5种栽培模式对油茶土壤微生物及酶活性的影响
2016-04-17冯金玲郑新娟杨志坚陈世品
冯金玲 郑新娟 杨志坚 陈世品 陈 辉
(福建农林大学林学院,福建福州350002)
5种栽培模式对油茶土壤微生物及酶活性的影响
冯金玲 郑新娟 杨志坚 陈世品 陈 辉
(福建农林大学林学院,福建福州350002)
以毛豆、花生、绿豆、四季豆套种及覆草为试验栽培模式,探讨不同栽培模式对油茶、土壤微生物及土壤酶活性的影响。结果表明:5种栽培模式对油茶新梢生长、单株产果量及出油率有显著影响,但对鲜出籽率影响不明显;对土壤微生物数量和酶活性均存在显著差异,根际土壤微生物数量和酶活性比非根际土壤高;综合分析得出在油茶林下套种毛豆和绿豆2种模式最好。细菌与真菌、各种土壤酶活性以及出油率均有显著或极显著相关关系,说明土壤细菌可作用为判断土壤肥力状况的生物学指标。
油茶;根际;土壤;微生物;酶活性;套种
油茶(Camellia oleifera)为山茶科(Theaceae)山茶属(Camellia)木本油料植物,分布广泛,栽培历史已有2 300多年的历史。茶油是优质食用油,富含油酸和亚油酸等不饱和脂肪酸,含量占90%以上。油茶一经种植,连年收益,效益长达百年之久。但油茶秋花秋果,抱子怀胎,树体营养消耗大。并且油茶林长期掠夺式经营,粗放管理,导致养分大幅度递减,土壤板结,处于低产状态[1]。因此,急需提高油茶林栽培管理工作水平。采用生态栽培模式,林农套种,以耕代抚、以耕代管;充分发挥土地效益,既可节省大量抚育和管护经费,又可增加经济收入。油茶生态栽培已在产区采用,幼林套种豆科作物有利于油茶的生长发育[2-3],提高土壤养分含量,改善油茶土壤微生物的群落结构,增加各微生物物菌群的生物量[4]。其中间种大豆(Glycinemax)能提高油茶林地土壤的N、P、K含量[5];套种豌豆(Pisum sativum)提高土壤碱解氮含量[6];套种决明(Catsia tora)提高土壤细菌、真菌、N、P、K含量及含水率[4,7];而套种花生(Arachis hypogaea)除了提高经济效益外,也提高了土壤的磷含量[8]。同时油茶幼林覆草也能提高土壤理化性状,且效果随着覆草量的增加而显著增加[9];除此之外,油茶幼林下套种茶和红薯(Ipomoea batatas)等也都有利于油茶的生长[10-11]。但成年油茶林下套种研究较少,目前只见到成熟林下套种生姜(Zingiber officinale)的研究,且主要集中生态栽培的油茶产量以及经济效益等方面,而对地下部的微生物和酶活性等的影响还很缺乏研究[12]。
土壤系统和植被是一个有机整体,二者相辅相成。土壤系统为植被生长提供必需的营养物质,而植被生长又可改善土壤系统的结构和养分[13]。根际是由植物根系与土壤微生物之间相互作用所形成的独特圈带,是植物生长过程中形成的复杂、动态的微型生态系统,对植物生长发育起着至关重要的作用[14]。土壤微生物直接参与有机质分解,腐殖质形成,养分转化和循环的各个生化过程,是土壤生态系统变化的预警及敏感指标[15-16]。土壤酶活性是土壤生物活性和土壤肥力的重要指标,其中蔗糖酶、磷酸酶、脲酶等水解酶的总体活性对评价土壤肥力水平更具有重要意义[17]。因此,在研究不同生态栽培模式对油茶成熟林根际和非根际土壤效应时,探讨由此引起的土壤微生物和酶活性的动态变化具有重要意义。本试验以在20年油茶林下套种毛豆(Glycine max)、花生(Arachis hypogaea)、绿豆(Vigna radiate)、四季豆(Phaseolus vulgaris)以及覆草为材料,研究不同栽培模式下油茶根际微生物及酶活性的差异及变化特征,以期揭示不同生态栽培模式对油茶林生产的作用机理。
