物并优化反应条件,为下一步研究基因表达水平与产脂力关系提供了理论依据和技术参考。
关键词:马尾松;牻牛儿基牻牛儿基焦磷酸合成酶(GGPPS);基因克隆;荧光定量PCR
针叶树可分泌大量有特殊气味的树脂,也称之为松脂,是松树生理代谢的次生产物,也是重要的工业原料,长期以来为人们所研究和开发利用[1]。全世界约有20种产脂力较高的松树被用作主要采脂树种,马尾松(Pinusmassoniana)是中国最主要的采脂树种,有90%以上的松脂采自马尾松[2~3]。
松脂可加工分离为松节油和松香,其中松节油的主要成分是单萜和倍半萜化合物,约占松脂总量的1/5;松香的主要成分是树脂酸,是一类二萜化合物的异构体混合物,约占松脂总量的3/4[4~5]。从松脂组成成分分析,萜类物质的合成对松脂产量影响极大,特别是二萜化合物的合成,对松脂的产量和质量至关重要。从萜类合成途径中的酶入手,对其编码基因展开研究,并进一步利用DNA重组技术和转基因手段,定向改造萜类合成代谢途径,促进有用萜类合成,是松脂合成定向调控领域较为主流的研究思路[6]。
萜烯类的生物合成途径有两条,一条为甲羟戊酸途径,另一条为甲基-D-赤藓醇-4-磷酸途径。两条途径都产生萜类合成的共同的前体异戊烯焦磷酸(IPP)或其异构体二甲丙烯焦磷酸(DMAPP)。IPP和DMAPP在相应酶的作用下,合成牻牛儿基焦磷酸(GPP)、橙花基焦磷酸(NPP)、法呢基焦磷酸(FPP)和牻牛儿基牻牛儿基焦磷酸(GGPP)。以上前体再经各种萜类合成酶催化单萜、倍半萜、二萜[6~7]。
牻牛儿基牻牛儿基焦磷酸合成酶(GGPPS)是萜烯类物质合成的一个重要分支酶,催化GPP与2个IPP缩合成GGPP,GGPP是二萜合成中的关键前体,推测GGPPS基因很可能是松脂形成中的影响因子之一。
目前在针叶树中已报道了70多种萜类合成酶,主要是在大冷杉(Abiesgrandis)和挪威云杉(Piceaabies)中的研究报道,火炬松(Pinustaeda)、花旗松(Pseudotsugamenziesii)中也有少量研究,马尾松中则未见相关报道[7]。
本研究拟利用RT-PCR方法对马尾松中克隆得到的二萜底物合成酶GGPPS进行序列鉴定分析,同时根据目标基因设计荧光定量引物并优化反应条件,为下一步研究基因表达水平与产脂力关系提供理论依据和技术参考。
1材料方法
1.1实验材料
供试材料为马尾松古蓬种源0.5年生实生苗,取自广西林业科学研究院用材林研究所苗圃。
植物总RNA提取试剂盒、DNA提取、纯化及回收试剂盒购自北京天根生化科技有限公司。反转录试剂盒、LA Taq DNA聚合酶、载体pMD-18T购自TaKaRa公司,定量PCR试剂盒ABI SybrGreen PCR Master Mix(2X)购自上海生工;限制性内切酶购自Promega公司,抗生素及其他生化试剂购自北京索莱宝科技有限公司。E.coliDH5α菌株为广西优良用材林资源培育重点实验室保存。引物合成及测序由深圳华大基因生物技术有限公司完成。
