钩苞大丁草的叶片形态多样性研究
2016-03-31欧成川王谈笑徐晓丹
欧成川,郑 伟,王谈笑,陈 菁,王 炜,徐晓丹
(昆明理工大学现代农业工程学院,云南昆明 650500)
钩苞大丁草的叶片形态多样性研究
欧成川,郑 伟,王谈笑,陈 菁,王 炜,徐晓丹*
(昆明理工大学现代农业工程学院,云南昆明 650500)
摘 要:钩苞大丁草(Gerbera delavayi Franch)的叶片形态差异较大,而这与其叶背优异纤维种质的筛选密切相关。通过对叶片长、叶柄长、叶片宽、叶片面积、叶片鲜重和叶形指数的测定,对中国西南地区8个钩苞大丁草居群的叶片形态多样性进行了分析研究。结果表明,8个居群中,东川居群的叶片性状多样性较为丰富,Shannon-Wiener指数为2.08;而新平居群的各叶片性状更为稳定。钩苞大丁草叶片的宽度比长度的多样性更丰富。方差分析及多重比较结果显示,6个指标在居群间存在显著差异,F值分别为20.87,18.13,7.89,27.07,29.04和2.86;新平和德昌居群的叶片长、叶片宽、叶片面积以及叶片鲜重显著高于其他居群。通过UPGMA分析,8个居群的钩苞大丁草可分为2类,其中新平、德昌、洱源、开远为一类,丽江、武定、东川、石林为一类。
关键词:钩苞大丁草;叶片;种质资源;遗传多样性;聚类分析
DOI 10.16178/j.issn.0528-9017.20160217
钩苞大丁草(Gerbera delavayi Franch.)为菊科大丁草属的多年生草本植物,分布于云南省、四川省南部及越南北部[1],因其叶片的背部有白色纤维,古人将其撕下晒干作火镰打火用的火绒,故命名为火草,也称之为火石花[2-3]。钩苞大丁草的应用价值主要表现在:(1)纺织。钩苞大丁草叶背的毡毛纤维是我国西南少数民族人民编织火草服饰的原料。火草衣具有冬暖夏凉、越洗越白的特点[4],是我国西南地区彝族、傈僳族、白族、纳西族、壮族等民族爱与智慧的象征,常作男女的定情之物。火草纺织技艺已有500多年历史,是世界上罕见的将植物叶背毡毛纤维直接用于手工纺织的技艺。据文献记载,钩苞大丁草的叶背毡毛纤维是迄今用于纺织的最短的天然植物纤维(长度3~6 mm),其利用丰富了纺织工业的天然植物纤维种类。钩苞大丁草的叶背毡毛纤维的收集采用手工剥离并同时捻线的方式,与苎麻等天然植物纤维相比,采集过程无须经过任何化学处理,因而更能适应人类对环保和健康的要求。2009年,云南省马龙县和丘北县的火草纺织技艺分别被列为省级非物质文化遗产; 2007和2011年,四川省德昌县和米易县的火草纺织技艺分别被列为非物质文化遗产。(2)药用。钩苞大丁草的根系含有香豆素等杀菌成分,可用于治疗痢疾、胃痛、消化不良等[5]。(3)观赏。钩苞大丁草的花可作为鲜切花;钩苞大丁草与著名鲜切花非洲菊同属,可作为非洲菊远缘杂交育种的亲本。由此可见,钩苞大丁草的叶背毡毛纤维特色鲜明,具有极大的开发潜力。
然而,钩苞大丁草仍处于野生状态,尚未实现人工种植。随着火草衣价格的攀升(市场价每件为4 000~5 000元),钩苞大丁草鲜叶的价格也迅速上升(2014年高达500元·kg-1)。目前,过度的鲜叶采摘已严重影响到钩苞大丁草的正常生长和种群繁育。特别是根系被用作中药材,导致部分地区的钩苞大丁草还被大量的采挖。加上外来物种紫茎泽兰的肆意蔓延,钩苞大丁草的生存空间也越来越小。总之,钩苞大丁草的野生资源正受到多方面的威胁,急剧减少,钩苞大丁草的保护性开发势在必行。