1 材料与方法
1.1 试验区自然概况
试验地设在福建省闽侯桐口国有林场的华南油茶良种工区,该地地处北纬26.05°,东经119.17°,海拔260~300 m。年平均气温19.6℃,最冷月(1月)平均气温10.2℃,最热月(7月)平均气温28.7℃,年降水量1 342.5 mm,相对湿度77%,年积温6 457℃,日照时数1888 h,无霜期326 d。试验地为东南坡向,坡度为23°左右。
1.2 试验设计
选东南方向、中坡的普通油茶林地为试验地,油茶林郁闭度为0.65左右。林内油茶树龄均为20 a,长势良好。试验地共设6个处理,18个小区,随机区组排列。每个小区12株,面积为100 m2,周围匀设保护行。处理1~4分别套种毛豆、花生、绿豆、四季豆,处理5为覆草,处理6作为CK。
处理前,先对油茶林全面松土,套种密度以作物生长后能够覆盖地表为宜。2009年3月初次播种,条播。处理1,矮脚毛豆株距30~35 cm,每穴3~4粒;处理2,花生株距15~20 cm,每穴2粒;处理3,绿豆株距约25 cm,每穴5~6粒;处理4,四季豆株距35~40 cm,每穴3粒;处理5,五节芒覆盖厚度10 cm;处理6作空白CK。试验在2010—2012年再连续种植3次。
1.3 土样的采集
于2012年的4、7、10月和12月4次分别取样。取样法为“S”形混合法,在每个小区内设5个样点,每个采样点距离油茶植株30 cm,除去表面的土层和枯枝败叶,在0~20 cm土层内挖取油茶的主根,沿着主根找到根系末梢的须根,抖落须根上大块土壤(不含根系)作为非根际土壤,附着在根系表面的细粒土壤(<4 mm)作为根际土壤。将5个样点的土样混合后用四分法取1 kg,作为各个小区分析土壤微生物的样土放入无菌采样袋,封口、贴上标签,带回实验室,并于4℃冰箱保存,备用。剩余的部分放置实验室阴凉通风处进行自然风干,风干1周后过1 mm筛进行土壤酶活性分析。
1.4 土壤测定指标及方法
土壤微生物数量的测定,采用梯度稀释法制备土壤悬液,稀释涂抹平板计数法进行分离,测定细菌、真菌、放线菌数量,3次重复。细菌培养基为牛肉膏蛋白胨培养基,细菌采用10-5浓度进行涂板,28℃培养4 d后计数;真菌培养基为马丁氏-孟加拉线培养基,真菌采用10-3浓度进行涂板,28℃培养3 d后计数;放线菌培养基为改良高氏1号培养基,放线菌采用10-4浓度进行涂板,28℃培养8 d后计数[18]。
土壤脲酶、酸性磷酸酶、蔗糖酶、和过氧化氢酶活性,分别采作苯酚钠-次氯酸钠比色法、磷酸苯二钠比色改进法[19]、3,5-二硝基水杨酸比色法、0.021mol/L KMnO4滴定法测定[20]。以3次测定结果的平均值为测定值,所有数据都通过测定各自的土壤含水量换算为每克烘干土的含量。
1.5 植物测定指标及方法