1.2GGPPS基因的克隆
取0.5年生实生苗针叶组织,按照试剂盒说明书方法提取总RNA并反转录合成cDNA,保存备用。
根据NCBI检索已报道的GGPPS基因序列,用Vector NTI软件分析保守区并设计引物:Sense primer GGPPS-F1:ATGGCTTACAGTGGTAGAC,Anti-sense primer GGPPS-R1:TCAGTTTTGTCGAATGCAA。
以cDNA作为模板,扩增得到的目的基因cDNA片段与pMD-18T载体连接并转化至E.coliDH5α,随机挑取阳性克隆PCR验证后测序。
1.3序列生物信息学分析
采用多种在线分析工具以及生物信息学分析软件[8]对克隆到的基因序列进行分析:首先对序列用NCBI的Blastn在线工具进行同源性分析;利用ORF Finder搜寻阅读框并对DNA序列翻译;用Blastp分析蛋白序列同源性及保守区域、活性位点;运用ExPaSy网站的ProtParam完成蛋白序列的氨基酸组成、等电点及亲疏水性分析;应用丹麦科技大学(DTU)提供的TMHMM和SignalP工具分析序列中的跨膜区和信号肽;利用ExPaSy中的psipred分析序列二级结构;应用SWISS-MODEL软件对基因氨基酸序列进行同源三级结构建模并分析;应用MEGA 5.0工具的ClustalW分析PmaGGPPS与其他物种中GGPPS基因序列,并采用Neighbor-Jioning算法构建进化树。
1.4荧光定量引物设计及条件优化
根据GGPPS基因序列信息, Vector NTI软件设计荧光定量引物:
qRT-PCR F: AGGCACTGGAAAGGG
qRT-PCR R: AATGCACAGAACAGG
以针叶组织cDNA为模板,对模板稀释浓度(稀释3倍、5倍、8倍、10倍)、引物用量(0.1μM、0.5μM、1.0μM、5μM)及PCR扩增温度(55℃、60℃、65℃)等条件进行优化实验。反应体系按照试剂盒说明配制,PCR循环条件:95℃ 3min,95℃15sec,60℃40sec,40个循环。检测各优化条件下荧光定量扩增效果,绘制扩增曲线和熔解曲线。
2结果与分析
2.1GGPPS基因克隆
按实验方法提取总RNA并电泳检测,28S、18S条带清晰,RNA提取浓度及完整度好(图1)。

图1 总RNA提取电泳图
以cDNA为模板GGPPS基因引物进行PCR扩增,条带清晰且无非特异性杂带,得到1 000bp左右目的片段(图2)。目的片段与pMD-18T载体连接获得的阳性克隆,经菌落PCR验证,扩增如预期得到长度为1 000bp左右的片段(图3),说明重组质粒构建成功,命名为pT-GGPPS。
