苗期玉米叶片碳氮平衡与干旱诱导的叶片衰老之关系
2016-05-06熊炳霖王鑫月陈道钳王仕稳1殷俐娜1邓西平1
熊炳霖,王鑫月,陈道钳,王仕稳1,*,殷俐娜1,,邓西平1,
(1 中国科学院水利部水土保持研究所,黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨陵 712100;2 中国科学院大学,北京 100049;3 西北农林科技大学 水土保持研究所,陕西杨陵 712100)
苗期玉米叶片碳氮平衡与干旱诱导的叶片衰老之关系
熊炳霖1,2,王鑫月1,2,陈道钳3,王仕稳1,3*,殷俐娜1,3,邓西平1,3
(1 中国科学院水利部水土保持研究所,黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨陵 712100;2 中国科学院大学,北京 100049;3 西北农林科技大学 水土保持研究所,陕西杨陵 712100)
摘要:为了探究干旱诱导的碳氮平衡破坏与干旱诱导的叶片衰老之间的关系,该实验以8个在干旱胁迫下叶片衰老进程有明显差异的玉米品种为实验材料,采用PEG模拟干旱处理,通过测定光合速率、叶绿素含量和叶绿素荧光参数等叶片衰老指标以及非结构性碳水化合物(可溶性糖、淀粉)和全氮含量等变化,分析玉米中干旱诱导的叶片衰老与叶片中碳氮平衡(碳氮比)之间的关系。结果显示:(1)干旱胁迫下,8个玉米品种叶片净光合速率受到严重抑制,Fv/Fm大幅下降,叶绿素含量显著降低,说明干旱诱导了玉米叶片的衰老;(2)干旱诱导玉米叶片衰老的同时,8个玉米品种的叶片中可溶性糖含量显著升高,淀粉含量小幅上升,全氮含量大幅降低,碳氮比显著升高,碳氮平衡遭到了破坏;(3)8个玉米品种叶片的叶绿素含量与非结构性碳水化合物含量以及碳氮比呈极显著负相关关系,与全氮含量呈极显著正相关关系。因此,碳氮代谢与干旱诱导的叶片衰老紧密联系,碳氮平衡可能参与了干旱诱导的叶片衰老调控。
关键词:玉米;干旱;叶片衰老;碳氮平衡
干旱是农业生产的主要威胁,对农作物生长造成的损失在所有非生物胁迫中居首位,其不仅仅在干旱、半干旱区频繁发生,在非干旱地区也经常季节性发生,严重影响了农作物的正常生长和发育[1]。干旱胁迫诱导并加速作物叶片的衰老从而导致作物光合同化能力的降低和冠层面积的减少,进而引起作物产量的降低和品质的下降,干旱胁迫引起的叶片衰老是干旱造成作物减产的重要因素[2-5]。另一方面,干旱胁迫下叶片的衰老,叶绿素的降解,进而减少植物的蒸腾,是植物响应干旱胁迫的一种重要途径,是植物在严重持续干旱下提高存活率的一种适应策略[6]。
碳氮代谢是作物生长发育及产量形成的物质基础。碳水化合物和氮代谢是植物代谢以及生长发育的重要调节因子[7]。植物体内碳氮代谢紧密联系,相互影响。碳氮平衡,即细胞内碳氮代谢产物的比例,对植物代谢和生长发育也具有重要的调节作用。1904年,Klebs通过进行大量试验提出碳氮比学说,即对开花起决定性作用的不是碳水化合物和含氮化合物的量,而是其比例。当植物体内C/N比高时促进开花,反之延迟开花[8]。碳氮平衡而不是碳或氮的独立作用在调节拟南芥幼苗生长和光合基因表达过程中起主导作用[9]。除了对幼苗生长的调控作用,碳氮平衡在控制叶片衰老中也具有重要作用。