2012年6月中旬选择每个小区土壤5个取样点的5株植株为样树,每株树随机选30枝新梢测定长度。2012年10月下旬采集每个小区全部植株的果实,称质量,取株数平均值为单株产果量,从中随机挑选100个果实,用电子天平测定鲜果重和鲜籽质量,鲜出籽率=鲜籽质量/鲜果质量。种仁含油率采用索氏抽提法测定[21]。
1.6 数据处理
采用Excel软件对数据进行处理,用DPS统计软件进行方差分析、多重比较(LSD法,α=0.05)和相关分析。
2 结果与分析
2.1 不同栽培模式对油茶林生长及产量的影响
由表1可知,不同栽培模式对油茶新梢生长、单株产果量及出油率有显著影响,但对鲜出籽率的影响不明显,5种栽培模式中油茶新梢生长及产量都比CK高。5种栽培模式中,新稍长度按大小排列为:花生、绿豆>毛豆>四季豆>覆草,比CK分别提高了39.8%、22.4%、15.3%、2.0%。其中花生和绿豆的新梢平均生长长度为13.7 cm,覆草的长度为10.0 cm。单株产果量按大小排列为:花生>毛豆>绿豆>四季豆>覆草,比CK分别提高了173.9%、104.8%、82.1%、46.3%、19.0%。其中花生栽培中油茶单株产果量为9.928 kg,覆草的为4.313 kg,鲜出籽率中各个栽培模式变化不大,毛豆、花生和绿豆都是为0.40,覆草与CK相同为0.38,而四季豆比CK低,为0.37。出油率按大小排列为:毛豆>四季豆>绿豆>覆草>花生,分别比CK提高了17.4%、12.3%、7.7%、7.7%、6.9%。其中毛豆的出油率为45.9%,而花生为41.8%,说明不同栽培模式对油茶鲜出籽率影响不大,影响最大的是单株产量,其次是新梢生长,最后是出油率。因此不同栽培模式即促进了油茶的营养生长,也促进了生殖生长。
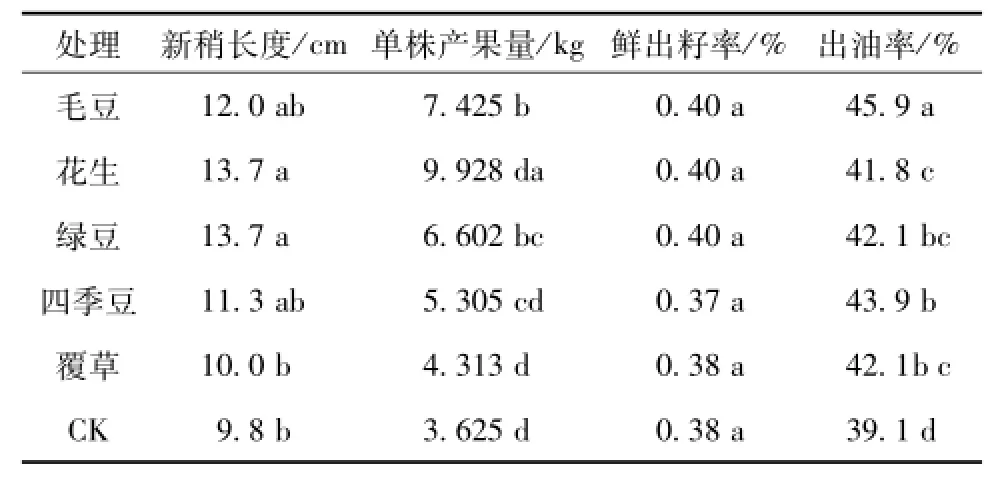
表1 不同栽培模式对油茶新梢生长及产量的影响Tab.1 Shoot growth and yield of Camellia oleifera under different cultivation modes
2.2 不同栽培模式油茶林根际和非根际土壤微生物数量分布特征
由表2可见,不同栽培模式对油茶林土壤微生物的分布产生显著影响。其中,细菌数量最多,占96.72%;其次是放线菌2.99%,真菌最少0.29%。说明细菌在根际微生态中起着生要作用,不同栽培模式油茶林根际土壤微生物数量明显高于非根际土壤且均显著高于CK。
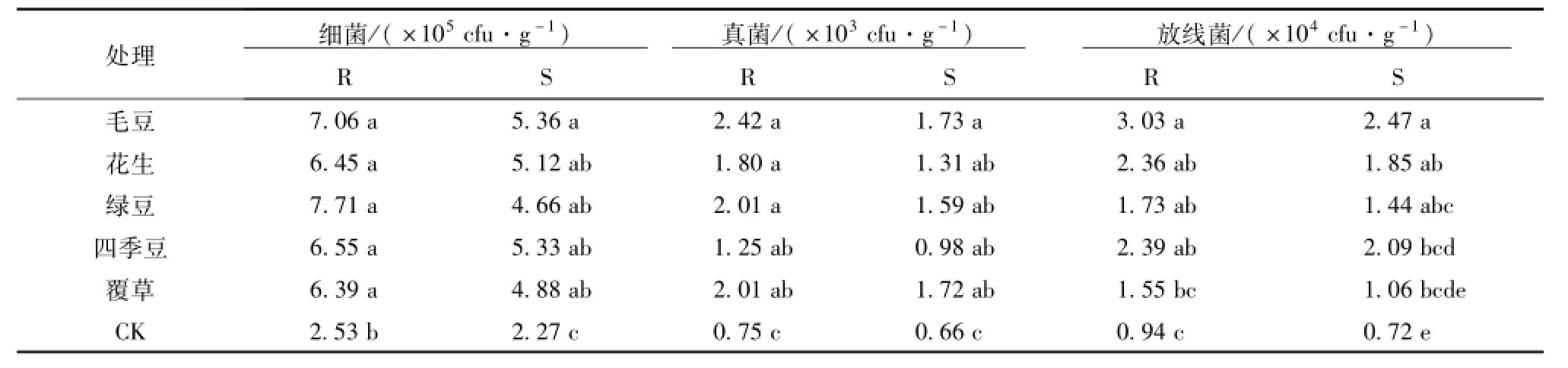
表2 不同栽培模式油茶林土壤微生物数量Tab.2 The amount ofmicrobes in Camellia oleifera soil under different cultivation modes
5种林下栽培模式根际细菌数量变幅为6.39× 105~7.71×105cfu/g,其中绿豆显著高于CK地,覆草最低,按大小排列,绿豆>毛豆>四季豆>花生>覆草,相对CK增加的倍数分别为2.05、1.79、1.59、1.55、1.53倍。非根际细菌数量变幅在4.66×105~5.36×105cfu/g其中毛豆最高,绿豆最低,大小排列如下:毛豆>四季豆>花生>覆草>绿豆,相对CK增加的倍数分别为1.36、1.35、1.26、1.15、1.05倍。可见不同林下栽培模式都促进油茶林土壤细菌的显著提高,其中绿豆最有利于细菌的生长。