对pT-GGPPS的插入序列进行测序,显示克隆得到的GGPPS基因cDNA序列长度1 143bp。利用NCBI在线blast分析,结果显示序列中包含完整CDS区,与同属的辐射松(Pinusradiata,JQ262154.1)、火炬松(Pinustaeda,FJ060226.2)一致性达到99%,与松科的挪威云杉(Piceaabies,EU432050.1)、白云杉(Piceaglauca,BT118582.1)北美云杉(Piceasitchensis,BT071056.1)序列一致性达到93%以上,与北美冷杉(Abiesgrandis,AF425235.2)、海南粗榧(Cephalotaxusmannii,JX971119.1)、西藏红豆杉(Taxuswallichiana,DQ364604.1)、加拿大紫衫(Taxuscanadensis,AF081514.1)一致性也达到了82%以上,可以初步确定克隆所得片段为GGPPS基因,命名为PmaGGPPS。
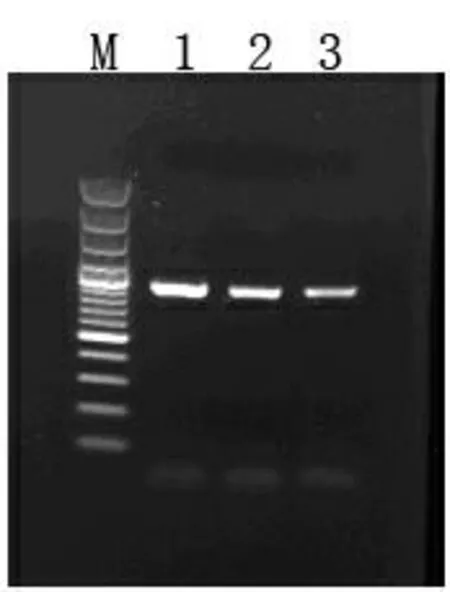
图2 目的基因cDNA片段扩增
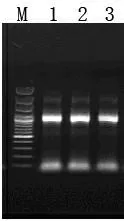
图3 阳性克隆PCR验证

图4 PmaGGPPS编码氨基酸序列保守结构域分析
应用NCBI的ORF Finder在线工具对PmaGGPPS基因编码区进行翻译,结果显示该基因编码380个氨基酸,用blastp比对氨基酸序列,结果显示序列中包含有分属异戊烯焦磷酸合成酶超家族的特征性结构域,即Trans IPPS结构域。将PmaGGPPS编码序列与其他GGPPS蛋白序列比对,结果显示在183~189位点和324~328位点具有“DDXXXXD”和“DDXXD”两个富含天冬氨酸区域,它们分别是烯丙基底物和IPP的结合位点[9]。分析结果显示,蛋白序列特点与GGPPS蛋白的酶学功能相符。
氨基酸序列比对表明,PmaGGPPS编码蛋白与Genbank上注释的挪威云杉(ACA21461.1)GGPPS蛋白序列一致性90%、北美冷杉(AAL17614.2)一致性86%,银杏(AAQ72786.1)一致性71%、加拿大紫衫(AAD16018.1)一致性69%,进一步证实克隆到的基因序列为GGPPS基因。
2.2PmaGGPPS编码蛋白序列分析
2.2.1蛋白质理化性质及结构预测
利用ExPaSy中的Protparam在线工具分析PmaGGPPS编码蛋白序列,结果显示该蛋白具有380个氨基酸残基,分子式为C1825H2950N510O549S21;相对分子量为41.49kD;理论pI值为7.56,为碱性蛋白;总平均亲水性为-0.039,不稳定系数为44.64,大于40,为非稳定性蛋白。
应用TMHMM检测PmaGGPPS编码蛋白跨膜区,并未发现跨膜结构,SignalP在线分析结果显示其也不具备信号肽序列,该蛋白不具备膜蛋白或分泌蛋白的特征结构,更可能是一种定位于细胞质基质中的游离蛋白质,这种细胞定位也与该基因的功能相符合。
