杉木不同无性系主要经济性状变异分析
2016-04-17段红静郑会全胡德活张馨文张心悦孙宇涵
段红静 曹 森 郑会全 胡德活 林 军 张馨文 张心悦 陈 鹏 孙宇涵 李 云
(1.北京林业大学林木育种国家工程实验室,北京林业大学生物科学与技术学院,北京100083;2.广东省林业科学研究院,广东广州510520;3.广东省乐昌市龙山林场,广东乐昌512221)
杉木不同无性系主要经济性状变异分析
段红静1曹 森1郑会全2胡德活2林 军3张馨文1张心悦1陈 鹏1孙宇涵1李 云1
(1.北京林业大学林木育种国家工程实验室,北京林业大学生物科学与技术学院,北京100083;2.广东省林业科学研究院,广东广州510520;3.广东省乐昌市龙山林场,广东乐昌512221)
对广东省乐昌市龙山林场杉木基因库中272个10年生杉木无性系的树高、胸径、树皮厚、单株材积4个生长性状,心材比、木材基本密度等2个材质性状进行测定分析。结果表明:272个杉木无性系间存在丰富的表型变异,表型变异系数均>10%;以单株材积变异系数最大,为69.04%,遗传变异系数为73.97%,变异幅度0.003 8~0.479 1m3。6个重要性状在无性系间均表现为极显著差异,表明所测各性状均受较强的遗传控制。遗传相关系数和表型相关系数均是材积最高;密度遗传变异系数最低,为13.03%,重复力属中上水平(0.684 3),仅心材比的重复力低于0.5。在10%、20%、30%和50%不同入选率下计算,所测性状遗传增益值均随入选比例的增大而降低。树高、胸径、树皮厚、材积4个生长性状间呈显著正相关,且均与木材基本密度呈极显著负相关。
杉木;无性系;变异;生长性状;材质性状
遗传变异研究是林木育种的重要前提条件。丰富的遗传变异,可以增强对环境的适应,扩大遗传基础,加速林木的育种进程[1-4]。基因型与环境相互作用导致林木表型的表达。表型性状的多样性是生物多样性的表现形式之一,掌握植物重要性状的变异情况及变化规律,对遗传资源的保护及育种利用具有重要的意义[5-7]。
杉木(Cunninghamia lanceolata)是杉科杉木属的一种常绿针叶树种,为我国特有,具有速生、材质优良且无明显病虫害等特点[8-10],是一种重要的经济用材树种,广泛分布于我国南方的广东、福建、浙江等17省,研究表明,杉木种源间个体生长性状及材质性状存在着比较丰富的变异[11-14]。杉木的遗传改良研究始于20世纪50年代,研究水平一直处于国内领先地位,通过种源试验的开展、种子园的营建、无性系的选育、多世代遗传改良等的研究,大量丰富的育种资源以种子园、种质资源库和子代林等各种方式得到了保存,目前,杉木的第3代种子园已建成。为了使现有的育种水平得到持续的提高,合理评价和利用现存的杉木育种资源,加快育种进程,需要丰富多样的种质遗传资源。本研究对保存于龙山林场的广东省内272个10年生杉木优树无性系进行树高、胸径、树皮厚、单株材积、心材比、木材基本密度等性状的测定,解释其生长与材质性状的遗传变异规律及相关关系,以期为广东杉木优树育种资源的保存和开发利用提供依据,并为杉木多性状综合选择提供参考。
1 材料与方法
1.1 试验材料
试验材料选自保存在龙山林场的10年生杉木无性系,分别由乐昌、怀集、信宜等广东省内9个林分的优树无性繁殖而来,优树基因型共272个,每个基因型嫁接4株,随机分布,该嫁接无性系种植在龙山林场基因库中,株行距3 m×3 m,基因库所在地海拔451m,西北向,地处东经113°28′,北纬25°13′。林区属亚热带粤北气候区,全年气候温和,雨量充沛,为杉木的生长发育提供了较为适宜的生长环境。
1.2 重要表型性状的测定
