不同施氮水平下甜菜光合特性比较
2016-04-24史树德
刘 莹,史树德
(内蒙古农业大学农学院,内蒙古呼和浩特 010018)
在甜菜生长过程中,氮作为肥料三元素之首,对其生长发育有极其重要的作用,施氮水平会极大影响叶片的光合能力。光合作用是甜菜块根产糖量形成的重要影响因素,在甜菜块根产量中,90%~95%的有机质是由光合作用所固定并转化的[1-2]。众多学者研究认为植物光合作用受水分、温度等多种外部因素影响,特别是群体光合作用受冠层结构影响较大[3-4]。施氮水平会对甜菜的冠层结构产生重要影响。作物产量及品质的形成,既决定于作物的遗传性状,也受土壤水分[5]、矿质元素状况[6]、CO2浓度、温度及湿度等外部环境的影响[7]。如果氮素不足,通常表现出叶绿素含量较低,光合能力较弱。施氮过多,甜菜产质量不但不能提高,反而呈下降趋势[8]。因此,适量施氮可以提高甜菜光合能力,同时提高其干物质积累能力,并对品质产生积极影响。
本试验设计5个施氮梯度,分别为0,50,100,150,200 kg/hm2,以甜菜KWS9167为试验对象,通过对叶绿素含量、SPAD值和光合特性,以及产量和含糖率等指标的研究,以探究不同施氮条件下甜菜光合能力的差异以及相关性,为甜菜农业生产提供理论依据和参考生理指标。
1 材料和方法
1.1 试验设计
试验于2015年5-10月于内蒙古农业大学科研实习农场进行,土壤类型为壤土,耕作土壤肥力中等。
试验设 5 个施氮水平,分别为 0,50,100,150,200 kg/hm25个施氮水平,3次重复,随机区组排列。供试肥料为尿素(含N46%),以基肥40%、追肥60%的比例进行施氮,追肥于叶丛生长期进行。磷肥和钾肥分别施以过磷酸钙(P2O5含量为18%)和硫酸钾(K2O含量为50%),按200 kg/hm2施肥,以基肥的形式一次施用。
供试甜菜品种为KWS9167。试验于5月1日人工播种,10月13日收获,试验田管理同大田。种子采取穴播方式,株距、行距为25 cm×50 cm。
1.2 测定指标及方法
从长出第六片真叶时(6月5日)开始取样,每隔20 d左右在晴朗天气的9:00—11:00时,每处理随机取3株,对其完全展开的功能叶片(倒4叶)进行光合速率和SPAD的测定,然后将整株取回。选取光合速率测定叶片,先测定SPAD值,再去叶脉剪碎混匀以测定叶绿素含量。
含糖率测定:以糖量计测定(数据矫正采用《手持糖量计读数的温度修正表》进行矫正)。
光合速率值测定:每小区随机选取3株甜菜,用LCpro-SD测定倒4叶片光合速率、气孔导度、胞间CO2浓度和蒸腾速率,测定时间为9:00—12:00时。
SPAD值测定:采用日本产SPAD-502型叶绿素仪对甜菜倒2叶的顶部进行测定[9]。
叶绿素的测定:取LCpro-SD和SPAD测定后的对应叶片取回实验室,去掉叶脉后剪碎,分别称取0.5 g用80%丙酮研磨提取叶绿素,并以80%丙酮作为空白对照,分别测定663 nm和645 nm处的吸光度并计算叶绿素浓度。具体方法参见张志良(2009年第四版)《植物生理学实验指导》[10]。
1.3 数据分析
采用Excel和SPSS处理软件进行数据分析。
2 结果与分析
2.1 不同施氮条件下甜菜叶片叶绿素变化情况
由图1可知,不同施氮水平下,叶绿素整体趋势均表现出单峰曲线,所有处理苗期最低,然后增加,在8月6日(块根膨大期)前后达到高峰,而后又逐渐降低。不同施氮条件下存在差异,含量最低点出现在N0的苗期,然后逐渐升高,最高点出现在8月6日N150水平下,不同处理间的变化趋势相似。