遗传多样性研究是保护性开发的重要依据。本文选取云南和四川的8个钩苞大丁草居群64份单株为试验材料,通过对其叶片的各项形态指标进行统计分析和聚类分析,为钩苞大丁草野生种质资源的保护和优异纤维种质的筛选提供参考数据,促进钩苞大丁草的保护性开发。
1 材料与方法
1.1材料来源
试验材料于2014年8—12月采自8个不同野生钩苞大丁草居群。采样前,查阅《云南植物志》,了解钩苞大丁草的分布区,在分布区内均匀选取云南省的昆明市东川区、石林彝族自治县、洱源县、丽江市、开远市、新平彝族傣族自治县、武定县,以及四川省的德昌县8个采样点(图1)。采样时,参考金燕等[6]的方法,每个居群随机取样30~50份同质的单株(相互间隔>5 m)。最后,将所有样本植株在昆明理工大学的试验基地集中种植,统一管理。

图1 钩苞大丁草不同居群的采样点
叶片样本采集在2015年7月进行,每个居群随机选择8份无病虫害的植株,分别采集1片新成熟的叶片用于叶片形态多样性分析。
1.2方法
叶长、叶宽、叶柄长均用游标卡尺测量,精确到0.01 mm。叶宽为叶片最宽处的长度[7]。叶片鲜重采用电子天平测量,精确到0.01 g。
叶面积的测定方法。(1)将钩苞大丁草叶片固定在标准方格纸上,用数码相机对其正面拍照; (2)将照片插入到Auto CAD 2010中,运行“工具-查询-面积”的功能,描出方格纸单元格和叶片在图片中的面积; (3)方格纸单元格的实际面积已知,根据比例算出叶片实际面积;精确到0.01 mm2。
1.3数据处理
运用Auto CAD 2010计算钩苞大丁草的叶片面积;运用Bio-dap软件计算Shannon-Wiener指数;运用IBM SPSS Statistics 19.0软件进行叶片长、叶片宽、叶柄长、叶片面积、叶片鲜重、叶形指数的统计分析和聚类分析。
2 结果与分析
2.1钩苞大丁草不同居群叶片形态的多样性指数分析
由表1可知,8个钩苞大丁草居群的平均Shannon-Wiener指数为4.10。叶宽和叶形的Shannon-Wiener指数为4.14,而叶长的Shannon-Wiener指数为4.04。说明与长度相比,钩苞大丁草叶片在宽度上更富有变化,而长度上的多样性更多地体现在叶柄长度上(4.13)。各个居群中,Shannon-Wiener指数最大的是东川,为2.08;最小的是新平,为2.05。说明东川居群的钩苞大丁草叶片性状多样性较为丰富,而新平的各叶片性状更为稳定。

表1 钩苞大丁草不同居群的叶片形态多样性指数
2.2钩苞大丁草不同居群叶片形态指标的方差分析与多重比较
由图2可知,通过对8个居群的钩苞大丁草叶片进行方差分析表明,不同居群间的6个形态指标均达到显著差异,其中叶长、叶宽、叶柄长、叶面积、鲜重的F值分别为20.87,18.13,7.89,27.07和29.04,差异均达到极显著水平。叶鲜重差异最大,其次是叶面积和叶长,叶形指数差异最小。这说明不同居群钩苞大丁草种质资源叶片之间存在广泛差异。