对多个生长性状和材质性状进行测量记录,每个基因型测定3个单株,作为3次重复,共816株杉木。树高、胸径和树皮厚3个生长性状的数据在龙山林场直接测量获得。树高(m)采用测杆测量,胸径(cm)用测树围尺测定,树皮厚(mm)是在树干胸径处上坡方向剥下1块3 cm×2 cm截面大小至能见到边材的树皮进行测定。通过对优树单株树高、胸径的测定计算材积[15];在胸径处,利用树木生长锥钻取优树木芯,装于未完全密封的塑料管中标记保存,随后进行材质测定。心材比即心材边缘到髓心的长度为半径的圆盘面积除以整个圆盘面积所得的比值,通过测量所取木芯的心材长度和边材长度进行折算(心材是指木本茎干最内的颜色通常较深的木材,边材是指树木次生木质部的外围活层);木材基本密度的测定采用饱和含水率法[15-17],将取回来的样品在水中浸泡(每隔2天换水1次)至恒质量,称取湿质量后使用型号为BJYSL-DHG-9030A的实验室用大型烘箱(90℃温度下)将其烘干至恒质量,取出称其干质量。单位均为g。
1.3 数据统计分析
材积计算公式:
V=0.000 058 777 042×D1.9699831×H0.89646157
式中:V、D和H分别表示单株材积、胸径及树高。
木材基本密度:ρ=1/(Gmw/Gh-0.346)
式中:Gmw表示试样饱和含水质量;Gh表示绝干样质量。
重复力:R=1-1/F
式中:F为方差分析的F值;
遗传增益Δ=S×R
式中:S为选择差;R为重复力。
数据统计分析结合Microsoft Excel 2010和Statistical Analysis System(SAS V8.1)进行,其中方差分析应用Duncan’s检验。
2 结果与分析
2.1 杉木优树无性系生长性状和材质性状的差异
对保存于龙山林场的广东省内9个不同试验林来源的干型通直、整枝良好且无明显病害的272个基因型进行测定,并对全部单株间及无性系间的表型变异系数进行计算。在群体水平上,各无性系树高、胸径、树皮厚、单株材积等生长性状和心材比、木材基本密度等材质性状的差异状况见表1。

表1 优树无性系生长性状和材质性状的差异Tab.1 The difference situation about growth and timber characteristics of plus tree clones
由表1可知,广东省内被调查的优树无性系生长性状中单株材积变异幅度最大,为0.003 8~0.479 1m3;估算的无性系间表型变异系数在生长性状中也最高,达69.04%。其次为胸径,变异幅度为2.3~27.0 cm,最大值与最小值间比值为11.7。变异幅度最小的是树皮厚,最大值与最小值间比值为5,对应于无性系间的表型变异系数最低,为17.32%。杉木无性系的心材比和木材基本密度存在不同程度的变异,总体变幅分别为0.96%~63.06%和0.124 0~0.577 8 g/cm3,最大值与最小值间比值分别为65.7和4.5,其中无性系间心材比表型变异系数高达29.82%,而木材基本密度表型变异系数为13.03%。单株间表型变异系数均大于无性系间变异系数,但差异都不大,说明无性系单株间变异水平较低,表明环境对性状的影响不大。
2.2 杉木优树无性系表型性状的方差及相关性分析
对杉木无性系表型性状进行统计分析,各测定性状无性系间均呈极显著的差异(表2)。进一步作方差分量估算,其结果(表3)表明,6个表型性状均具有明显的无性系方差分量,其中树高性状无性系方差分量占总方差分量的百分数最高,达65.8%;胸径、材积、树皮厚方差分量占总方差分量的百分比均>50%,分别为63.0%、60.0%和56.5%;6个性状中,心材比方差分量所占方差总量的百分比最低,为20%。这些方差分量表明,所测杉木的表型性状虽然受一定程度环境的影响,但受明显的遗传控制。优树无性系间生长与材质性状存在着广泛的变异,这些表型性状上的变异为选择优良基因型个体提供了可能。
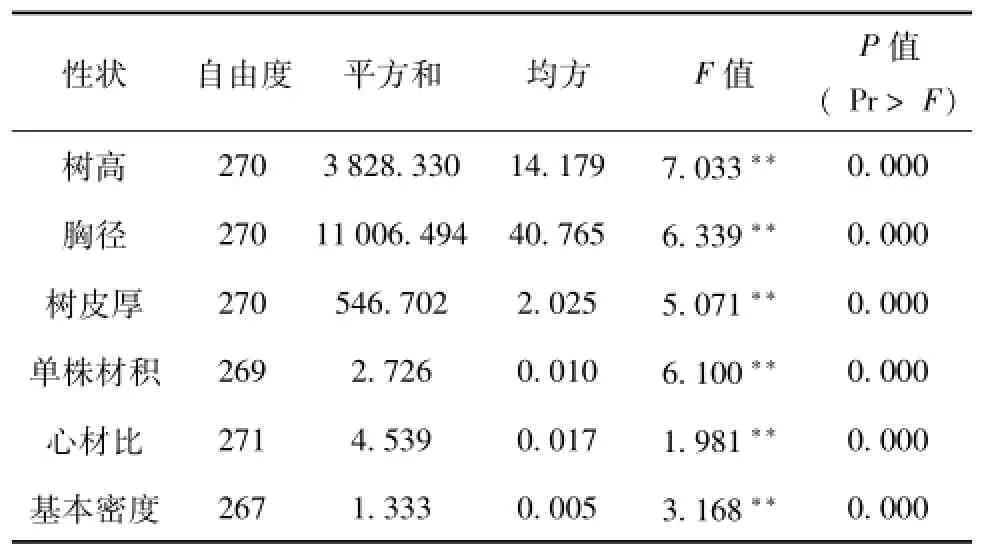
表2 杉木无性系生长和材质性状方差分析Tab.2 The variance analysis results of growth and timber characteristics of Chinese fir clones