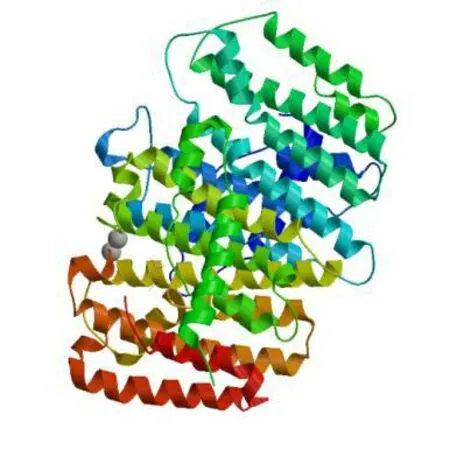
图5 PmaGGPPS编码蛋白三级结构同源建模
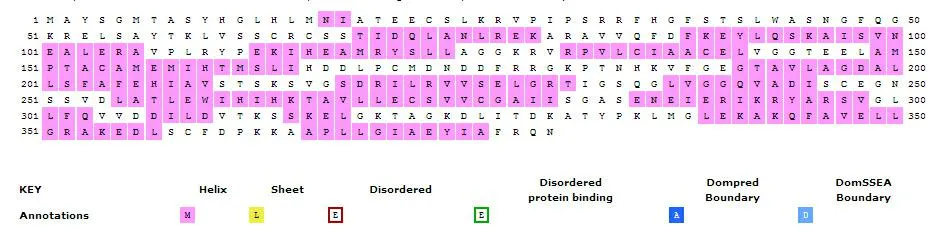
图6 PmaGGPPS编码蛋白二级结构分析
利用ExPaSy的psipred预测蛋白序列二级结构,结果与运用SWISS-MODEL对PmaGGPPS编码蛋白进行的同源三级结构建模(图5)结果相符,该蛋白的二级结构主要由16个α-螺旋组成,不含β-折叠结构(图6)。三级结构同源建模的参考模板为欧薄荷(Menthapiperita)的异聚牻牛儿基焦磷酸合酶大亚基[10],提交蛋白序列与模板覆盖率为77%,构建范围从第86个氨基酸到第380个氨基酸,序列一致性66.67%,推测PmaGGPPS编码蛋白可能具备与之相同的催化活性。
2.2.2系统进化树分析
在GenBank蛋白质数据库中检索陆生植物GGPPS蛋白质序列,共检索到53条序列,在其中选择包含完整CDS的18条序列用于比对分析。应用MEGA 5.0分析PmaGGPPS编码蛋白与这18条GGPPS蛋白序列的同源性,并构建进化树(图7)。系统进化分析显示,PmaGGPPS编码蛋白与同属松柏纲的红豆杉聚为一类,之后与银杏聚为一类,其他参比序列中,单子叶植物中的小麦和山羊草聚为一类,而除麻风树之外的其他双子叶植物聚为一类,PmaGGPPS编码蛋白所在聚类则与双子叶植物之间亲缘关系更近。

图7 PmaGGPPS系统进化树分析
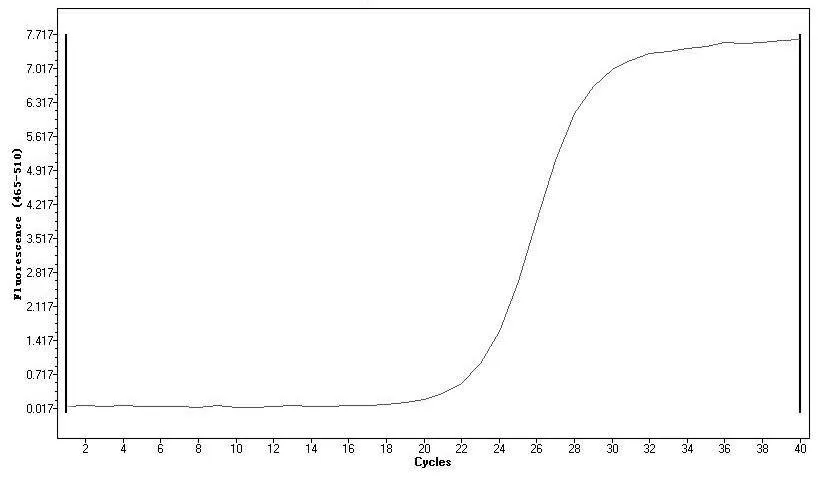
图8 荧光定量扩增曲线
2.3荧光定量引物设计及优化
以针叶组织cDNA为模板,优化反应条件并对引物进行荧光定量扩增效果检测,结果显示模板稀释8倍、引物用量为0.5μM、PCR扩增温度60℃条件下实验结果最佳,绘制的扩增曲线和熔解曲线如图8~9。熔解曲线呈现单一峰,Tm值较高,说明引物对目的基因特异性很高。扩增曲线显示信号上升拐点出现在22~24个循环范围内,30个循环之后信号基本持平,引物扩增效果较好。在优化后的实验条件下,扩增信号起峰在22~24个循环范围内,且扩增曲线也未出现明显的引物二聚体杂峰,说明优化条件下可获得较好实验效果,为开展后续实验提供技术参考。
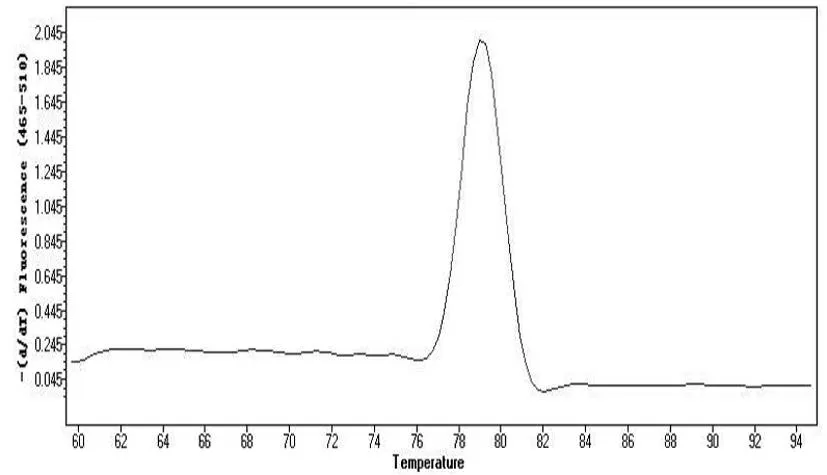
图9 荧光定量熔解曲线
3结论与讨论
3.1结论
