长江口邻近海域夏季大型底栖生物群落特征*
2016-03-15李新正王洪法张宝琳
杨 梅 李新正① 王洪法 张宝琳
(1. 中国科学院海洋研究所 海洋生物分类与系统演化实验室, 青岛 266071; 2. 青岛海洋科学与技术国家实验室 海洋生物学与生物技术功能实验室, 青岛 266200)
长江口邻近海域夏季大型底栖生物群落特征*
杨 梅1,2李新正1,2①王洪法1张宝琳1
(1. 中国科学院海洋研究所 海洋生物分类与系统演化实验室, 青岛 266071; 2. 青岛海洋科学与技术国家实验室 海洋生物学与生物技术功能实验室, 青岛 266200)
本文根据 2014年 6月长江口邻近海域大型底栖生物的调查资料, 采用优势度指数、物种多样性指数、丰富度指数、均匀度指数、Bray-Curtis相似性聚类和非参数多维标度排序(nMDS)标序方法, 分析了长江口邻近海域夏季大型底栖生物的群落特征。本次调查共采集大型底栖生物135种, 其中多毛类动物66种、甲壳动物33种、棘皮动物14种、软体动物13种、其他类群动物9种。调查海域的大型底栖生物平均丰度为122.2ind./m2, 平均生物量为7.8g/m2。丰度、生物量和多样性指数在不同海区间的空间差异均不显著。大型底栖生物在 20%的相似性水平上可以划分为 8个群组,各站位间相似性水平较低。ABC曲线表明, 伴随海岸带开发及人类活动的持续影响,长江口邻近海域底栖生境的稳定性受到轻微扰动, 建议继续开展长期连续监测, 为研究长江口海域环境变化和大型底栖生物群落演替提供基础资料。
长江口; 大型底栖生物; 多样性; 群落结构
长江口是我国最大的河口, 受长江冲淡水、苏北沿岸流、黄海海流和台湾暖流等影响,其水文结构复杂, 环境因子多变, 营养盐含量高, 饵料资源丰富, 为众多鱼类及无脊椎动物的产卵、育幼及索饵洄游等提供了极佳的场所(李建生等, 2006)。大型底栖生物是海洋生态系统中的重要组成部分, 通过摄食和次级生产等方式参与生态系统的物质循环和能量流动(李新正等, 2012)。此外, 大型底栖生物还兼有种类丰富、分布广泛、回避污染能力差等特点, 对环境变化较为敏感, 因此研究其多样性特征和群落结构有助于揭示海洋生态环境的动态变化(蔡文倩等, 2013)。有关长江口底栖生物的生态调查始于 20世纪 50年代, 即1958~1960年开展的第一次全国海洋普查就已涉及该海区。伴随研究方法的改进和一系列研究项目的实施, 相关学者对长江口及邻近海域底栖生物的丰度和生物量动态变化(吴耀泉, 2007; 刘勇等, 2008; 王延明等, 2009)、多样性指数变动趋势(王金辉等, 2004)、群落特征(吴耀泉和李新正, 2003; 李宝泉等, 2007;孙亚伟等, 2007; 杨金龙等, 2014)及与环境因子的相关性(Shou et al., 2013; 贾海波等, 2014)等进行了诸多分析, 积累了大量基础资料。近年来, 随着沿岸经济的快速发展、边滩湿地的围垦及大型海岸工程的建设、人类向海洋排污量的增加, 水体营养盐比例失调, 入海泥沙沉积率改变, 长江口生态系统的稳定性遭到破坏, 底栖生物的多样性和空间分布均发生较大变化, 长江口生态面临生境退化、群落结构单一脆弱、水产资源衰退等一系列生态困境(叶属峰等, 2004; 刘录三等, 2008, 2012)。本文根据2014年6月长江口邻近海域大型底栖生物的调查资料, 对底栖生物的多样性特征和群落结构等进行分析, 并与历史资料进行对比,旨在为长江口生态环境的保护和底栖生物资源的可持续利用提供科学依据和资料积累。
1 材料与方法
1.1 调查站位和样品采集
于2012年6月在长江口邻近海域122°E~ 125°E、28.65°N~33°N范围内设置30个站位(图1) 开展大型底栖生物定点调查。每站使用0.1m2箱式采泥器成功取样两次合并为当站样品, 泥样经孔径 0.5mm 网目的套筛冲洗, 保留底栖生物样品, 固定保存于75% 的酒精中,带回实验室进行种类鉴定、个体计数和称重。样品的采集、保存、计数和称重均按《海洋调查规范》(2008年)进行。
1.2 数据处理
1.2.1 优势度指数
优势度指数的计算采用下列公式:
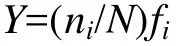