图2 钩苞大丁草不同居群的叶片(背面)
由表2可知,通过对8个居群的6个性状做多重比较表明,(1)在8个居群中,新平和德昌的叶片最长,分别为122.70和122.62 mm,且与其他居群存在显著差异;叶长最短的是东川居群,为73.60 mm。(2)叶片最宽的居群是新平和德昌,分别为50.99和50.35 mm,且与其他居群存在显著差异;叶片最窄的是丽江居群,为28.19 mm。(3)叶柄最长的是开远居群,为58.86 mm;最短的是东川居群,为28.40 mm。(4)叶面积最大的是德昌居群,为5 722.03 mm2,其次是新平居群,为5 667.40 mm2,且与其他居群存在显著差异;叶面积最小的是武定居群,为1 788.99 mm2。(5)叶鲜重最大的是新平居群,为2.21 g,其次是德昌居群,为2.03 g,且与其他居群差异显著;鲜重最小的是丽江居群,为0.69 g。(6)叶形指数最大的是石林居群,为2.94;最小的东川居群,为2.29。
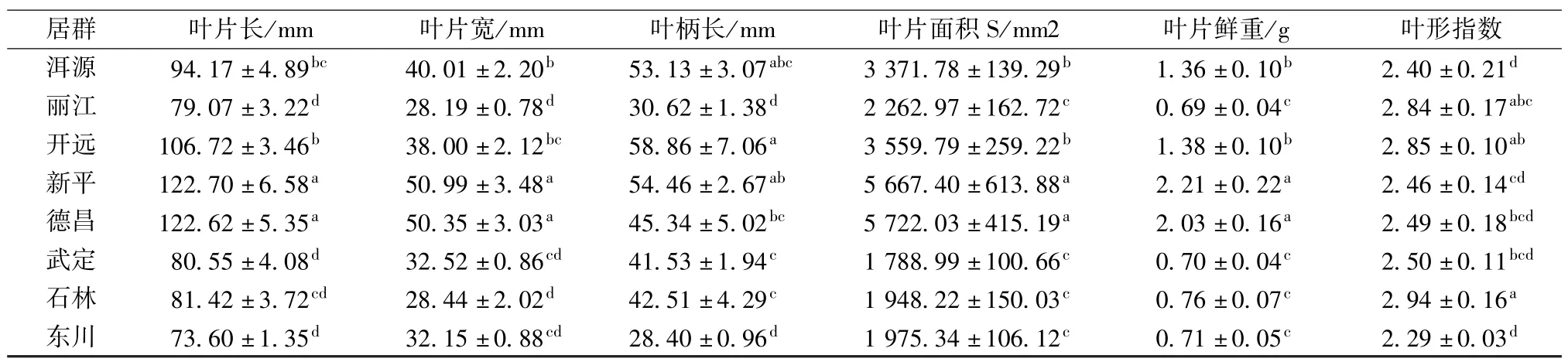
表2 钩苞大丁草不同居群的叶片形态比较
基于以上结果,在叶长、叶宽、叶面积以及叶片鲜重等指标的均值上,新平和德昌钩苞大丁草居群普遍高于其他居群,且与其他居群存在显著性差异,而这2个居群之间不存在显著性差异。究其原因,四川德昌和云南新平在地域上并不相近,但相对于其他几个居群,这2个地区的共同点是海拔相对较低,温度较高,可能是气候因素导致了差异性,具体原因还有待进一步研究。
2.3聚类分析
利用IBM SPSS Statistics 19.0软件对钩苞大丁草进行聚类分析(图3)。在欧氏距离为15时,可将8个居群的钩苞大丁草分为2类。其中,新平彝族傣族自治县、四川省的德昌县、洱源县、开远市的4个居群为1类;昆明东川区、石林彝族自治县、丽江市、武定县的4个居群为另1类。

图3 钩苞大丁草8个居群的聚类分析
3 小结与讨论
在野外不同环境中表现出的叶片变异性状,当集中种植时可能会消失[8]。因此,要从表型水平准确了解物种的遗传变异状况,须将植物在集中地栽培,尽可能减少环境条件的差异带来的影响[9]。本研究中,不同居群的钩苞大丁草在同一试验地集中种植时,叶片形态不仅在居群间存在显著差异,而且在居群内也显示出较高的变异。这说明钩苞大丁草在叶片形态上具有丰富的遗传多样性。钩苞大丁草是严格的异花传粉植物,实生繁殖后代变异大,可能是居群内叶片形态变异较大的主要原因。而西南地区海拔起伏较大、气候类型多样可能是导致居群间叶片形态变异较大的主要因素。
聚类分析结果显示,在欧氏距离为15时,可将8个居群的钩苞大丁草分为2类。新平彝族傣族自治县、四川省的德昌县、洱源县、开远市的4个居群为1类;昆明东川区、石林彝族自治县、丽江市、武定县的4个居群为1类。在第1类群中,德昌居群与其他3个居群的地理距离较远,因此与实际的地理格局并不相同。但这4个居群在海拔和温度上都具有海拔相对较低、温度较高的特点,这可能是它们聚为一支的主要原因。
一般来讲,Shannon-Wiener指数越高,生态系统的物种越丰富,生态系统越稳定,其维持、繁衍和保护物种多样性的能力越强,因此其物种保育价值也越高[10]。本研究通过采集8个居群的200多份钩苞大丁草样本,在昆明构建了资源圃,并从叶片形态上对其遗传多样性展开了初步研究,可为钩苞大丁草的种质资源保护提供科学依据。