根际和非根际的真菌数量均是毛豆最高,四季豆最低。根际的真菌数量变幅在1.25×103~2.42× 105cfu/g,大小排列为:毛豆>覆草>绿豆>花生>四季豆,相对于CK提高的倍数依次为2.22、1.68、1.68、1.40、0.67倍。非根际的真菌数量变幅在0.98×103~1.73×105cfu/g,大小排列如下:毛豆>覆草>绿豆>花生>四季豆,相对于CK提高的倍数依次为1.62、1.60、1.40、0.98、0.48倍。不同林下栽培模式对根际和非根际土壤真菌的影响相同,毛豆最有利于真菌的生长,四季豆最差。
不同栽培模式油茶土壤放线菌数量差异显著。根际土壤放线菌数量最大为毛豆3.03×104cfu/g,覆草最少为1.55×104cfu/g,放线菌数量大小排列如下:毛豆>四季豆>花生>绿豆>覆草;相对于CK处理,各样地增加的倍数为:2.21、1.53、1.50、0.83、0.64倍。非根际土壤放线菌数量最大的为毛豆2.47×104cfu/g,最少的是覆草1.06×104cfu/g,其变化如下:毛豆>四季豆>花生>绿豆>覆草,相对于CK,增加的倍数为:2.41、1.90、1.56、1、0.47倍。可见,毛豆对提高土壤放线菌数量的效果最好,覆草最差。因此油茶林下间种豆科植物,可以显著增加土壤微生物的数量。
2.3 不同栽培模式油茶林根际土壤和非根际土壤酶活性状况
由表3可见,不同栽培模式油茶根际土壤和非根际土壤的蔗糖酶活性存在显著差异。CK样地的蔗糖酶活性最低1.81 mg/g,根际土壤的蔗糖酶活性明显高于非根际。5种栽培模式油茶根际土壤蔗糖酶活性排列如下:绿豆>花生>四季豆>毛豆>覆草,相对于CK分别提高了12.16、8.15、7.71、6.72、3.40倍,绿豆最高,花生次之,覆草最低。非根际土壤蔗糖酶活性排列如下:绿豆>花生>毛豆>四季豆>覆草,相对于CK分别提高了8.28、5.16、4.74、4.63、2.26倍,绿豆最高,花生次之,覆草最低。可见,5种栽培模式油茶非根际和根际土壤蔗糖酶活性最高均为绿豆,分别是16.65和23.82,最低均为覆草,分别是5.85和7.97。
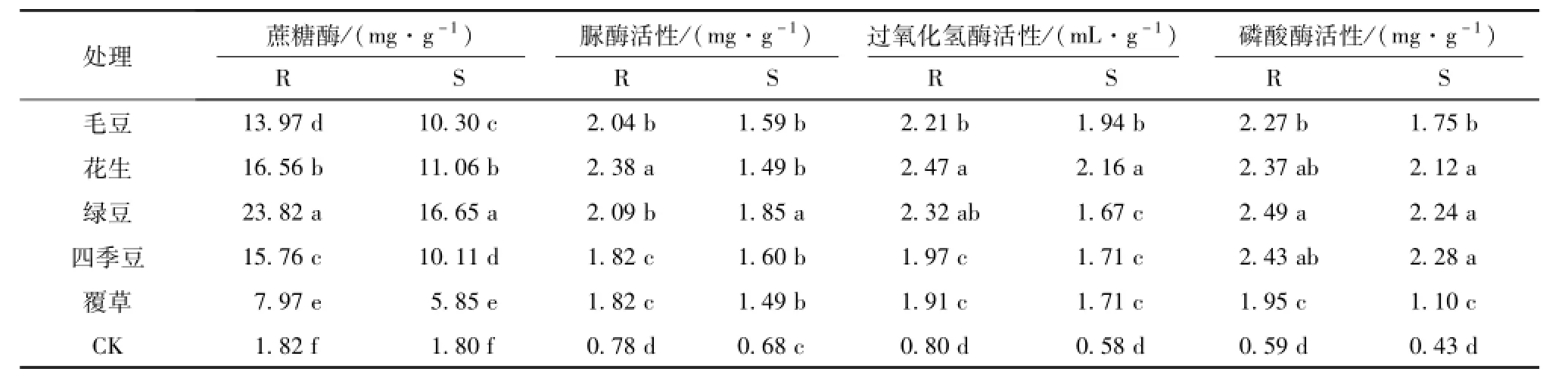
表3 不同生态栽培油茶林土壤酶活性分布特征Tab.3 The distribution characteristics of enzyme activity in Camellia oleifera Abel soil under different cultivationmodes
5种栽培模式油茶根际土壤和非根际土壤脲酶活性存在显著差异。但均比CK样地的脲酶活性高,其中CK非根际土壤脲酶活性只有0.68 mg/g,5种栽培模式油茶非根际脲酶活性变幅在1.49~1.82之间,根际脲酶活性变幅在1.82~2.38之间,根际土壤脲酶活性高于非根际。5种栽培模式非根际土壤脲酶活性排列如下:绿豆>四季豆>毛豆>花生>覆草,相对于CK样地分别提高了1.74、 1.37、1.35、1.20、1.20倍,绿豆最高,毛豆次之,覆草最低;根际土壤脲酶活性排列如下:花生>绿豆>毛豆>覆草、四季豆,相对于CK样地分别提高了2.07、1.69、1.60、1.34、1.34倍,花生最高,绿豆次之,四季豆最低。