式中, Y为底栖生物优势度指数; ni为第i种的总个体数; N为在所有站位采集的底栖生物总个体数; fi为该物种在各调查站位中出现的频率。优势度指数Y≥0.02时, 即认为该物种为优势种(徐兆礼和陈亚瞿, 1989)。为便于比较, 将Y值小于但接近0.02的常见种也在表中列出。

图1 长江口邻近海域大型底栖生物调查站位示意图Fig.1 Sampling stations of macrobenthos in adjacent sea of Yangtze River estuary
1.2.2 群落多样性指数
群落多样性是衡量群落稳定性的重要尺度, 一般采用Shannon-Wiener指数、物种丰富度指数和物种均匀度指数来综合分析物种的多样性及影响因素。
物种多样性指数(H'), 采用 Shannon-Wiener计算公式(Shannon and Wiener, 1949):

物种丰富度指数(D), 反映一个群落中物种数目的多寡, 采用Margalef (1968)计算公式:
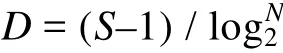
物种均匀度指数(J), 表现群落中全部物种个体数目的分配情况, 即表现种属组成的均匀程度, 采用Pielou (1966)计算公式:

式中, N为采泥样品中所有种类的总个体数; S为采泥样品中的种类总数; Pi为第i种的个体数与样品中总个数的比值(ni/N)。
采用SPSS 16.0软件中的单因素方差分析(one-way ANOVA)来判断各生物指标(包括丰度和生物量)在长江口北部海区(I1-I6, K1-K6)、长江口海区(M1-M5, 3100-2, 3100-3, 3100-6)和舟山海区(DH3000-2, DH3000-4, DH3-2, DH3-3, DH4-1, DH4-2, DH4-3, DH4-5, DH5-1, DH5-2, DH5-3)的空间差异。
1.2.3 群落结构分析
使用多元统计软件Primer 6.0(Clarke, 1993)分析长江口邻近海域大型底栖生物的群落结构。分析之前去除只在一个站位出现的物种,以减少稀有种效应。同时, 为避免个别数值过高或过低引起的偶然性误差, 将大型底栖生物丰度数据进行平方根转化(周红和张志南, 2003), 然后采用 Bray-Curtis相似性等级聚类(CLUSTER)、非度量多维标度(nMDS)及相似性百分比(similarity percentage program, SIMPER)方法, 分析比较群组间的相似性和主要贡献种。采用丰度/生物量比较曲线(abundance biomass comparison, ABC)(田胜艳等, 2006)进行群落稳定性分析, 以研究长江口邻近海域大型底栖生物群落受污染或其他因素扰动的程度。
2 结果
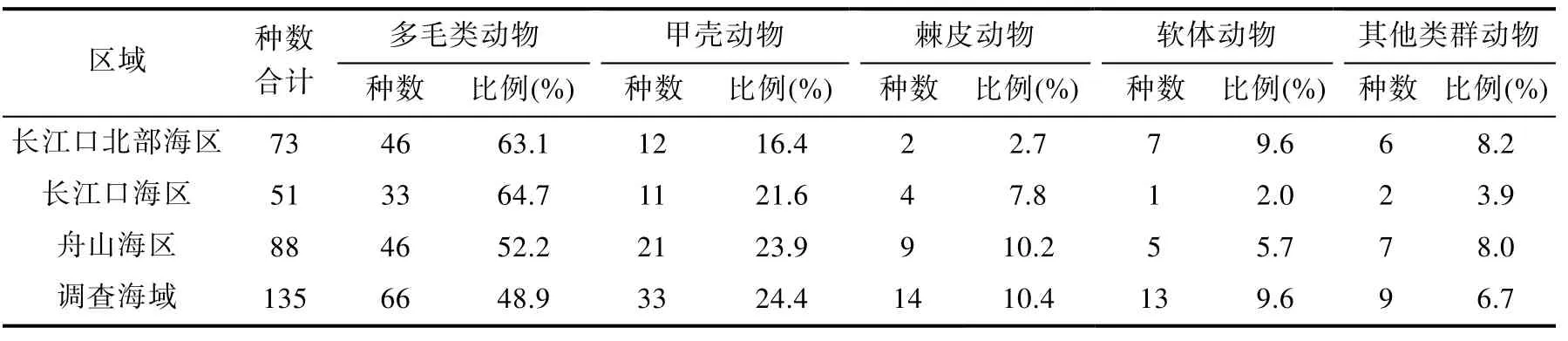
表1 长江口邻近海域大型底栖生物种类组成及所占比例Tab.1 Species composition of macrobenthos in adjacent sea of Yangtze River estuary
2.1 种类组成和优势种
本次调查共采集大型底栖生物135种, 其中环节动物多毛类最多, 有 66种(占 48.9%);甲壳动物次之, 33种(24.4%); 棘皮动物14种(10.4%); 软体动物 13种(9.6%); 其他类群动物(腔肠动物、纽虫、星虫和鱼类) 9种(6.7%)。各海区底栖生物种类组成以舟山海区最多,为88种; 其次为长江口北部海区, 73种; 长江口海区最少, 为51种(表1)。采到物种数最多的站位是DH4-3, 共26种, 最少的是M3, 仅有1种。

表2 长江口邻近海域大型底栖生物的优势度(Y)Tab.2 Dominance (Y) of macrobenthos in adjacent sea of Yangtze River estuary
表2列出了本航次优势度(Y)前10位的物种, 全部为多毛类动物, 优势种包括背蚓虫(Notomastus latericeus)、蜈蚣欧努菲虫(Onuphis geophiliformis) 和长叶索沙蚕(Lumbrineris longifolia), 其中背蚓虫的优势度最高(Y=0.026)。
2.2 丰度与生物量