Aoyama等[10]的研究结果显示,无论是在低氮培养基上添加蔗糖,还是在水培条件下控制氮浓度和提高环境CO2浓度都会加速叶片衰老症状的出现,相反,单纯的低氮或高糖环境则不能诱导衰老,该结果直接证明了碳氮平衡信号参与调控叶片衰老。在农业生产中适量施氮可以大幅提高作物产量,除了认为适量氮肥可以保证作物的氮素营养,延缓叶片衰老外,部分学者推测其保持了作物有适宜的C/N比,延缓了叶片的衰老[11-12]。
在干旱胁迫条件下,植物碳代谢受到严重干扰,光合作用受到严重抑制,碳同化速率降低[13],但可溶性糖作为重要的渗透调节物质在叶片中大量积累[14-16]。与此同时,干旱胁迫引起氮素吸收减少和氮素的重新分配,造成下部叶片氮亏缺[17-18]。目前,大多数对于干旱诱导叶片衰老的调控方面的研究都集中在细胞分裂素(ABA)和脱落酸(CK)等植物激素和活性氧(ROS)作用方面[19-20]。干旱胁迫引起作物中下部叶片可溶性糖的积累和氮素含量的降低,从而导致碳氮平衡被破坏。但是干旱诱导的碳氮平衡的破坏与干旱诱导的叶片衰老之间的关系尚不明确。我们前期的研究表明,干旱胁迫下高粱叶片碳氮平衡的破坏(表现为C/N比上升)要早于叶片衰老症状的出现,说明碳氮平衡可能参与了干旱诱导的叶片衰老调节[21]。因此,本实验以在干旱胁迫下衰老进程和耐旱能力有明显差异的8个玉米品种为材料,于水培条件下利用PEG-6000进行模拟干旱处理,测定光合速率、叶绿素含量和叶绿素荧光参数等叶片衰老指标以及可溶性糖、淀粉和全氮含量等碳氮代谢产物变化,探讨玉米中干旱诱导的碳氮平衡的破坏与干旱诱导的叶片衰老之间的关系。以期为全面了解干旱诱导叶片衰老和碳氮平衡的关系,为作物的抗旱育种和旱地农业生产提供理论论据。
1材料和方法
1.1材料
通过杨凌地区种子站购买8个本地农民常用且表型相近的玉米品种作为实验材料。供试玉米(ZeamaysL.)材料分别为‘兴民18’(为方便表述,编号为A,下同)、‘郑单958’(B)、‘德利农988’(C)、‘农乐988’(D)、‘先玉335’(E)、‘太玉339’(F)、‘晋单52号’(G)和‘并单2号’(H)8个品种。
1.2干旱处理及指标测定
实验于中国科学院水利部水土保持研究所干旱大厅人工气候室内进行。种子用1%次氯酸钠消毒20 min后,28 ℃暗中萌发4 d。种子萌发后移至盛有1/2 Hoagland营养液的塑料箱内,置于人工气候室内培养,光照强度为600 μmol·m-2·s-1,光照时间为每天14 h,昼/夜温度为28 ℃/23 ℃,相对湿度为45%~55%。幼苗生长至5叶期后,将各个品种幼苗等分成对照和干旱2组。对照组继续用1/2 Hoagland营养液培养,干旱组在1/2 Hoagland营养液中加入10% PEG-6000(-0.2 MPa)。营养液每2 d更换1次,全天通气。
PEG处理6 d后,选取第5片叶检测相关生理生化指标。对照组和干旱组各取10株地上部分先105 ℃杀青30 min,再80 ℃烘至恒重后称重。用压力室法测定叶片水势[22],根据Chen等[23]的方法利用露点渗透势仪测定渗透势;用Li-6400便携式光合系统测定气体交换参数;Imaging-PAM调制荧光仪测定光系统Ⅱ最大化学效率(Fv/Fm)[24];丙酮浸提比色法测定叶绿素含量;硫酸蒽酮比色法测定可溶性糖和淀粉含量[25];凯氏定氮法测定全氮含量。