在苗期(5月30日)不同施氮条件下甜菜的叶绿素浓度依次为:N200>N50>N0>N150>N100,N150与其他4个处理间差异显著(P<0.05);叶丛生长期前期(6月20日)叶绿素含量大小依次为:N150>N100>N50>N200>N0,N200与其他处理间差异显著(P<0.05);叶丛生长期后期(7月11日)叶绿素含量大小依次为N150>N100>N200>N50>N0,各处理间均存在差异且差异较明显(P<0.05);块根膨大期(8月6日和8月26日)叶绿素含量大小均为:N150>N200>N100>N50>N0,N0和N200与其他处理间存在显著差异(P<0.05);糖分积累期(9月23日)叶绿素含量大小为:N150>N100>N200>N50>N0,其中N200与N150间存在差异,与其他处理间存在显著差异(P<0.05);收获时(10月13日)叶绿素含量为:N150>N100>N50>N200>N0,其中N50与其他处理间存在显著差异(P<0.05)。
2.2 不同施氮条件下甜菜叶片SPAD值变化情况
由图2可看出,不同施氮水平下,甜菜的SPAD值从苗期到糖分积累期整体呈现先上升后下降的趋势。到8月6日(糖分积累前期)前后,N150水平下达到最大值,其次为N200水平下的SPAD值,最低点出现在苗期。
不同施氮条件下甜菜苗期的SPAD值大小依次为:N0>N150>N50>N200>N100;叶丛生长期(6月20日至7月11日),SPAD值呈上升趋势,不同施氮条件下甜菜的SPAD值大小依次为:N100>N50>N150>N200>N0,N0与其他各处理间差异显著(P<0.05);块根膨大期(8月6日和8月26日),不同施氮条件下甜菜的SPAD值大小均为:N150>N200>N100>N50>N0,块根膨大初期(8月6日)各处理间均存在显著差异(P<0.05),其中N150最高;块根膨大后期(8月26日),N50与其他各处理间差异显著(P<0.05);糖分积累期(9月23日),不同施氮条件下甜菜的SPAD值大小为:N150>N200>N100>N50>N0,N50和 N0与其余各处理间差异显著(P<0.05);收获时(10月13日),N0与其他处理间差异显著(P<0.05)。
2.3 不同施氮条件下甜菜叶片光合速率变化情况
从图3可以看出,不同施氮水平下甜菜叶片光合速率在叶丛生长期和块根膨大期,叶片光合速率均呈现上升趋势,达到最高值后则呈现下降趋势。苗期各处理的光合速率最低,最高点出现在N150的块根膨大期初期(8月6日)前后,到糖分积累期则呈下降趋势。
苗期,不同施氮条件下甜菜的光合速率大小依次为:N100>N150>N200>N50>N0,各处理间的差异不显著;叶丛生长初期(6月20日),不同施氮条件下甜菜的光合速率大小依次为:N100>N50>N150>N200>N0,N100与其他各处理间存在显著差异(P<0.05);叶丛生长后期(7月11日),不同施氮条件下甜菜的光合速率大小依次为:N150>N100>N50>N200>N0,N150与 N0间存在显著差异(P<0.05),与其他处理间存在差异;块根膨大前期(8月6日),不同施氮条件下甜菜的光合速率大小依次为:N150>N200>N100>N50>N0,各处理间不存在显著差异;块根膨大后期(8月26日),不同施氮条件下甜菜的光合速率大小依次为:N200>N100>N50>N150>N0,N0与N200存在显著差异,与其他处理间存在差异;糖分积累期(9月23日),不同施氮条件下甜菜的光合速率大小依次为:N200>N100>N0>N50>N150,N200与N100和N150间存在显著差异,与N0和N50间存在差异;收获时(10月13日),不同施氮条件下甜菜的光合速率大小依次为:N200>N0>N150>N100>N50,N200与其他处理间存在显著差异(P<0.05)。