2014年, 长江口邻近海域夏季大型底栖生物的平均丰度为122.2ind./m2(表3), 不同类群的平均丰度由高到低依次为多毛类动物(占总丰度的比例是 75.2%)、甲壳动物(11.5%)、棘皮动物(7.1%)、其他类群动物(3.7%)、软体动物(2.5%)。各海区平均丰度的大小顺序为长江口北部海区(165.9ind./m2) >舟山海区(115.6ind./m2) >长江口海区(85ind./m2), 单因素方差分析表明, 丰度在不同海区间的差异不显著(F2,27=0.191, P=0.665)。由图2可以看出, 调查站位的丰度呈不连续的斑块状分布, 其中丰度最高的站位为K3(285ind./m2), 最低的为M3(5ind./m2)。

表3 长江口邻近海域大型底栖生物主要类群的丰度和生物量Tab.3 The abundance and biomass of main macrobenthic groups in adjacent sea of Yangtze River estuary

图2 长江口邻近海域大型底栖生物丰度的空间分布Fig.2 Abundance (ind./m2) distribution of macrobenthos in adjacent sea of Yangtze River estuary
调查海域大型底栖生物的平均生物量为7.8g/m2(表 3), 不同类群的贡献率由高到低为多毛类动物41.0%、其他类群动物25.6%、棘皮动物 16.7%、甲壳动物 11.5%、软体动物5.2%。就不同海区而言, 平均生物量最高的是舟山海区(9.4g/m2), 其次是长江口北部海区(8.5g/m2), 长江口海区最低(5.4g/m2), 单因素方差分析表明, 平均生物量的空间差异不显著(F2,27=0.685, P=0.513)。30个调查站位的平均生物量存在较大差异, 最高的站位为 DH5-1 (22.3g/m2), 最低的为K4(0.03g/m2) (图3)。

图3 长江口邻近海域大型底栖生物生物量的空间分布Fig.3 Biomass (g/m2) distribution of macrobenthos in adjacent sea of Yangtze River estuary
2.3 群落多样性指数
长江口邻近海域大型底栖生物的群落多样性指数见表4。从空间上看, 舟山海区的物种多样性指数(H')、物种丰富度指数(D)和物种均匀度指数(J)最高, 长江口北部海区次之,长江口海区的多样性指数最低; 单因素方差分析表明, 上述多样性指数在不同海区间的差异不显著 (H': F2,27=1.970, P=0.159; D: F2,27= 1.427, P=0.257; J: F2,27=0.754, P=0.480)。除M3站位外 (只采集到1个物种), H'值最高的为 K5站位(4.34), 其次为 DH4-3(4.25), 最低为K4(1.00); D值最高的为DH4-3(6.30), 其次为 K5(6.09), 最低为 K4(1.44); J值最高的为K4和M4(均为1.00), 其次为DH5-3(0.98), 最低为K1(0.68)。
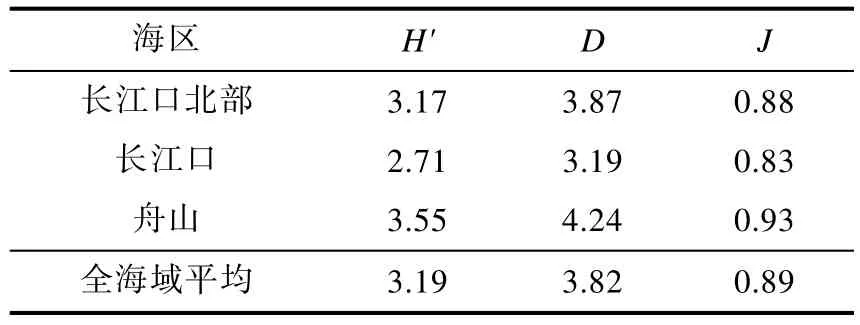
表4 长江口邻近海域大型底栖生物群落多样性指数Tab.4 Shannon-Wiener index (H'), Margalef richness index (D) and Pielou evenness index (J) of macrobenthos in adjacent sea of Yangtze River estuary
2.4 群落结构分析
对长江口邻近海域的大型底栖生物群落进行 CLUSTER聚类和非参数多维标度排序(nMDS)分析 (图4), 结果表明, 在20%的Bray-Curtis相似性水平上, 调查海域的大型底栖生物可以划分为8个群组(图4), 没有相似性水平超过 60%的站位, 站位间的相似性水平较低。