1.3数据处理
采用SPSS 19软件进行数据统计分析,用最小显著差异法(LSD)进行单因素方差分析,显著性水平为0.05;相关性分析采用Pearson相关系数分析;SigmaPlot 12.0制图。
2结果与分析
2.1参试玉米品种幼苗的抗旱性比较
2.1.1干重经过6 d的PEG模拟干旱处理,除品种H之外,其余7个品种幼苗的地上部分干重均比对照(CK)显著降低(P<0.05);各个品种下降幅度差异明显,从60.21%(品种F)到10.79%(品种H)不等(图1,Ⅰ)。为了更直观地比较品种间耐旱能力的差异,排除各品种自身生长特性的影响,进一步计算了地上部相对干重(图1,Ⅱ)。结果表明,各品种间耐旱能力差异明显,其中的品种H地上部相对干重在干旱胁迫下维持在对照水平的89.21%,而品种F只有对照水平的39.89%,其余品种介于两者之间。说明品种H的耐旱力最强,而品种F最弱。
2.1.2叶片水势和渗透势水势是反映植物水分亏缺或水分状况的一个直接指标[26]。如图2,Ⅰ所示,在模拟干旱胁迫条件下,8个玉米品种幼苗叶片水势都比对照显著下降,且品种间差异显著(P<0.05),并以品种G叶片水势值最低(-1.006 7 MPa),而品种B最高(-0.713 3 MPa)。与叶片水势相似,8个玉米品种叶片渗透势在干旱胁迫下都比对照显著下降,品种间也差异显著,并以品种A叶片渗透势值最低(-1.153 3 MPa),而品种C最高(-0.727 8 MPa,图2,Ⅱ)。以上结果说明干旱胁迫对各个玉米品种幼苗叶片水势和渗透势影响显著,且品种间差异也较大。

*表示同一品种的处理和对照之间在0.05水平存在显著性差异;

图2 干旱胁迫下8个玉米品种叶片水势和渗透势的变化
2.2参试玉米品种幼苗叶片衰老指标的比较
2.2.1气体交换参数如图3,Ⅰ所示,8个品种幼苗叶片的净光合速率在干旱胁迫前存在一定的差异,在干旱胁迫后都比对照大幅显著下降(P<0.05);干旱胁迫后各个品种的净光合速率都处在很低的水平,品种间净光合速率的差异变小。干旱胁迫下各个玉米品种的净光合速率的降幅差异显著,如品种H的净光合速率下降幅度达85.52%,而品种G只有66.67%。与净光合速率变化趋势一致,在干旱胁迫条件下,8个玉米品种的叶片气孔导度、蒸腾速率也均比对照大幅度显著降低,并保持在很低的水平,品种间差异变小(图3,Ⅱ、Ⅲ)。

图3 干旱胁迫对8个玉米品种光合参数的影响
2.2.2PSⅡ最大光化学效率PSⅡ最大光化学效率(Fv/Fm)是PSⅡ在光合过程中潜在的最大光能转换效率,能反映胁迫对PSⅡ复合体伤害的程度。如图4所示,干旱胁迫不同程度地降低了各玉米品种幼苗的Fv/Fm。其中,品种B和E下降幅度较小,与对照组相比差异性不显著;而其它6个品种的Fv/Fm都受到干旱胁迫的显著抑制,降幅为3.70%~22.49%不等。
2.2.3叶绿素含量如图5所示,在干旱胁迫下,除品种D和品种H的叶绿素含量与对照没有显著差异外,其余6个品种叶片叶绿素含量都显著下降16.41%~48.65%,品种间差异显著(P<0.05)。同样的,干旱胁迫下各个品种的叶绿素a和叶绿素b含量都有所下降,其中,叶绿素a下降幅度为20.93%~51.28%;叶绿素b的降幅为0.71%~41.40%,可以得出叶绿素a下降幅度较叶绿素b大,从而导致叶绿素a/b显著降低,品种间差异明显。