不同栽培模式油茶根际土壤和非根际土壤过氧化氢酶活性差异显著。CK样地的过氧化氢酶活性最低仅有0.58 mL/g,非根际土壤过氧化氢酶活性变幅在1.67~2.16间,根际土壤过氧化氢酶活性变幅在1.91~2.47间,根际土壤过氧化氢酶活性明显高于非根际。5种栽培模式非根际土壤过氧化氢酶活性排列如下:花生>毛豆>覆草>四季豆>绿豆,相对于CK样地分别提高了2.72、2.34、1.95、1.94、1.87倍,花生最高,毛豆次之,绿豆最低;根际土壤过氧化氢酶活性排列如下:花生>绿豆>毛豆>四季豆>覆草,相对于CK样地分别提高了2.11、1.91、1.77、1.48、1.40倍,花生最高,绿豆次之,覆草最低。
不同栽培模式油茶根际土壤和非根际土壤酸性磷酸酶活性差异显著。CK样地的酸性磷酸酶活性最低仅有0.43mg/g,非根际土壤酸性磷酸酶活性变幅在1.10~2.28,根际土壤酸性磷酸酶活性变幅在1.95~2.49,根际土壤酸性磷酸酶活性明显高于非根际。5种栽培模式非根际土壤酸性磷酸酶活性排列如下:四季豆>绿豆>花生>毛豆>覆草,相对于CK样地分别提高了4.25、4.16、3.90、3.04、1.54倍,四季豆最高,绿豆次之,覆草最低;根际土壤酸性磷酸酶活性排列如下:绿豆>四季豆>花生>毛豆>覆草,相对于CK样地分别提高了3.22、3.13、3.01、2.84、2.31倍,绿豆最高,四季豆次之,覆草最低。
2.4 不同栽培模式油茶生长指标与土壤微生物数量及酶活性之间的相关性
土壤酶主要来源于土壤微生物的活动及其分泌物,微生物数量在一定程度上表征着土壤酶活性,二者之间存在相关性[22]。对6种油茶栽培模式中的油茶生长指标、土壤微生物数量及酶活性进行相关性分析,结果见表4。
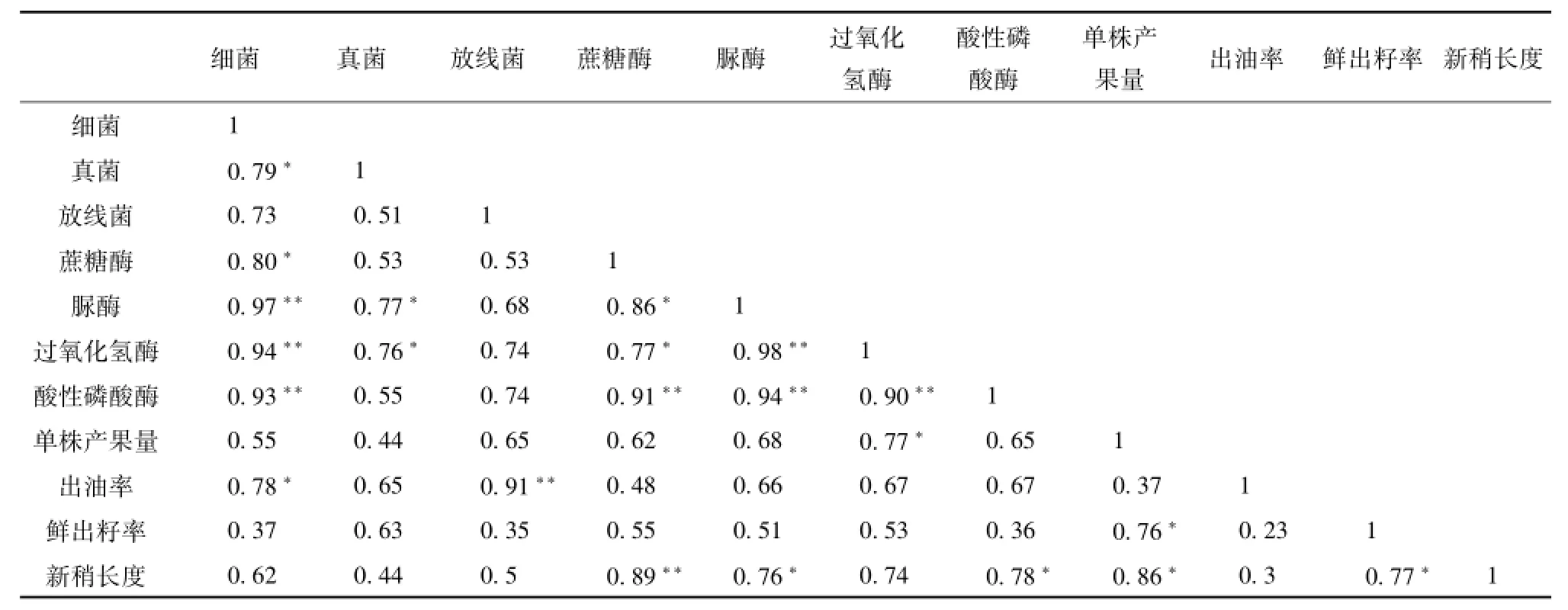
表4 不同栽培模式油茶林新梢生长、产量与土壤微生物、酶活性的相关分析Tab.4 Correlation analysis between growth of new shoot,yield,soilmicrobe number and enzyme activity under different cultivationmodes
由表4可知,细菌与脲酶、过氧化氢酶、酸性磷酸酶呈极显著相关(P<0.01),与真菌、蔗糖酶和出油率呈显著相关(P<0.05),说明细菌影响土壤肥力和油茶的产油量。真菌与细菌、脲酶及过氧化氢酶呈显著相关,说明真菌与土壤抗逆性有关。放线菌只与出油率呈极显著相关;蔗糖酶与细菌、脲酶、过氧化氢酶呈显著相关,与酸性磷酸本科和新梢长度呈极显著相关性;脲酶与细菌、过氧化氢酶和酸性磷酸酶呈极显著相关性,与真菌和新梢生长呈显著相关性;过氧化氢酶与细菌、脲酶和酸性磷酸酶呈极显著相关性,与真菌、蔗糖酶和单株产果量呈显著相关;酸性磷酸酶与细菌、蔗糖酶、脲酶及过氧化氢酶呈极显著相关;单株产果量与过氧化氢酶及鲜出籽率相关。由此说明土壤各种酶以及微生物之间相互作用共同影响油茶生长。可见,在油茶林下栽培不同的植物,根际分泌物促使微生物活动更为活跃,增强了酶促作用强度,从而促进油茶生长和提高其产量。
3 结论与讨论