群组1包括半数以上的调查站位(I3、I4、I5、K1、K3、K5、K6、M5、3100-2、3100-3、3100-6、DH3000-2、DH3-2、DH3-3、DH4-1、DH4-3、DH5-1和DH5-2), 平均相似性为20.1%,主要贡献种有背蚓虫(Notomastus latericeus) (贡献率为15.4%, 下同)、长吻沙蚕(Glycera chirori) (13.9%)、长叶索沙蚕(Lumbrineris longifolia) (12%)和蜈蚣欧努菲虫(Onuphis geophiliformis) (8.7%)。群组2包括调查海域最南端的DH4-5和DH5-3站, 平均相似性为30.3%, 贡献种包括阳遂足科(Amphiuridae)的一种倍棘蛇尾(40%)、尖叶长手沙蚕(Magelona cincta) (20%)、双栉虫(Ampharete acutifrons)(20%)和蜈蚣欧努菲虫(20%)。群组3由长江口北部海域的I1和I2站组成, 平均相似性为32.6%, 主要贡献种是塞切尔泥钩虾(Eriopisella sechellensis) (42.9%)。群组4包含K2、M3和M4站位, 平均相似性为 33.2%, 中华内卷齿蚕(Aglaophamus sinensis) (69.2%)是主要贡献种。群组 5包含K4和M2站位, 平均相似性为28.6%, 贡献种是奇异稚齿虫(Paraprionospio pinnata)(100%)。M1、DH3000-4和DH4-2站则各自成为一个群组。
2.5 ABC曲线分析
丰度/生物量比较法, 即ABC法能较客观地反映海洋环境污染对底栖生物群落稳定性的影响。一般认为, 稳定的海洋环境中, 底栖生物群落结构近乎平衡, 生物量曲线始终位于丰度曲线的上方, 即丰度比生物量具有更高的多样性; 群落受到中度扰动时, 生物量占优势的大个体消失, 使丰度和生物量的分布优势难以区分, 生物量曲线和丰度曲线出现部分交叉或重合; 当底栖生物群落遭到严重的环境污染或扰动时, 群落逐渐由一种或几种小个体种类占优势, 导致丰度分布比生物量分布更显优势, 丰度曲线位于生物量曲线之上(Warwick and Clarke, 1994; 田胜艳等, 2006)。图5(a)显示长江口邻近海域夏季大型底栖生物的 ABC曲线, 其生物量曲线虽略高于丰度曲线, 但前者的起始较低, 生物量的优势不明显, 说明调查海域的大型底栖生物群落受到了轻微扰动。不同海区大型底栖生物的ABC曲线如图5(b)~图5(d)所示。可以看出,长江口北部海区和舟山海区的ABC曲线情况与调查海域的总体情况基本一致, 生物量曲线起始较低, 大型底栖生物群落受到轻微扰动; 长江口海区的 ABC曲线显示环境状况较好, 大型底栖生物群落结构基本稳定。
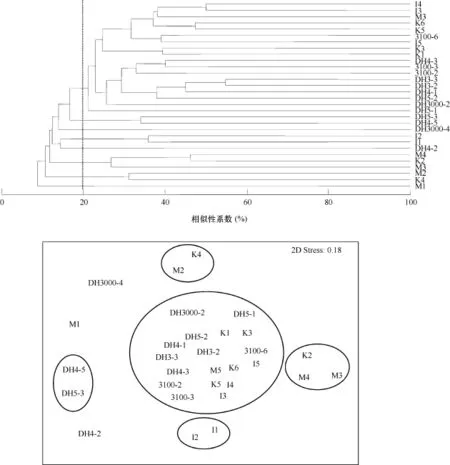
图4 长江口邻近海域大型底栖生物群落结构聚类分析和nMDS排序Fig.4 Cladogram of Bray-Curtis similarity and nMDS ordination of macrobenthic community in adjacent sea of Yangtze River estuary
3 讨论
3.1 种类组成和优势种变化
本文的研究共采集大型底栖生物 135种,各主要类群物种数占采集总物种数的比例由高到低依次为多毛类动物、甲壳动物、棘皮动物、软体动物。各类群所占比例的高低顺序与李宝泉等(2007)(2002年 9月航次)、Shou等(2013)(2006年7~8月、2006年12月~2007年1月、8月和10~11月共4个航次)、Chao等(2012)(2010年5月、8月、11月和2011年2月共 4个航次)、袁一鸣等(2015)(2011~2014年的8月共4个航次)的研究结果不尽相同。差别之处主要在于甲壳动物和软体动物种类数的高低, 这可能是由调查年份和季节不同造成的, 也有可能是由调查海域的范围及站位设定不同造成的。此外, 与20世纪80年代的数据相比, 环境适应能力较强、耐污的多毛类动物在群落中所占比例明显增加(刘录三等, 2012),本调查中的优势种和常见种也全部是多毛类。以往的研究显示, 优势种和常见种种类在不同动物类群中基本呈均衡分布, 如甲壳动物中的豆形短眼蟹(Xenophthalmus pinnotheroides)、葛氏长臂虾(Palaemon gravieri); 软体动物中的秀丽织纹螺(Nassarius dealbatus)、金星蝶饺蛤(Trigonothracia jinxingae); 棘皮动物中的日本倍棘蛇尾(Amphioplus japonicus)等(刘瑞玉等, 1992; 李宝泉等, 2007; 刘录三等, 2008)。随着长江口邻近海域底栖生境的不断变迁和人类活动的持续影响, 各生态类群的优势地位已发生显著更替, 个体较小、生长周期短的多毛类动物取代了个体较大、生长周期较长的其他动物, 成为最主要的优势类群(刘录三等, 2008; 袁一鸣等, 2015)。