2.3参试玉米品种幼苗叶片碳氮指标的比较
2.3.1非结构性碳水化合物含量如图6,Ⅰ所示,在干旱胁迫条件下,各玉米品种叶片可溶性糖含量都比对照不同程度升高,且除品种G外升幅都达到显著水平(P<0.05),品种间也差异明显;干旱胁迫对各玉米品种叶片淀粉含量的影响相对较小,品种A、C、D和F均显著高于对照,而其余品种与对照差异不显著(图6,Ⅱ);同时,在干旱胁迫条件下,与叶片可溶性糖含量表现相似,各品种叶片的非结构性碳水化合物(TNC)含量均不同程度比对照升高,且除品种G外升幅(21.2%~58.9%)均达到显著水平(P<0.05),且TNC升高幅度表现出明显的品种间差异(图6,Ⅲ)。其中,叶片TNC升高幅度以品种G最低(21.2%),而品种E最高(58.9%)。以上结果说明干旱胁迫导致了各玉米品种叶片中可溶性糖和淀粉的积累。

图4 干旱胁迫对8个玉米品种PSⅡ最大

图5 干旱胁迫对8个玉米品种叶绿素含量的影响
2.3.2全氮含量干旱胁迫会引起烟草[17]和羊草[18]氮素吸收减少和氮素的重新分配,造成其下部叶片氮亏缺。本研究结果(图7,Ⅰ)显示,8个玉米品种叶片全氮含量也在干旱胁迫下比对照都显著下降(P<0.05),品种D下降幅度最小(23.8%),品种H降幅最大(44.1%),并以品种D全氮含量在干旱胁迫下最高(26.36 mg·g-1),而以品种H全氮含量最低(20.55 mg·g-1)。

图6 干旱胁迫对8个玉米品种非结构性
2.3.3碳氮比如图7,Ⅱ所示,各个玉米品种叶片碳氮比于正常水分条件下维持在3.57~4.44之间,而在干旱胁迫条件下升高至6.73~10.04之间。在干旱胁迫下,由于叶片TNC的升高和全氮含量的降低,导致8个品种叶片碳氮比都显著升高(P<0.05),且品种间差异变大。
2.4各玉米品种叶片衰老指征与碳氮平衡相关分析
在干旱胁迫条件下,玉米叶片衰老指征叶绿素含量、叶绿素a/b(图5,Ⅰ、Ⅱ)和Fv/Fm都显著下降(图4),非结构性碳水化合物含量显著升高(图6,Ⅲ),全氮含量显著降低(图7,Ⅰ),碳氮比显著升高(图7,Ⅱ)。但是各项指标在干旱胁迫下的变化幅度表现出明显的品种间差异。为了明确叶片衰老与碳氮平衡之间的关系,将指征叶片衰老程度的叶绿素含量分别与TNC、全氮含量和碳氮比做相关分析。结果(图8)显示,玉米叶片TNC和碳氮比与其叶绿素含量之间均呈极显著的负相关,而全氮含量与叶绿素含量之间呈极显著的正相关。
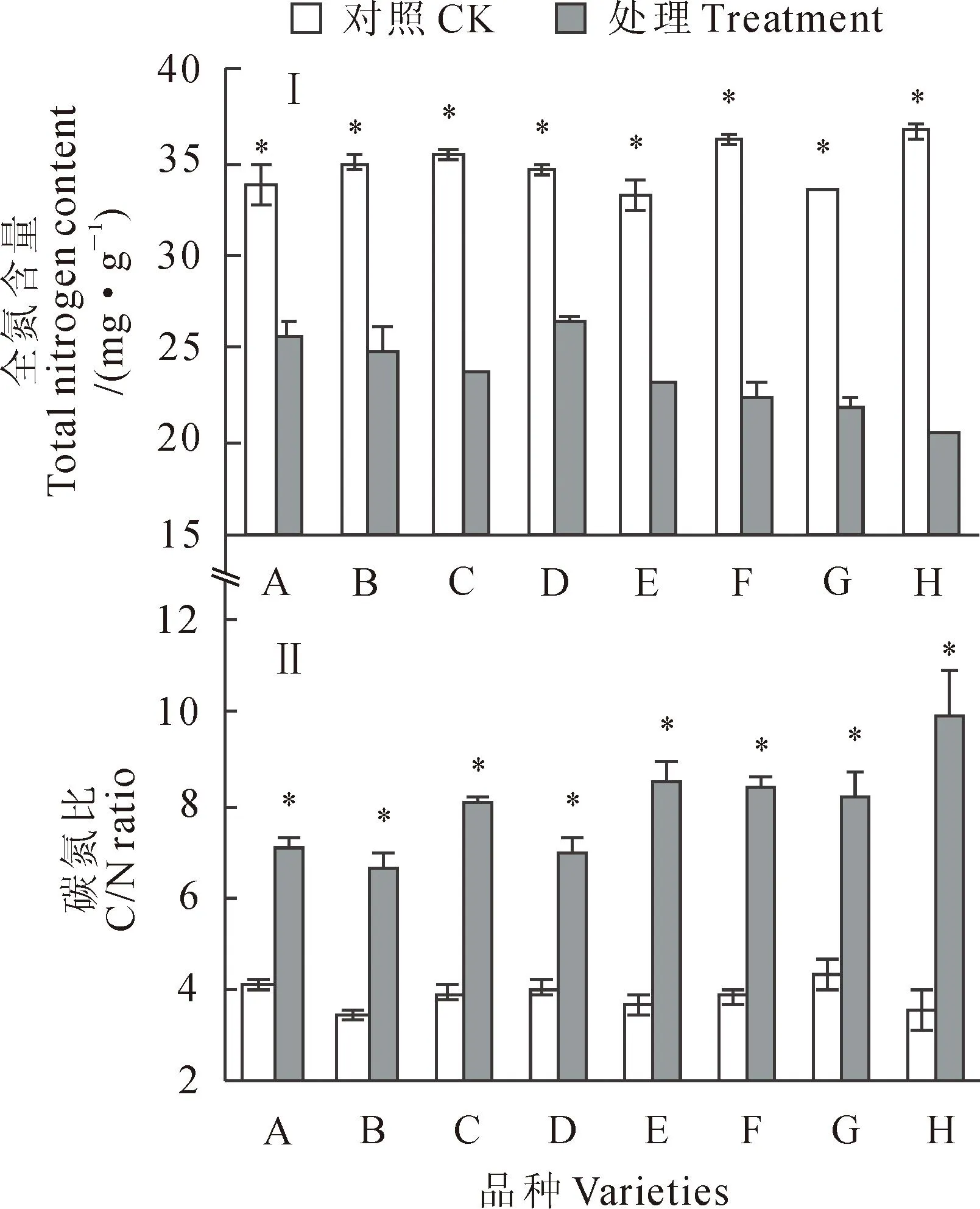
图7 干旱胁迫对8个玉米品种全氮含量和碳氮比的影响
3讨论