表3 杉木无性系生长和材质性状方差分量估算Tab.3 The variance component estimation of the growth and timber characteristics of Chinese fir clones
对杉木无性系各性状遗传参数进行估算,结果见表4。由表4可知,材积的遗传变异系数最高,为73.97%,重复力为0.836 1,属较高水平;密度的遗传变异系数最低,为17.49%,但其重复力较心材比高,为0.684 3,属中上水平。心材比的遗传变异系数较树高、胸径、树皮厚和密度均高,为47.20%,但重复力最低,为0.495 2。对杉木无性系在10%、20%、30%和50%入选率状况下进行计算,各性状的遗传增益估算值均有所变化,所测性状遗传增益值均随入选比例的增大而降低,10%的入选率下,胸径的遗传增益值最高,心材比的遗传增益值最低,为0.10%;其次为密度,遗传增益值为0.11%。
广东杉木优树无性系性状间的相关性分析见表5。对广东杉木无性系生长性状和材质性状间的表型相关系数进行估算后发现,4个生长性状间呈紧密的正相关,均达极显著水平。其中,树高与胸径的正相关水平最高,相关系数达0.833 25,其次为树皮厚,树高与材积的相关性水平最低,为0.434 48。4个生长性状与心材比均呈正相关,除了材积与心材比相关性不显著外,其他性状均与心材比呈极显著的正相关,而心材比与密度也呈极显著的正相关,但是,所有的生长性状均与木材基本密度呈极显著的负相关,其中,胸径与密度的负相关程度最高,最低的是心材比。尽管树高与心材比在表型上达极显著的正相关,但是在遗传水平上二者之间的正相关水平不明显,另外,材积与心材比在表型上相关性不明显,但在遗传水平上二者成极显著的正相关,其他各性状间的遗传相关性和表型相关性一致。

表4 杉木无性系生长和材质性状遗传参数估算Tab.4 The genetic parameter estimation of the growth and timber characteristics of Chinese fir clones

表5 杉木无性系性状间的相关关系Tab.5 The correlation between traits of Chinese fir clones
2.3 杉木优树无性系的再选择
对不同林分来源无性系的分析结果见表6。由表6可知,广东省林业科学研究院来源的优树在树高、胸径和单株材积方面性状优势较为明显,树高平均值为10.35 m,胸径平均值为16.10 cm,材积平均值为0.133 9 m3。龙山林场来源的优树树皮厚的平均值则高于其他来源优树群体,为5.10 mm,胸径平均值为16.10 cm,与广东省林业科学研究院来源的优树胸径平均值相同。综合各性状发现,广东省林业科学研究院和龙山林场来源的优树群体的生长表现最为突出。
此外,综合胸径和密度2个性状,选出>胸径平均值的无性系共126个,其中32个无性系(52,95,124,138,160,182,184,202,211,255,277,368,388,415,429,436,586,596,605,625,629,630,666,669,673,692,697,698,705,708,723,726)的基本密度在平均密度值以上,这些优良无性系中有10个来自于广东省林业科学研究院和龙山林场。

表6 各来源优树生长性状差异情况Tab.6 Differences of growth traits of superior trees
3 结论与讨论