图5 长江口邻近海域大型底栖生物ABC曲线Fig. 5 ABC plots of macrobenthos in adjacent sea of Yangtze River estuary
3.2 丰度和生物量
一定时期内, 特定海域中底栖生物丰度和生物量的变化可反映其群落的变化, 进而可推断环境的变化。2014年, 长江口邻近海域夏季大型底栖生物的平均丰度是 122.2ind./m2,低于1999~2001年(1999年5月为333.24ind./m2, 2000年11月为213.08ind./m2, 2001年5月为411.91ind./m2)(吴耀泉和李新正, 2003)、2002年秋季(231.5ind./m2)(李宝泉等, 2007)、2004年(2月、5月、8月、11月分别为375.0ind./m2、623.0ind./m2、309.5ind./m2、781.7ind./m2)(吴耀泉, 2007)、2009年春季(212.4ind./m2)(刘录三等, 2012)的调查结果, 与 2005~2006年(146.4ind./m2)(刘录三等, 2008)、2011年夏季(109.3ind./m2)(袁一鸣等, 2015)的数据基本持平, 高于 2010年春季(70ind./m2)(刘录三等, 2008)、2012年和2013年夏季(分别为82.8ind./m2、59.2ind./m2)(袁一鸣等, 2015)的调查结果。这表明, 1999~2014年长江口邻近海域大型底栖生物的平均丰度在不同年份和季节存在一定波动。
调查海域夏季大型底栖生物的平均生物量是7.8g/m2, 远低于1999~2001年(1999年5月为 14.04g/m2, 2000年 11月为 25.65g/m2, 2001年 5月为 28.14g/m2)(吴耀泉和李新正, 2003)、2002年秋季(27.66g/m2)(李宝泉等, 2007)、2004年(2月、5月、8月、11月分别为19.7g/m2、23.4g/m2、12.7g/m2、19.6g/m2)(吴耀泉, 2007)、2005~2006年(12.8g/m2)(刘录三等, 2008)的调查结果, 与2009年和2010年春季(分别为10.9g/m2、9.1g/m2)(刘录三等, 2012)的调查结果相近, 高于2011~2013年夏季(分别为5.9g/m2、3.5g/m2、1.6g/m2)(袁一鸣等, 2015)的调查数据, 表明长江口邻近海域大型底栖生物的平均生物量在不同年份和季节都有差异。
丰度和生物量的空间分布一致性较高,均呈不连续的斑块或镶嵌状分布。研究表明,海域底栖生物的分布格局与沉积物类型密切相关(Gray, 1974; 罗秉征和沈焕庭, 1994; 寿鹿, 2013), 长江口邻近海域多为黏土质沉积物, 分为 4种类型: 细砂、砂-粉砂-黏土、黏土质粉砂和粉砂质黏土, 且它们的分布格局呈斑块状, 因此调查海域物种的丰度和生物量分布也表现为斑块状格局(杨光复和吴景阳, 1994)。
长江口邻近海域不同海区间平均丰度大小依次为长江口北部海区(165.9ind./m2)>舟山海区(115.6ind./m2)>长江口海区(85ind./m2);平均生物量大小依次为舟山海区(9.4g/m2)>长江 口 北 部 海 区 (8.5g/m2)>长 江 口 海 区(5.4g/m2)。长江口海区的底质环境受长江径流及航道疏浚等的影响, 沉积速率高, 底质表面浮泥厚, 沉积环境不稳定, 对底栖生物的生存构成威胁, 所以其平均丰度和生物量最低。
3.3 多样性和群聚结构
本次调查所获得的长江口邻近海域大型底栖生物群落的 3个多样性指数 H'、D和 J分别为3.19、3.82和0.89。其中, H'值高于吴耀泉和李新正(2003)(1999年 5月 为 2.41、2000年11月为2.87、2001年5月为2.88)、李宝泉等(2007)(2002年9月为2.06)、刘录三等(2012)(2009年4月为1.36、2010年3月为1.08)的研究结果, 反映了长江口邻近海域生态环境有所恢复, 这应该与近年来长江口海域底栖生物增殖流放、长江口滨海湿地生态修复等措施密切相关(袁一鸣等, 2015)。