叶片衰老是由内部遗传因素和外部环境共同控制的高度有序的过程,是植物细胞程序性死亡(programmed cell death,PCD)的一种类型。主要表现为光合速率和Fv/Fm下降,叶绿素、蛋白质和脂质等大分子分解[27]。环境胁迫,如干旱、高温、盐等因素会诱导和促进叶片的衰老[3]。本研究中,在干旱胁迫下,8个玉米品种叶片净光合速率受到严重抑制,Fv/Fm大幅下降,叶绿素含量和叶绿素a/b值显著降低,这些结果都表明干旱诱导了玉米叶片的衰老。尽管干旱胁迫下8个玉米品种的衰老指标都表现出显著降低的趋势,但是各个品种的下降幅度呈现出较大的差异,如8个玉米品种叶片Fv/Fm、叶绿素含量和叶绿素a/b的下降幅度分别为3.70%~22.49%、16.41%~48.65%和8.25%~27.45%。这说明玉米不同品种对干旱诱导的叶片衰老的响应具有明显的差异。

图8 叶片衰老指征与碳氮平衡相关分析
干旱胁迫条件下,作物碳氮代谢受到严重干扰。干旱胁迫会引起叶片中可溶性糖含量的积累和氮素含量的降低,从而导致下部叶片碳氮平衡的破坏。
在本研究中,在干旱胁迫下,8个玉米品种叶片的可溶性糖含量显著升高,淀粉含量不变或小幅度升高;叶片全氮含量大幅降低;由于非结构性碳水化合物(TNC)含量的升高和全氮含量的降低,引起8个品种中叶片碳氮比都显著升高。这与以往相关研究结果一致[13-18],说明在干旱胁迫下,玉米叶片的碳氮代谢受到严重干扰,叶片碳氮平衡被破坏。另外,干旱胁迫下各个玉米品种叶片碳氮比升高的幅度大小明显不同,说明不同玉米品种碳氮代谢对干旱胁迫的响应存在品种间差异。
碳氮代谢是作物生长发育及产量形成的物质基础,且植物体内碳氮代谢紧密联系,相互影响。碳氮平衡,即细胞内碳氮代谢产物的比例,对植物代谢和生长发育也具有重要的调节作用。Aoyama等[10]研究结果显示碳氮平衡,而不是碳或氮的独立作用在调节叶片衰老过程中起主导作用。为了明确干旱诱导的叶片衰老与碳氮平衡之间的关系,我们将指征叶片衰老程度的叶绿素含量分别与TNC、全氮含量和碳氮比做相关分析。结果显示叶绿素含量和TNC以及碳氮比呈极显著的负相关,而与全氮含量呈极显著的正相关。这一结果表明,碳氮代谢在干旱诱导的叶片衰老中紧密联系,碳氮平衡可能参与了干旱诱导的叶片衰老调控。
目前,大多数关于干旱诱导叶片衰老的调控研究都集中在细胞分裂素(CK)和脱落酸(ABA)等植物激素和活性氧(ROS)作用的等方面[19-20]。本研究结果表明碳氮平衡也可能参与干旱诱导的玉米叶片衰老的调控。但是碳氮平衡调控叶片衰老的机制尚不清楚,有待进一步明确。细胞分裂素(CK)能有效延缓叶片衰老[28],如干旱胁迫下维持较高的CK水平可以延缓干旱诱导的叶片衰老并提高植物抗旱能力[19]。而最近的研究表明,干旱胁迫下维持较高的CK水平是通过碳氮代谢的协调控制来提高抗旱能力[29]。碳氮代谢的协调是碳氮平衡的基础。ABA能加速叶片的衰老[30]。Wingler等[31]提出植物对非生物胁迫的响应可能依赖于糖信号和ABA以及它们的相互作用。另外,很多ABA合成和信号组分同样是糖信号通路中的重要组分[32]。以上研究表明,碳氮平衡可能通过与CK和ABA的相互作用来调节干旱诱导的叶片衰老过程。
本研究采用在干旱胁迫下叶片衰老进程有明显差异的8个不同玉米品种进行试验,这8个玉米品种的耐旱性存在显著差异。在本研究中叶片衰老程度和作物耐旱能力呈现负相关,即叶片衰老越严重,其耐旱能力越强。由于实验中测定的是下部叶片,而且实验是在一个持续严重干旱的过程中进行,并不包括复水的过程,在持续严重干旱胁迫下,下部叶片碳氮平衡的破坏诱导其衰老,从而减少水分耗散和促进养分向上部幼嫩组织转移,进而提高植物耐旱能力。同时在本研究中,碳氮比和水势变化呈现相反的趋势,干旱胁迫后水势越低的品种其碳氮比越高,而与渗透势变化趋势关系不明显,说明碳氮比的升高可能不是植物主动渗透调节的结果。