遗传变异的研究,对植物资源的保护和可持续利用具有十分重要的意义。保存于龙山林场杉木基因库中的广东省9个不同试验林10年生嫁接无性系杉木,表型性状具有丰富的变异,但各性状变异程度有所不同,其中以材积性状变异程度最大,所测重要性状在无性系间均表现为极显著差异,其中心材比的无性系方差分量占总方差分量的百分比仅为20%,即心材比受环境影响的强度较大,但是差异显著性最小。本试验中杉木单株随机分布,因此,这些表型性状上的变异表明,广东杉木可能存在较大程度的遗传变异。杉木丰富的表型变异,为选择优良基因型个体提供了依据,为杉木育种资源的改良提供了可能,本研究结果综合生长与材质性状选择出一批优良杉木无性系。杉木生长性状是可以进行早期预测的[18],施季森等的研究表明,杉木幼龄材与成熟材的性状是密切相关的,利用10年生左右幼龄材的密度预测成熟材的密度与管胞长度是可行的[19]。因此,在杉木的遗传改良工作中,本研究对10年生杉木嫁接无性系进行的表型性状多样性的研究,为广东成熟材材质的预测提供了依据。
生长与材质性状相关性的研究对杉木育种群体的遗传改良具有重要意义。有研究表明,杉木生长与材质性状间是微弱的负相关或者无关[11,19-22],而部分研究则表现为中等强度以上的负相关[10,23-25]。本研究对广东种源10年生杉木优树的生长和材质性状间的相关系数进行估算后发现,4个生长性状间呈紧密的正相关,并分别与心材比呈极显著或微弱的正相关,但是,所有的生长性状均与木材基本密度呈极显著的负相关,其中,胸径与密度的负相关程度最高,而密度是衡量木材材性的重要指标之一。因此,本研究综合胸径和密度两性状进行二维选择,选择出多个生长与材质性状兼优的优良无性系,为优良种源的收集与保存提供来源。
杉木是重要的速生材性树种,不同基因型个体在表型性状上存在着不同程度的变异,本研究从表型性状方面分析广东杉木的变异性,还需要进一步开展分子水平的研究,以期从表型和分子2个层次对广东杉木优树资源出作综合评价。
[1] Nevo E,Chen G.Droughtand salt tolerances in wild relatives for wheat and barley improvement[J].Plant,Cell&Environment,2010,33(4):670-685.
[2] Hoffmann A A,SgròCM.Climate change and evolutionary adaptation[J].Nature,2011,470(7335):479-485.
[3] Du Q Z,Pan W,Xu B H,et al.Polymorphic simple sequence repeat(SSR)loci within cellulose synthase(PtoCesA)genes are associated with growth and wood properties in Populus tomentosa[J].New Phytologist,2013,197(3):763-776.
[4] Matuszewski S,Hermisson J,Kopp M.Catch me if you can:Adaptation from standing genetic variation to amoving phenotypic optimum[J].BioRxiv,2015:015685.
[5] 姚淑均,张守攻,王军辉,等.滇楸花部性状的表型多样性研究[J].中南林业科技大学学报,2013,33(10):20-24.
[6] 张强英,张启翔,程堂仁.榆叶梅野生群体表型多样性的研究[J].中南林业科技大学学报,2012,32(5):156-160.
[7] 文亚峰,韩文军,吴顺.植物遗传多样性及其影响因素[J].中南林业科技大学学报,2010,30(12):80-86.
[8] 陈岳武,施季森.杉木遗传改良中的若干基本问题[J].南京林业大学学报(自然科学版),1983,25(4):5-18.
[9] 郑会全,胡德活,王润辉,等.杉木速生无性系选择与材质分析[J].中国农学通报,2012,28(10):27-31.
[10] 郑会全,梁瑞友,胡德活,等.杉木大径材优树选择与主要经济性状变异分析[J].西南林业大学学报,2012,32(4):26-29.
[11] 施季森,叶志宏,翁玉臻,等.杉木生长与材性联合遗传改良研究[J].南京林业大学学报,1993,17(l):l-8.