本研究利用Genbank数据库信息,通过同源检索序列并比对分析设计引物,仅利用RT-PCR技术直接从马尾松针叶组织中克隆得到PmaGGPPS基因,并对其序列进行生物信息分析。结果显示该基因cDNA长1 143bp,开放阅读区编码380个氨基酸。PmaGGPPS编码的蛋白质序列分析显示具有Trans IPPS结构域及FARM、SARM两个富含天冬氨酸区域,序列特点与GGPPS蛋白的酶学功能相符。同源性分析结果显示PmaGGPPS核酸序列与其他两种松属植物GGPPS基因同源性达到99%以上,PmaGGPPS编码蛋白与挪威云杉等植物中GGPPS蛋白同源性也在69%以上。对PmaGGPPS编码蛋白进行理化性质及结构的生物信息学分析,结果显示该蛋白为一种无信号肽、无跨膜区的碱性蛋白质,二级结构由16段α-螺旋组成,三级结构同源建模显示与欧薄荷的异聚牻牛儿基焦磷酸合酶大亚基同源性最高。系统进化树分析显示该蛋白与红豆杉、银杏亲缘关系较近。在此基础上,根据目标基因序列设计荧光定量引物并优化反应条件,得到了理想的实验效果。
3.2讨论
目前基因克隆中大多是采用RACE技术获取基因的全长,优势是可以在通过其他实验手段仅获得目的基因部分序列信息的条件下获取全长基因,但操作步骤较为繁琐,实验周期相对较长,而在针对目的基因的功能性研究中,有时需要的仅仅是目的基因的编码区,并不需要3’和5’端非编码区序列时,此时在同源序列信息采集充分、比对分析精细甄别、引物设计质量较高的前提下,通过同源分析设计引物直接获取编码区序列则更为便捷,效率更高。
在GenBank蛋白质数据库中登录的GGPPS蛋白质序列与其他代谢途径中的关键酶相比,序列数量较少,除了用作模式植物开展研究的小麦、苜蓿外,更多的是在药材、花卉、香料植物中报道较多,比如杜仲(Eucommiaulmoides)、丹参(Salviamiltiorrhiza)、桑(MorusNotabilis)、茉莉(Jasminumsambac)、欧薄荷等。在松科植物中针对GGPPS基因的研究主要在DNA层面,涉及到蛋白活性、功能鉴定的研究工作还在起步阶段,还有一定的研究空间。
GGPPS在代谢通路中,不仅为二萜合成提供骨架,也参与类胡萝卜素、赤霉素及叶绿素侧链等的合成过程,有研究发现松树针叶中叶绿素的含量与松树产脂力存在着密切的关系,与普通马尾松类型相比较,马尾松高产脂无性系的叶绿素含量极显著提高[11]。综合GGPPS对叶绿素合成以及二萜合成中的重要作用,提示GGPPS很可能是产脂力相关基因之一,其表达水平很可能与产脂力具有相关性。
本文克隆得到的马尾松GGPPS基因,为深入研究其编码酶的酶学功能及活性提供基础,在本研究基础上,将GGPPS基因转化植物,研究其对二萜合成的影响,深入研究该基因在松脂合成中的作用,具有很大的研究空间和意义,也是本研究组下一步的研究目标。此外,本研究根据目标基因设计荧光定量引物并优化反应条件,也为下一步研究基因表达水平与产脂力关系提供了理论依据和技术参考。
参考文献:
[1]王以珊,曾令海,罗敏,等.马尾松天然林采脂试验分析[J].广东林业科技,2002,18(2):1-4.
[2]罗炼平,舒文波,聂海泉,等.马尾松土贡种源纯林与混交林生长量和产脂力比较[J].广西林业科学,2013,42(1):66-70.