参考文献:
[1]CHAVES M M,PEREIRA J S,MAROCO J,etal.How plants cope with water stress in the field.Photosynthesis and growth[J].Ann.Bot.,2002,89:907-916.
[2]MUNNE-BOSCH S,ALEGRE L.Die and let live:leaf senescence contributes to plant survival under drought stress[J].Funct.PlantBiol.,2004,31(3):203-216.
[3]ZHANG H S,ZHOU C J.Signal transduction in leaf senescence[J].PlantMol.Biol.,2013,82(6):539-545.
[4]BORRELL A K,HAMMER G L,HENZELL R G.Does maintaining green leaf area in sorghum improve yield under drought?Ⅱ.Dry matter production and yield[J].CropSci.,2000,40:1 037-1 048.
[5]RIVERO R M,KOJIMA M,GEPSTEIN A,etal.Delayed leaf senescence induces extreme drought tolerance in a flowering plant[J].Proc.Natl.Acad.Sci.USA,2007,104(49):19 631-19 636.
[6]安玉艳,梁宗锁.植物应对干旱胁迫的阶段性策略[J].应用生态学报,2012,23(10):2 907-2 915.
AN Y Y,LIANG Z S.Staged strategy of plants in response to drought stress[J].ChineseJournalofAppliedEcology,2012,23(10):2 907-2 915.
[7]CORUZZI G M,ZHOU L.Carbon and nitrogen sensing and signaling in plants:emerging ‘matrix effects’[J].Curr.Opin.Plan.,2001,4(3):247-253.
[8]曲波,张微,陈旭辉,等.植物花芽分化研究进展[J].中国农学通报,2010,26(24):109-114.
QU B,ZHANG W,CHEN X H,etal.Research progress of flower bud differentiation mechanism of plant[J].ChineseAgriculturalScienceBulletin,2010,26(24):109-114.