油茶根系主要分布在0~40 cm土层范围内,该层根系数量占根系总数的70%~80%[23]。绿肥在生长过程中,可以分泌有机酸类,释放土壤中一部分难吸收的化合物,变成可利用的化合物[24]。本试验成熟油茶林下间种豆科植物营养生长较好,但不利于生殖生长。主要是林分郁密度为0.65左右,且树高在2.5m左右,林下植物能够得到相对较好的阳光。成熟油茶林下间种豆科植物促进油茶的营养生长和生殖生长,有可能是花生、绿豆、毛豆以及四季豆根系分布主要分布在0~30 cm,与油茶主要根系充分相互作用,从而影响新梢生长、产果量和出油率。不同栽培模式油茶根际土壤微生物数量明显高于非根际土壤。地面植被类型和生长状况决定了土壤微生物的数量和群落组成,对养分的转化吸收和根系生长有其独特影响[25]。根际细菌数量最大是绿豆,同时非根际中细菌数量最小也是绿豆,说明绿豆的根系对油茶根际细菌有非常明显的刺激作用。真菌数量在根际和非根际中均是毛豆最高,四季豆最低。可能是根系再生活力影响根系分泌物不同,从而导致真菌生长强弱不同[26],因为相比其他栽培模式的豆科植物,大豆根系再生活力最强,四季豆再生能力弱。
细菌与蔗糖酶活性呈显著相关,与脲酶、过氧化氢酶和酸性磷酸酶呈极显著相关;真菌与脲酶和过氧化氢酶呈显著相关;而放线菌与土壤酶无显著关系。说明本试验土壤酶活性的主要来源是细菌,其次是真菌。蔗糖酶的活性可以表示土壤肥力水平[27]。根据蔗糖酶活性大小,得出不同栽培模式对油茶林土壤肥力的影响为绿豆模式最大,覆草最低。可能是不同栽培模式油茶根系土壤中的有机质含量、腐殖质和微生物分泌物的不同,造成了不同栽培模式之间的蔗糖酶活性差异显著。这与相关分析中得出蔗糖酶的活性与脲酶呈显著相关相符合。不同栽培模式根际土壤酶活性大小与非根际的不同,根际土壤脲酶活力可能与套种植物的侧根生长长度和生长量有关,以便套种植物与油茶充分进行物质交流,因此5种套种模式的油茶新稍长度与油茶根际脲酶活性大小排列相似;而非根际土壤脲酶活力可能只与套种植物本身对N的需求量决定,需求量大的则土壤脲酶活力大[28]。过氧化氢酶能氧化土壤呼吸作用产生的过氧化氢,减少对植物体的毒害作用,在一定程度上可以反映土壤呼吸强度[29]。不同栽培模式对土壤的生物氧化作用不同,使得土壤的酸碱度、根系分泌物存在差异,土壤的呼吸强度不同。花生过氧化氢酶活性最高,这可能是花生本身的氧化还原作用较强,带动了油茶根系的生物氧化过程,增强土壤呼吸强度。土壤磷酸酶活性与细菌及新梢长度呈极显著说明酸性磷酸酶具有转化有机或无机磷酸盐的能力,提高磷的有效性[30-31],促进细菌的生长,有效磷及细菌数量的增多进一步提高油茶新梢的生长。绿豆和四季豆对提高油茶根际酸性磷酸酶活性效果显著;覆草的材料主要是五节芒,缺乏足够的酸性磷酸酶酶促底物,磷素循环较慢,磷酸酶活性较低。
因此,不同栽培模式对土壤微生物和酶活性的影响不同,且土壤酶与土壤微生物关系密切,在一定程度上综合影响油茶林的生长和产量,但以绿豆和毛豆最优。不同品种对油茶的生长和产量影响也很大[32],不同栽培模式与不同品种的交互效应还有待进一步研究。
[1] 史洁,尹佟明,施季森.我国油茶产业的发展历程及存在问题述评[J].西南林业大学学报,2011,31(6):82-87.
[2] 李纪元,肖青,李辛雷,等.不同套种模式油茶幼林水土流失及养分损耗[J].林业科学,2008,44(4):167-172.