[3]杨章旗.广西松脂原料林发展现状、存在问题与对策研究[J].广西林业科学,2007,36(3):143-146.
[4]安宁,丁贵杰.广西马尾松松脂的化学组成研究[J].中南林业科技大学学报,2012,32(3):59-62.
[5]杨章旗.马尾松不同年龄产脂量及松香组分分析[J].林业科学,2014,50(6):147-151.
[6]黄瑛,曾庆平.萜类生物合成的基因操作[J].中国生物工程杂志,2006,26(1):60-64.
[7]龚治,李典谟,张真.针叶树萜类合成酶研究进展[J].林业科学,2010,46(1):123-130.
[8]陈博雯,蒋华,覃鹏飞,等.尾叶桉GLU4无性系肉桂酰乙醇脱氢酶基因克隆及序列分析[J].西部林业科学,2014,43(2):19-24.
[9]Hemmi H,Noike M,Nakayama T,etal.An alternative mechanism of productchain-length determination in type III geranylgeranyl diphosphate synthase[J].Eur JBiochem,2003,270: 2186-2194.
[10]Chang TH,Hsieh FL,Ko TP,etal.Structure of a heterotetrameric geranyl pyrophosphate synthase from mint(Menthapiperita)reveals intersubunit regulation[J].Plant Cell,2010,22(2):454-467.
[11]黄益江,蔡邦平,梁一池.叶绿素含量与马尾松产脂力相关研究[J].福建林学院学报,1998,18(1):58-61.
Cloning and Bioinformatics Analysis of GGPPS of Pinus massoniana CHEN Bo-wen,QIN Zi-hai,WANG Peng-liang,JIA Jie,CHEN Hu,LIU Hai-long
(Guangxi Key Laboratory of Superior Timber Trees Resource Cultivation,Guangxi Academy of Forestry,Nanning Guangxi 530002,P.R.China)
Abstract:Geranylgeranyl diphosphate synthase(GGPPS)is the key synthase in terpenes synthesis.Its gene was cloned from the needles of Pinus massoniana seedlings by using specific primers based on the highly conserved sequences,and it was named as PmaGGPPS.Sequence analysis indicated that its cDNA sequence were 1 143 bp with its open reading frame encoding 380 amino acid residues.PmaGGPPS encoding sequence was analyzed,the result showed it has a Trans IPPS domain and two aspartate-rich motif which were FARM and SARM respectively,and the protein sequence characteristics consistent with GGPPS protein enzyme function.The results of homology analysis showed that PmaGGPPS nucleotide sequence had more than 99% sequence homology with the other two pine plants,and PmaGGPPS encoding protein had more than 69% homology with Picea abies and other plants.The physicochemical property and structure of PmaGGPPS encoding protein were analyzed by using bioinformatics tools.The result showed it was an basic protein with no signal peptide or transmembrane domain,and its secondary structure was composed of 16 alpha helix segments.Tertiary structure modeling showed it was most similar to heterotetrameric geranyl pyrophosphate synthase from mint.Phylogenetic analysis showed that it had close relation with Taxus and Ginkgo.The cloning of PmaGGPPS gene provided effective resources to further research on pine resin synthesis.This design of qRT-PCR primers according to the GGPPS gene in this study could optimize the reaction conditions,and provide theoretical basis and technical reference for next-step study on the relations of gene expression level and l pine resin production.
Key words:Pinus massoniana;geranylgeranyl diphosphate synthase(GGPPS);gene clone;qRT-PCR
中图分类号:S 791.248
文献标识码:A
文章编号:1672-8246(2016)02-0001-06
通讯作者简介:刘海龙(1980-),男,高级工程师,主要从事植物生物技术研究。E-mail:lhl-hb@sohu.com
第一作者简介:陈博雯(1983-),女,工程师,主要从事植物生物技术研究。E-mail:grfi_bwchen@163.com
基金项目:广西自然科学基金项目(2014GXNSFBA118106),广西林科院基本科研业务费项目(201419)。
*收稿日期:2015-07-23