[9]MARTIN T,OSWALD O,GRAHAM I A.Arabidopsis seedling growth,storage lipid mobilization,and photosynthetic gene expression are regulated by carbon:nitrogen availability[J].PlantPhysiol.,2002,128(2):472-481.
[10]AOYAMA S,HUARANCCA R T,GUGLIELMINETTI L,etal.Ubiquitin ligase ATL31 functions in leaf senescence in response to the balance between atmospheric CO2and nitrogen availability inArabidopsis[J].PlantCellPhysiol.,2014,55(2):293-305.
[11]王利纳,陶红斌,戴明宏,等.包膜尿素对夏玉米产量及碳氮代谢的影响[J].玉米科学,2009,17(2):124-129.
WANG L N,TAO H B,DAI M H,etal.Effect of coated urea on grain yield and metabolism of summer maize[J].JournalofMaizeSciences,2009,17(2):124-129.
[12]刘海坤,刘小宁,黄玉芳,等.不同氮水平下小麦植株的碳氮代谢及碳代谢与赤霉病的关系[J].中国生态农业学报,2014,22(7):782-789.
LIU H K,LIU X N,HUANG Y F,etal.Effect of nitrogen application on carbon and nitrogen metabolisms and relationship between carbon metabolism and wheat scab[J].ChineseJournalofEco-Agriculture,2014,22(7):782-789.
[13]CHAVES M M.Effects of water deficits on carbon assimilation[J].J.Exp.Bot.,1991,42:1-16.
[14]WANG Z,QUEBEDEAUX B,STUTTE GW.Osmotic adjustment-effect of water-stress on carbohydrates in leaves,stems and roots of apple[J].Aust.J.PlantPhysiol.,1995,22(5):747-754.
[15]NIO S A,CAWTHRAY G R,WADE L J,etal.Pattern of solutes accumulated during leaf osmotic adjustment as related to duration of water deficit for wheat at the reproductive stage[J].PlantPhysiol.Biochem.,2011,49(10):1 126-1 137.
[16]REGUERA M,PELEG Z,ABDEL-TAWAB Y M,etal.Stress-induced cytokinin synthesis increases drought tolerance through the coordinated regulation of carbon and nitrogen assimilation in rice[J].PlantPhysiol.,2013,163(4):1 609-1 622.
[17]FERRARIO-MERY S,VALADIER M H,FOYER C H.Overexpression of nitrate reductase in tobacco delays drought-induced decreases in nitrate reductase activity and mRNA[J].PlantPhysio.,1998,117(1):293-302.
[18]XU Z Z,ZHOU G S.Nitrogen metabolism and photosynthesis inLeymuschinensisin response to long-term soil drought[J].J.PlantGrowthRegul.,2006,25(3):252-266.
[19]YANG J C,ZHANG J H,WANG Z Q,etal.Involvement of abscisic acid and cytokinins in the senescence and remobilization of carbon reserves in wheat subjected to water stress during grain filling[J].PlantCellandEnviron,2003,26(10):1 621-1 631.
[20]KHANNA-CHOPRA R.Leaf senescence and abiotic stresses share reactive oxygen species-mediated chloroplast degradation[J].Protoplasma,2012,249(3):469-481.
[21]CHEN D Q,WANG S W,XIONG B L,etal.Carbon/nitrogen imbalance associated with drought-induced leaf senescence inSorghumbicolor[J].PlosOne,2015,10(8):e0137026.
[22]LIU P,YIN L N,DENG X P,etal.Aquaporin-mediated increase in root hydraulic conductance is involved in silicon-induced improved root water uptake under osmotic stress inSorghumbicolorL[J].J.Exp.Bot.,2014,65(17):4 747-4 756.
[23]CHEN D Q,YIN L N,DENG X P,etal.Silicon increases salt tolerance by influencing the two-phase growth response to salinity in wheat (TriticumaestivumL.)[J].ActaPhysiol.Plant,2014,36(9):2 531-2 535.
[24]XU W Z,DENG X P,XU B C,etal.Photosynthetic activity and efficiency ofBothriochloaischaemumandLespedezadavuricain mixtures across growth periods under water stress[J].ActaPhysiol.Plant,2014,36(4):1 033-1 044.
[25]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006:144-148.
[26]王丁,姚健,杨雪,等.干旱胁迫条件下6种喀斯特主要造林树种苗木叶片水势及吸水潜能变化[J].生态学报,2011,31(8):2 216-2 226.