2.4 不同施氮条件下甜菜叶片蒸腾速率变化情况
由图4可知,不同施氮条件下,甜菜蒸腾速率呈现前期较低、中期较高、后期又降低的变化趋势。苗期,N150和N0与其他处理间存在显著差异(P<0.05);叶丛生长前期(6月20日),各处理间不存在显著差异;叶丛生长末期(7月11日)大小依次为:N150>N100>N200>N0>N50,N150与其他处理间存在显著差异(P<0.05);块根膨大初期(8月6日)大小依次为:N150>N100>N200>N50>N0,N150与 N0间存在显著差异(P<0.05);块根膨大末期(8月26日)大小依次为:N150>N100>N50>N200>N0,N150与N200和N50间存在显著差异(P<0.05);糖分积累期(9月23日)各处理间不存在显著差异;收获时(10月13日)大小依次为:N150>N200>N50>N0>N100,N150与N100间存在显著差异(P<0.05)。
2.5 不同施氮条件下甜菜叶片胞间CO2浓度变化情况
由图5可知,甜菜不同施氮水平下叶片胞间CO2浓度的变化趋势相类似,均呈现单峰曲线趋势。最高点出现在N100水平下的7月11日(叶丛生长末期),最低点出现在苗期。苗期,N0与N200间存在显著差异(P<0.05),与其他处理间也存在差异;叶丛生长初期(6月20日),不同施氮水平下甜菜叶片胞间CO2浓度大小为:N0>N50>N100>N150>N200,N0和N150与N200间存在显著差异(P<0.05),与其他处理间也存在差异;叶丛生长末期(7月11日),各处理间差异不显著;块根膨大初期(8月6日),不同施氮水平下甜菜叶片胞间CO2浓度大小为:N50>N0>N100>N150>N200,N50 与 N150、N200之间差异显著(P<0.05);块根膨大末期(8月26日),不同施氮水平下甜菜叶片胞间CO2浓度大小为:N50>N100>N150>N200>N0,N100、N150 和 N200之间并没有差异,但与N0和N50存在显著差异(P<0.05);糖分积累期(9月23日),不同施氮水平下甜菜叶片胞间CO2浓度大小为:N50>N0>N100>N150>N200,N100与 N50间存在显著差异(P<0.05);收获时(10月13日),不同施氮水平下甜菜叶片胞间CO2浓 度 大 小 为 :N0>N50>N100>N200>N150,N150、N200和N0间差异显著(P<0.05)。
2.6 不同施氮条件下甜菜叶片气孔导度变化情况
从图6可以看出,不同施氮条件下甜菜叶片气孔导度生育前期较低,中期较高,后期呈现平稳下降的变化规律,在叶丛生长后期(7月11日)达到最高。苗期,不同施氮条件下甜菜叶片气孔导度大小依次为:N150>N100>N50>N200>N0,N150 与 N50、N200和N0间差异显著(P<0.05);叶丛生长期(6月20日至7月11日),各处理间不存在显著差异(P<0.05);块根膨大前期(8月6日),不同施氮条件下甜菜叶片气孔导度大小依次为:N200>N150>N0>N100>N50,N50与N0、N150和N200间存在显著差异(P<0.05);块根膨大后期(8月26日)和糖分积累期(9月23日),不同施氮条件下甜菜叶片气孔导度大小分别为:N150>N100>N50>N200>N0、N150>N100>N200>N50>N0,差异性均为N150与N0间存在显著差异(P<0.05);收获时(10月13日),各处理间差异不显著(P<0.05)。