钩苞大丁草在纺织原料中的价值和在民族文化中的特殊含义都是不可多得的瑰宝。今后,可结合叶片背面的纤维形态多样性研究,筛选具有优异纤维的种质资源,进而提供人工种植的优质种源,促进钩苞大丁草的人工种植,继承和发扬火草纺织技艺这项非物质文化遗产。
参考文献:
[1]程用谦.中国植物志79卷[M].北京:科学出版社,1996:73.
[2]李晓岑,李云.中国西南少数民族的火草布纺织[J].云南社会科学,2010 (2):64-67.
[3]吴征镒,彭华.国产广义大丁草属的订正及地理分布[J].云南植物研究,2002 (2):137-146.
[4]欧丽.彝族“罗噜颇”的火草麻布纺织[J].毕节学院学报,2009 (11):42-47.
[5]LIU S Z,FENG J Q,WU J,et al.A new monoterpenecoumarin and a new monoterpene-chromone from Gerbera delavayi [J].Helvetica Chimica Acta,2010,93:2026-2029.
[6]金燕,卢宝荣.遗传多样性的取样策略[J].生物多样性,2003,11 (2):155-162.
[7]扈慧灵,刘遵春,苗卫东,等.柿树种质资源叶片形态多样性研究与聚类分析[J].河南农业科学,2012,41 (6):137-139.
[8]HOVENDEN M J,SCHOOR J K V.Nature versus nurture in the leaf morphology of Southern beech.Nothofagus cunninghamii (Nothofagaceae)[J].New Phytologist,2004,161:585-594.
[9]HOVENDEN M J,VANDER SCHOOR J K.The response of leaf morphology to irradiance depends on altitude of origin in Nothofagus cunninghamii[J].New Phytologist,2006,169:291-297.
[10]王兵,郑秋红,郭浩.基于Shannon-Wiener指数的中国森林物种多样性保育价值评估方法[J].林业科学研究,2008,21 (2):268-274.
(责任编辑:张瑞麟)
通信作者:徐晓丹,E-mail:125128370@qq.com。
作者简介:欧成川(1990—),男,安徽六安人,在读硕士,研究方向为植物种质资源,E-mail:977693612@qq.com。
基金项目:云南省应用基础研究计划项目(31560086)
收稿日期:2015-12-09
中图分类号:S567
文献标志码:A
文章编号:0528-9017(2016)02-0194-04
文献著录格式:欧成川,郑伟,王谈笑,等.钩苞大丁草的叶片形态多样性研究[J].浙江农业科学,2016,57 (2):194-197.