WANG D,YAO J,YANG X,etal.Changes of leaf water potential and water absorption potential capacities of six kinds of seedlings in Karst mount area under different drought stress intensities:taking sink forestation seeding in karst Mountainous region for examole[J].ActaEcologicaSinica,2011,31(8):2 216-2 226.
[27]LIM P O,KIM H J,NAM H G.Leaf senescence[J].Annu.Rev.PlantBiol.,2007,58:115-136.
[28]ZHANG H S,ZHOU C J.Signal transduction in leaf senescence[J].PlantMolecularBiology,2013,82(6):539-545.
[29]CUTLER S R,RODRIGUEZ P L,FINKELSTEIN R R,etal.Abscisic acid:emergence of a core signaling network[J].Annu.Rev.PlantBiol.,2010,61:651-679.
[30]JUVANY M,MULLER M,MUNNE-BOSCH S.Leaves of field-grown mastic trees suffer oxidative stress at the two extremes of their lifespan[J].J.Integr.PlantBiol.,2012,54(8):584-594.
[31]WINGLER A,PURDY S,MACLEAN J A,etal.The role of sugars in integrating environmental signals during the regulation of leaf senescence[J].J.Exp.Bot.,2006,57(2):391-399.
[32]ROLLAND F,BAENA-GONZALES E,SHEEN J.Sugar sensing and signalling in plants:conserved and novel mechanisms[J].AnnualReviewofPlantBiology,2006,57:675-709.
(编辑:裴阿卫)
Carbon/Nitrogen Balance Associate with Drought-induced Leaf Senescence in Maize (Zeamays) Seedling
XIONG Binglin1,2,WANG Xinyue1,2,CHEN Daoqian3,WANG Shiwen1,3*,YIN Lina1,3,DENG Xiping1,3
(1 State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau,Institute of Soil and Water Conservation,Chinese Academy of Sciences,and Ministry of Water Resources,Yangling,Shaanxi 712100,China;2 University of Chinese Academy of Science,Beijing 100049,China;3 Institute of Soil and Water Conservation,Northwest A&F University,Yangling,Shaanxi 712100,China)
Abstract:To investigate the association between drought-induced carbon/nitrogen imbalance and leaf senescence,we treated eight maize varieties in leaf senescence progress under drought stress with PEG to mimic the drought stress.Leaf senescence symptoms,such as photosynthesis,photosystem II photochemistry efficiency (Fv/Fm) and chlorophyll content,and the non-structural carbohydrates (soluble sugars,starch) content and nitrogen content were determined.Then,the correlation between drought-induced leaf senescence and carbon/nitrogen imbalance (C/N ratio) was analyzed.Our results showed that:(1)the net photosynthetic of eight maize varieties were severely suppressed,Fv/Fm has fallen dramatically and chlorophyll content decreased significantly,which indicated that drought induced maize leaf senescence.(2)Drought stress induced the maize leaf senescence accompanied with the accumulation of soluble sugar content,slightly increased starch content,decreased of total nitrogen content and increased significantly of C/N ratio,carbon and nitrogen balance were damaged.(3)Both the non-structural carbohydrates content and carbon/nitrogen ratio showed a significant negative correlation with chlorophyll content,and there was a significant positive correlation between total nitrogen content and chlorophyll content.Therefore,carbon and nitrogen metabolisms were tightly associated with drought-induced leaf senescence,and the carbon/nitrogen balance may involve in the regulation of drought-induced leaf senescence.
Key words:maize;drought;leaf senescence;carbon/nitrogen balance
中图分类号:Q945.79
文献标志码:A
作者简介:熊炳霖(1990-),女,在读硕士研究生,主要从事旱地作物生理生态的研究。E-mail:1210722732@qq.com*通信作者:王仕稳,副研究员,主要从事植物生理生态以及分子生物学研究。E-mail:shiwenwang@nwsuaf.edu.cn
基金项目:国家科技支撑计划(2015BAD22B01);国家重点基础研究发展计划(973项目,2015CB150402)
收稿日期:2015-11-06;修改稿收到日期:2016-03-01
文章编号:1000-4025(2016)03-0534-08
doi:10.7606/j.issn.1000-4025.2016.03.0534