[3] 周凯敏,廖美霞.不同抚育管理措施对油茶丰产栽培的影响[J].安徽农学通报,2013,19(4):51-52.
[4] 郑灏,杨志坚,冯金玲,等.不同林下套种模式对油茶幼林根区土壤化学特性及微生物的影响[J].福建农林大学学报(自然科学版),2015,44(2):147-153.
[5] 李云,黄永芳,叶小萍,等.油茶幼林间种大豆对土壤养分含量的影响[J].经济林研究,2013,31(2):54-59.
[6] 刘丁林,张杨珠,聂军,等.不同绿肥作物与油茶幼林套种对丘岗红壤pH值变化和有效养分含量的影响[J].湖南农业科学,2013(9):43-45.
[7] 柏明娥,洪利兴,沈建军.油茶林种植决明对土壤生态和油茶生长的影响[J].山东林业科技,2014,44(1):28-30.
[8] 王东雪,曾雯珺,杨菊华,等.油茶新造林套种花生试验初报[J].西北林学院学报,2012,27(4):157-159.
[9] 王玉娟,陈永忠,王瑞,等.稻草覆盖对油茶幼林土壤理化性质及油茶生长的影响[J].浙江农林大学学报,2012,29(6):811-816.
[10] 吴兆荣.油茶与茶的不同栽培模式对土壤化学性质和酶活性的影响[J].安徽林业科技,2013,39(4):43-45.
[11] 王玉娟,陈永忠,王瑞,等.覆草间种对油茶林土壤养分及生长量影响的主成分分析[J].中南林业科技大学学报,2010,30(6):43-49.
[12] 吴晓琳.仙居县老油茶林套种生姜复合栽培试点成功[J].浙江林业,2010(12):35.
[13] DE-Bashan L E,Hernandez JP,Bashan Y.The potential contribution of plant growth-promoting bacteria to reduce environmental degradation:A comprehensive evaluation[J].Applied Soil Ecology,2012,61:171-189.
[14] Nie M,Zhang X D,Wang JQ,et al.Rhizosphere effects on soil bacterial abundance and diversity in the Yellow River Deltaic ecosystem as influenced by petroleum contamination and soil salinization[J].Soil Biology and Bio-chemistry,2009,41(12):2535-2542.
[15] 王齐,师春娟,周德全,等.中水灌溉对城市绿地土壤微生物数量的促进效应[J].西南林业大学学报,2012,32(3):12-16.
[16] Sharma S K,Ramesh A,Sharma,M P,et al.Microbial Community Structure and Diversity as Indicators for E-valuating Soil Quality[M].Springer Netherlands,2011:317-358.