2.7 不同施氮水平对甜菜产量和含糖率的影响
通过表1可知,氮素水平对甜菜块根产量、含糖率和产糖量会产生不同影响。随施氮量的增加,甜菜块根产量呈上升趋势,并在N150处达到最大,块根产量的大小依次为:N150>N200>N100>N50>N0。N150条件下,块根产量为66970 t/hm2,与对照N0相比,增长了58.4%,两者间差异显著(P<0.05)。含糖率呈现相类似的特征表现,随施氮量的增加,甜菜含糖率呈上升趋势,并在N100处达到最大,含糖率的大小依次为:N100>N150>N50>N200>N0。N100条件下,甜菜含糖率为16.766%,与对照N0相比,提高了2.4百分点,两者间差异显著(P<0.05)。产糖量的最高点出现在N150处,达到10.459 t/hm2,与对照N0相比,增长了72.4%,两者间差异显著(P<0.05)。
2.8 不同施氮条件下光合特性等指标相关性分析
表2结果表明:施氮量与叶绿素含量达到极显著正相关水平(R=0.686**),与SPAD值、光合速率、产量和产糖量呈显著正相关水平(R=0.548*、R=0.518*、R=0.533*和 R=0.571*)。叶绿素含量与 SPAD值和产糖量达到极显著正相关(R=0.642**和R=0.684**),与含糖率和产量呈显著正相关(R=0.585*、R=0.640*)。SPAD值与含糖率、产量和产糖量呈显著正相关(R=0.605*、R=0.545*和 R=0.611*)。光合速率与产量呈显著正相关(R=0.580*)。蒸腾速率与胞间CO2浓度呈显著负相关(R=-0.516*)。产量和产糖量呈极显著正相关(R=0.984**)。
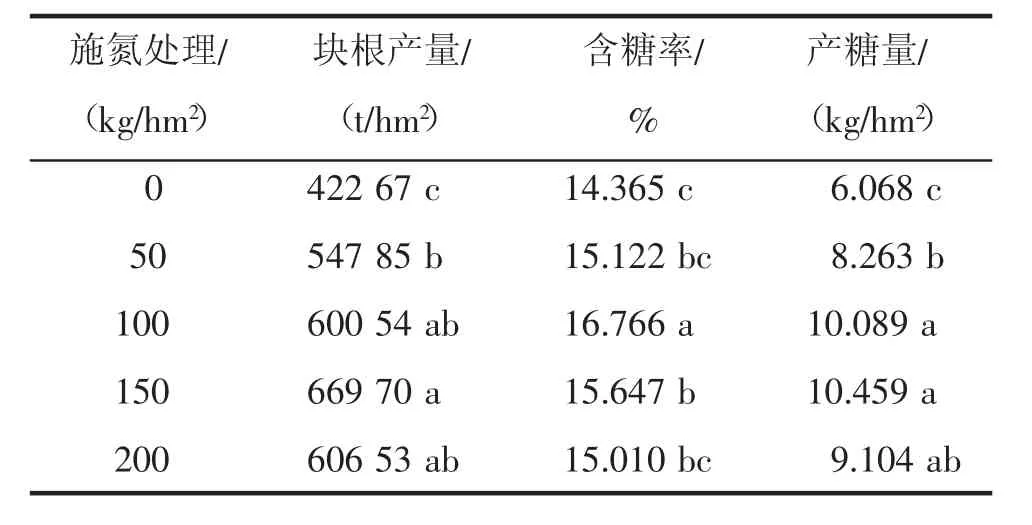
表1 不同施氮水平甜菜块根产量、含糖率和产糖量
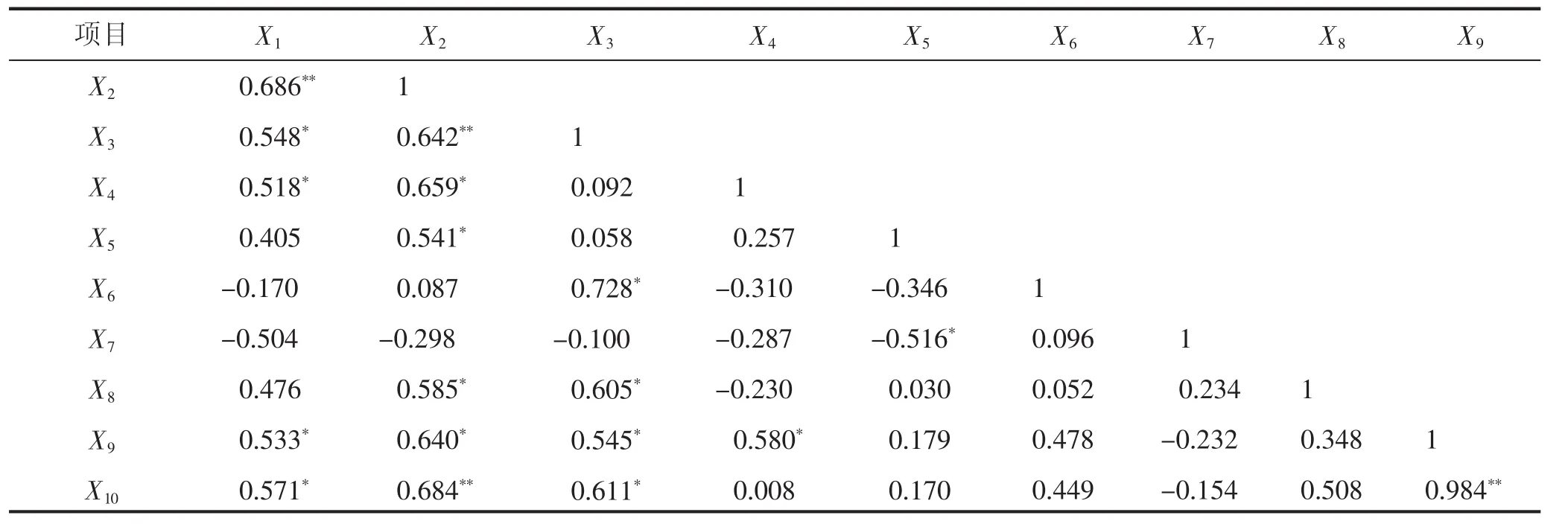
表2 不同施氮条件下各项指标相关性
3 讨论
施氮水平会对甜菜产质量产生重要影响,通过以上相关性分析可知:在甜菜生育期内,氮素水平与甜菜产量和产糖量呈显著正相关。最大产量和最大产糖量为N150水平下,最大含糖率出现在N100水平下,过量施氮时,甜菜产质量会随施氮量的增加而下降,并且会对环境产生影响[11]。由此表明,氮素水平调控对产量及含糖率有重要影响,适量施氮能增加产量,但过量施氮会对品质造成不利影响[12]。
随着叶片的生长,不同施氮条件下,叶片叶绿素含量呈单峰曲线增长,在块根膨大前期(8月6日)N150处理下达到最大值。
SPAD值和光合速率的变化规律也与叶绿素变化规律相似。由此表明,氮素水平对甜菜光合作用产生重要影响,当施氮量达到150 kg/hm2后,增施氮量对甜菜光合作用的促进作用不明显,甚至会降低。
通过相关性可以发现,在甜菜生长过程中,叶绿素含量、SPAD值、光合速率、蒸腾速率和施氮量之间表现为正相关,此特点与郭连旺[13]等人的研究结果相似。胞间CO2浓度与其他指标表现为负相关。叶绿素含量和SPAD值与块根产量、含糖量表现为显著正相关。光合速率与块根产量表现为显著正相关。