从空间上来看, 舟山海区的多样性指数H'、D和J最高(表4), 可能的原因是多种海流在此交汇, 使水体层化, 有利于悬浮泥沙迅速沉降, 提高了水体中光的利用率, 加速了浮游植物群落的增长, 能够为大型底栖生物提供丰富的饵料, 同时海区底层环境较为稳定, 有利于大型底栖生物的生长和繁衍(寿鹿, 2013)。
对长江口邻近海域的大型底栖生物进行群落CLUSTER聚类和nMDS排序分析, 结果表明, 站位间的相似性水平较低, 没有相似性水平超过 60%的站位, 与李宝泉等(2007)在2002年9月的调查结果一致。在20%的相似性水平上, 调查海域的大型底栖生物被划分为8个群组, 无明显规律可循, 可以看作是在一个较大的斑块上镶嵌一些较小的斑块。长江口邻近海域水文环境复杂, 形成不同的小生境, 不同小生境栖息着不同结构的底栖生物群落, 造成群落间相似性都较低(李宝泉等, 2007)。
3.4 长江口邻近海域大型底栖生物群落受污染扰动情况
ABC曲线通过将丰度曲线和生物量曲线进行对比, 来反映海洋环境污染对底栖生物群落结构的影响, 目前其已广泛应用于渔业资源和底栖生物群落的研究。对长江口邻近海域大型底栖生物群落进行 ABC曲线分析, 表明调查海域受到了一定程度的扰动。2002年,李宝泉等(2007)在长江口海域大型底栖生物群落进行ABC曲线分析时发现, 距离长江口较近的站位受到了轻微污染扰动, 较远的站位几乎未受影响。长江口邻近海域的生态环境本身就比较复杂, 受到扰动的人为因素较多,包括大型水利工程、航道整治工程、滩涂围垦和污水排放等, 造成底栖生境被破坏, 使大型底栖生物的多样性和群落稳定性下降。今后还需对该海域进行长期连续的监测, 并结合水文、底质类型、溶解氧和重金属等多种环境因子的监测, 以期为长江口邻近海域大型底栖生物群落的演替提供可靠的科学依据, 维持底栖生态系统的健康发展。
致谢 王金宝、董栋、寇琦、隋吉星、甘志彬、龚琳、徐勇、王跃云、孙悦等参加了本文研究的野外考察、实验室样品处理和鉴定等工作; 中国科学院海洋生物标本馆管理员为本文研究的样品处理给予了大力支持和协助,在此一并致以衷心的感谢。
蔡文倩, 孟伟, 刘录三, 等. 2013. 长江口海域底栖生态环境质量评价——AMBI和M-AMBI法. 环境科学, 34(5): 1725-1734
贾海波, 曹柳燕, 胡颢琰. 2014. 长江口及其邻近海域春季大型底栖生物群落与环境因子的典范对应分析. 中国环境监测, 30(1): 93-97
李宝泉, 李新正, 王洪法, 等. 2007. 长江口附近海域大型底栖动物群落特征. 动物学报, 53(1): 76-82
李建生, 李圣法, 程家骅. 2006. 长江口渔场鱼类组成和多样性. 海洋渔业, 28(1): 37-41
李新正, 王洪法, 张宝琳, 等. 2012. 黄海和东海大型底栖生物生态学研究进展. 可持续发展, 2: 80-102
刘录三, 孟伟, 田自强, 等. 2008. 长江口及毗邻海域大型底栖动物的空间分布与历史演变. 生态学报, 28(7): 3027-3034
刘录三, 郑丙辉, 李宝泉, 等. 2012. 长江口大型底栖动物群落的演变过程及原因探讨. 海洋学报, 34(3): 134-154
刘瑞玉, 徐凤山, 孙道元, 等. 1992. 长江口区底栖生物及三峡工程对其影响的预测. 海洋科学集刊, (33): 237-247
刘勇, 线薇薇, 孙世春, 等. 2008. 长江口及其邻近海域大型底栖动物生物量、丰度和次级生产力的初步研究. 中国海洋大学学报, 38(5): 749-756
罗秉征, 沈焕庭. 1994. 三峡工程与河口生态环境.北京: 科学出版社
寿鹿. 2013. 长江口及邻近海域大型底栖生物群落生态学研究. 南京: 南京师范大学博士学位论文
孙亚伟, 曹恋, 秦玉涛, 等. 2007. 长江口邻近海域大型底栖生物群落结构分析. 海洋通报, 26(2): 66-70
田胜艳, 于子山, 刘晓收, 等. 2006. 丰度/生物量比较曲线法监测大型底栖动物群落受污染扰动的研究. 海洋通报, 25(1): 92-96
王金辉, 黄秀清, 刘阿成, 等. 2004. 长江口及邻近水域的生物多样性变化趋势分析. 海洋通报, 23(1): 32-39