[17] Fekete I,Varga C,Kotrocao Z,et al.The relation between various detritus inputs and soil enzyme activities in a Central European deciduous forest[J].Geoderma,2011,167:15-21.
[18] 雍太文,杨文钰,向达兵,等.不同种植模式对作物根系生长、产量及根际土壤微生物数量的影响[J].应用生态学报,2012,23(1):125-132.
[19] 赵兰坡.土壤酶活性测定方法的探讨[M].北京:中国学术期刊电子出版社,2012:138.
[20] 关松荫.士壤酶及其研究法[M].北京:农业出版社,1986.
[21] 方敏,丁小霞,李培武,等.索氏抽提测定含油量的方法改良及其应用[J].中国油料作物学报,2012,34(2):210-214.
[22] 叶存旺,翟巧绒,郭梓娟,等.沙棘-侧柏混交林土壤养分、微生物与酶活性的研究[J].西北林学院学报,2007,22(5):1-6.
[23] 袁军,谭晓风,袁德义,等.油茶根系分布规律调查研究[J].浙江林业科技,2009,29(4):30-32.
[24] 徐祥玉,王海明,袁家富,等.不同绿肥对土壤肥力质量及其烟叶产质量的影响[J].中国农学通报,2009,25(13):58-61.
[25] Mariotte P,Vandenbenberghe C,Meugnier C,et al. Subordinate plant species impact on soilmicrobial communities and ecosystem functioning in grasslands:Findings from a removal experiment[J].Perspectives in Plant Ecology,Evolution and Systematics,2013,15(2):77-85.
[26] 田胜尼,刘登义,王峥峰,等.铜尾矿对5种豆科植物根系生长的影响[J].生态环境,2005,14(2):199-203.
[27] Hofman J,Bezchlebova J,Dusek L,et al.Novel approach tomonitoring of the soil biological quality[J].Environment International,2003,28(8):771-778.
[28] Akmal M,Aitaf M S,Hayat R,et al.Temporal changes in soil urease,alkaline phosphatase and dehydrogenaseactivity in rainfed wheat field of Pakistan[J].JAnim Plant Sci,2012,22(2):457-462.
[29] 葛晓改,肖文发,曾立雄,等.三峡库区不同林龄马尾松土壤养分与酶活性的关系[J].应用生态学报,2012,23(2):445-451.
[30] 任万军,黄云,吴锦秀,等.免耕与秸秆高留茬还田对抛秧稻田土壤酶活性的影响[J].应用生态学报,2011,22(11):2913-2918.
[31] Pant H K,Warman P R.Enzymatic hydrolysis of soil organic phosphorus by immobilized phosphatases[J].Biology and Fertility of Soils,2000,30(4):306-311.
[32] 陈隆升,陈永忠,王瑞,等.不同油茶新种质产量差异研究[J].西南林业大学学报,2015,35(4):95-98.
(责任编辑 张 坤)
Effects of Five Kinds of Cultivation on the Soil Microorganisms and Enzyme Activities of Camellia oleifera Forestry
Feng Jinling,Zheng Xinjuan,Yang Zhijian,Chen Shipin,Chen Hui
(College of Forestry,Fujian Agriculture and Forestry University,Fuzhou Fujian 350002,China)
In this paper,five cultivationmodes of experimentwhich are interplant of Soybean,peanut,mung bean,kidney bean and the covering grass,were explored to discuss the effects of different cultivation on Camellia oleifera,soilmicroorganisms and enzyme activities.The results indicated that new-tip length,the yield of single tree and oil yield of the five cultivationmodes had significantly differences,except for fresh seeds yield.The quantities ofmicrobes and the activities of enzymes in the rhizosphere soilwere higher than in the non-rhizosphere soil. Based on the above analysis,interplant of soy beans and mung beans were the most beneficialmodes to C.Oleifera.The bacteria was significant correlation with various kinds of soil enzyme and oil yield,which illustrated that the quantity of bacteria could be used as biological indicator to detect soil fertility,and providing the basis to judge C.Oleifera oil production.
Camellia oleifera;rhizosphere;soil;microorganisms;enzyme activity;interplant

S714.8
A
2095-1914(2016)02-0010-07
10.11929/j.issn.2095-1914.2016.02.002
2015-07-17
高等学校博士学科点专项科研基金(20123515110010)项目资助;福建省科技平台建设项目(2010N2001)资助;福建省教育厅科研基金(JA15191)资助;横向课题—南方红壤丘陵区油茶林水土保持研究与示范(KH1500310)资助。
第1作者:冯金玲(1978—),女,博士,讲师。研究方向:经济林栽培。Email:fjl9703@163.com。
陈辉(1957—),男,教授。研究方向:经济林栽培。Email:zjchchenh@163.com。