通过此次试验表明,叶绿素含量、SPAD值和光合速率的大小会对甜菜产量和品质产生重要影响。作物产量以及品质的形成,取决于多方面因素的影响,包括土壤水分[5]、矿质元素的营养情况[14]等外部生态因子[15],同时优良品种以及合理的种植密度和栽培方式也会对作物本身光合作用以及氮素同化作用产生重要影响[16]。因此,综合各方面因素,为甜菜生长创造一个适宜生长的环境,对甜菜增产和提高品质有重要意义。
参考文献:
[1]曲文章.甜菜生理学[M].哈尔滨:黑龙江科学技术出版社,1990.146-220.
[2]杜永成.氮磷钾使用量对甜菜光合能力和氮代谢酶的影响[D].哈尔滨:东北农业大学,2012.
[3]周健朝.甜菜不同品种光合生产能力的研究[J].中国糖料,2001(3):14-18.
[4]于海滨,周 芹,刘 娜,等.不同氮磷营养水平对甜菜叶片光合速率的影响[J].中国糖料,2001(3):19-21.
[5]夏 冰,阳树英,刘清波.生态因子对水稻叶片光合生理功能的影响综述[J].作物研究,2008(2):140-142.
[6]王月福,陈建华.土壤水分对小麦产量和品质的影响[J].莱阳农学院学报,2002(1):7-9.
[7]越 鹏,李彩凤,陈叶婷,等.氮素水平对甜菜功能叶片光合特性的影响[J].核农业学报,2010,24(5):1080-1085.
[8]薛鸿雁,公丽凤,张文成,等.氮素不同用量对甜菜产质量的影响[J].中国糖料,2004(2):28-29.
[9]陈 旭.甜菜内源激素与其抗丛根病的关系[D].呼和浩特:内蒙古农业大学,2011.
[10]张志良,瞿伟菁,李小方.植物生理学实验指导[M].北京:高等教育出版社,2009.58-60.
[11]Gary W,Hergert.Sugar beet fertilization[J].Sugar Technol ogy,2010,12(3/4):255-266.
[12]高妙真.甜菜根重与含糖率相关关系的研究[J].中国甜菜,1995(1):16-19.
[13]郭连旺,沈允钢.高等植物光合机构避免强光破坏的保护机制[J].植物生理学通讯,1996,32(1):1-8.
[14]姜东燕,于振文.土壤水分对小麦产量和品质的影响[J].核农学报,2007,21(6):641-645.
[15]Chen Genyun,Yong Zhenhua,Liao Yi,et al.Photosynthetic acclimations in rice leaves to free-air CO2enrichment related to both ribulose-1,5-bisphosphate carboxylation limitation and ribulose-1,5-bisphosphate regeneration limitation[J].Plant Cell Physiology,2005,46(7):1036-1045.
[16]Refay Y A.Root yield and quality traits of three sugar beet(Beta vulgaris L.)varieties in relation to sowing date and stand densities[J].World Journal of Agricultural Sciences,2010,6(5):589-594.