王延明, 方涛, 李道季, 等. 2009. 长江口及毗邻海域底栖生物丰度和生物量研究. 海洋环境科学, 28(4): 366-370, 382
吴耀泉. 2007. 三峡库区蓄水期长江口底栖生物数量动态分析. 海洋环境科学, 26(2): 138-141
吴耀泉, 李新正. 2003. 长江口区底栖生物群落多样性特征//中国甲壳动物学会. 甲壳动物学论文集,第4辑. 北京: 科学出版社
徐兆礼, 陈亚瞿. 1989. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志, 8(4): 13-15
杨光复, 吴景阳. 1994. 沉积与地球化学//罗秉征, 沈焕庭. 三峡工程与河口生态环境. 北京: 科学出版社
杨金龙, 周轩, 郭行磐, 等. 2014. 长江口潮下带大型底栖动物的群落结构特征. 水产科技情报, 41(4): 192-198
叶属峰, 纪焕红, 曹恋, 等. 2004. 河口大型工程对长江河口底栖动物种类组成及生物量的影响研究.海洋通报, 23(4): 32-37
袁一鸣, 秦玉涛, 刘材材, 等. 2015. 长江口海域夏季大型底栖动物群落结构分析. 水产学报, 39(8):1107-1121
周红, 张志南. 2003. 大型多元统计软件 PRIMER的方法原理及其在底栖群落生态学中的应用. 青岛海洋大学学报, 33(1): 58-64
Chao M, Shi Y R, Quan W M, et al. 2012. Distribution of benthic macroinvertebrates in relation to environmental variables across the Yangtze River Estuary, China. Journal of Coastal Research, 28(5): 1008-1019
Clarke K R. 1993. Non-parametric multivariate analyses of changes in community structure. Austral Ecology, 18(1): 117-143
Gray J S. 1974. Animal-sediment relationships. Oceanography and Marine Biology: An Annual Review, 12: 223-261
Margalef R. 1968. Perspective in Ecological Theory. Chicago: University of Chicago Press
Pielou E C. 1966. Species-diversity and patterndiversity in the study of ecological succession. Journal of Theoretical Biology, 10(2): 370-383
Shannon C E, Wiener W. 1949. The Mathematical Theory of Communication. Urbana: University of Illinois Press
Shou L, Zeng J N, Liao Y B, et al. 2013. Temporal and spatial variability of benthic macrofauna communities in the Yangtze River estuary and adjacent area. Aquatic Ecosystem Health & Management, 16(1): 31-39
Warwick R M, Clarke K R. 1994. Relearning the ABC: taxonomic changes and abundance/biomass relationships in disturbed benthic communities. Marine Biology, 118(4): 739-744
Characteristic of Summer Macrobenthic Community in Adjacent Sea of Yangtze River Estuary