[12] 韦如萍,胡德活,郑会全,等.杉木无性系生长和材性变异及多性状指数选择[J].林业科学,2012,48(3):45-50.
[14] 齐明,何贵平,李恭学,等.杉木不同水平试验林的遗传参数估算和高世代育种的亲本评选[J].东北林业大学学报,2011,39(5):4-8.
[15] 成俊卿.木材学[M].北京:中国林业出版社,1985:663-467.
[16] GB/T 1931—2009,木材含水率测定方法[S].北京:中国标准出版社,2009.
[17] GB/T 1933—2009,木材密度测定方法[S].北京:中国标准出版社,2009.
[18] 叶培忠,陈岳武.杉木早期选择的研究[J].南京林产工业学院学报,1981,5(l):106-116.
[19] 施季森,叶志宏,陈岳武,等.木材材性的遗传和变异研究II杉木种子园自由授粉子代间木材密度的遗传变异和性状之间的相关性[J].南京林业大学学报,1987,11(4):15-24.
[20] 胡伯智,冯建国.杉木优良家系实生苗与扦插苗造林效果比较[J].南京林业大学学报(自然科学版),1999,23(4):74-75.
[21] 李晓储,吕祥生.杉木生长与材性兼优无性系的选择[J].林业科技开发,2002,16(6):12-14.
[22] Zheng H,Hu D,Wang R,et al.Assessing 62 Chinese Fir(Cunningham ia lanceolata)Breeding Parents in a 12-Year Grafted Clone Test[J].Forests,2015,6(10):3799-3808.
[23] 林同龙.杉木杂交后代胸径生长和木材体积质量的遗传变异及联合选择[J].浙江林学院学报,2000,17(2):142-145.
[24] 朱栗琼.杉木无性系材性变异研究[D].南宁:广西大学,2003.
[25] 胡德活,阮梓材,钱志能,等.杉木无性系木材密度遗传变异及其与生长性状的相关性[J].中南林学院学报,2004,24(5):24-27.
(责任编辑 张 坤)
Variation Analysis on the Main Economic Characters of Chinese Fir Clones
Duan Hongjing1,Cao Sen1,Zheng Huiquan2,Hu Dehuo2,Lin Jun3,Zhang Xinwen1,Zhang Xinyue1,Chen peng1,Sun Yuhan1,Li Yun1
(1.National Engineering Laboratory for Forest Tree Breeding,College of Biological Sciences and Technology,Beijing Forestry University,Beijing 100083,China;2.Guangdong Academy of Forestry,Guangzhou Guangdong510520,China;3.Longshan Forest Farm in Lechang of Guangdong Province,Lechang Guangdong 512221,China)
272 ten years old of Chinese fir clones at Longshan farm,Lechang city of Guangdong provenance,4 phenotypic traits including tree height,diameter at breast height,bark thick,volume of timber,2 wood property characteristics including the heartwood ratio and density of wood were investigated.The results showed that there were abundant phenotypic variation in 272 clones.The phenotypic variation coefficient were all above 10%,of which,volume of timbre's was the biggest.The volume ranged from 0.003 8m3to 0.479 1 m3with the phenotypic and genetic variation coefficientwas69.04%and 73.97%,respectively.Very significant difference among clones of 6 measured traits and there were all variance component in clones which suggested a genetic control of them. Further study found that the variation tendency of genetic variation coefficient followed the phenotypic variation coefficient.The density presented the lowest genetic variation coefficientof13.03%,while had a relative high repeatability value(0.684 3)and only the heartwood ratio was lower than 0.5.All themeasured traits,the absolute ge-netic gain increased under lower selection ratio(10%,20%,30%and 50%).Among the 6 phenotypic traits,4 growth traits showed a significant relation between each other and all were significantly negative correlation related to wood basic density.
Chinese fir;clone;variation;growth traits;wood properties

S722.3
A
2095-1914(2016)02-0078-06
10.11929/j.issn.2095-1914.2016.02.013
2015-12-15
国家林业局重点项目(2012-06)资助;国家“863”重点项目(2011AA100203)资助;林业公益性行业科研专项(201404127)资助。
第1作者:段红静(1988—),女,博士生。研究方向:林木遗传育种与生物技术。Email:duan673356712@126.com。
孙宇涵(1983—),男,博士,讲师。研究方向:林木遗传育种与生物技术。Email:sunyuhan@bjfu.edu.cn。