YANG Mei1,2, LI Xin-Zheng1,2*, WANG Hong-Fa1, ZHANG Bao-Lin1
(1. Department of Marine Organism Taxonomy and Phylogeny, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. Laboratory of Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266200, China) *Corresponding author, E-mail: lixzh@qdio.ac.cn
In this paper, the characteristic of the macrobenthic community in adjacent sea of Yangtze River estuary was studied, based on data collected from cruise in June(summer) of 2014. A total of 135 macrobenthic species were identified, of which, 66 species were Polychaeta, 33 were Crustacea, 14 were Echinodermata, 13 were Mollusca, and the remaining 9 species belong to other animal groups. The average abundance of the macrobenthos in adjacent sea of Yangtze River estuary was 122.2 ind./m2, and the average biomass was 7.8g/m2. The abundance, biomass, Shannon-Wiener index(H'), Margalef's richness index(D), and Pielou index (J ) were not significantly spatial difference. Eight macrobenthic assemblages were identified on the 20% similarity level, with low similarity among stations. The results of the ABC analysis show that with the rapid economical development of coastal zones and the impact of human activities, the stability of the benthonic community in adjacent sea of Yangtze River estuary is disturbed slightly, therefore, long-term continuous monitoring is recommended and effective comprehensive management measures need to be explored to result in the healthy and sustainable development of the marine ecosystem in adjacent sea of Yangtze River estuary.
Yangtze River estuary; macrobenthos; diversity; community structure
P717
10.12036/hykxjk20160725001
* 资助项目: 国家自然科学基金 (41176133) ; 中国科学院战略性先导科技专项(A类)(XDA11020303)。杨梅,女, 博士后, E-mail: yangmeiquietsnow@ 163.com
① 通讯作者: 李新正, 男, 研究员, 从事海洋底栖生物生态学研究, E-mail: lixzh@qdio.ac.cn
2016-07-25, 收修改稿日期: 2016-07-29
