中国海洋底栖生物学发展回顾与展望*
2016-03-15张均龙史本泽徐奎栋
张均龙 史本泽 赵 峰 徐奎栋
(中国科学院海洋研究所 海洋生物分类与系统演化实验室, 青岛 266071)
中国海洋底栖生物学发展回顾与展望*
张均龙 史本泽 赵 峰 徐奎栋①
(中国科学院海洋研究所 海洋生物分类与系统演化实验室, 青岛 266071)
海洋底栖生物是海洋生态系统中的重要组成部分, 在底层生态系统的能量流动和物质循环过程中发挥着重要作用。我国海洋底栖生物学起步较晚, 经过数十年的发展, 已取得许多研究进展和成果, 但目前学科发展仍存在很多不足。总体而言,我国海洋底栖生物研究力量相对薄弱和分散, 仍为相对弱势的一个学科, 亟需一个交流与合作的平台来整合业内的研究力量和资源, 促进我国底栖生物学的研究与发展。本文综合分析了大型底栖生物、小型底栖生物和微型底栖生物研究的现状和发展趋势,并就未来发展提出了建议。
底栖生物; 海洋底栖生物学分会; 生态学; 分类学; 多样性
海洋的底层环境是继海洋水体之后, 地球上的第二大生物栖息地, 是海洋中生物多样性最为丰富的生态类型, 其生态功能极为重要。但是人类对这一广阔生物栖息地的了解甚少, 甚至不如对月球表面了解得多(Gray and Elliott, 2009)。海洋底栖生物占据海底这一广袤的生境, 从潮间带到深海均有分布,已知的海洋无脊椎动物中 90%为底栖生物。海洋底栖生物(benthos)是生活在海洋底表或沉积物中的各种生物, 包括底栖动物(zoobenthos)、底栖植物(phytobenthos)和底栖微生物(benthic microorganisms)。底栖动物根据其生态类型可分为底上动物(epifauna)、底内动物(infauna)和底游动物(nektobenthos或 hyperbenthos)。底上动物主要是指在底表移动生活或附着或固着在岩石、贝壳或植物等基物上的底栖动物; 底内动物是指埋栖于泥沙或在底内穴居、钻孔的底栖动物; 底游动物包括栖息于底层的鱼类和活动能力较强的无脊椎动物。海洋底栖植物主要指在海洋底栖的绿藻、褐藻、红藻等大型海藻, 有时也包括底栖硅藻等微型生物。底栖微生物通常是指个体微小的原核或真核的单细胞自养或异养微型生物, 主要包括细菌、真菌、原生动物和微藻。底栖生物根据其大小(图1见文后彩图), 又可分为微型底栖生物(microbenthos)、小型底栖生物(meiobenthos)和大型底栖生物(macrobenthos)。
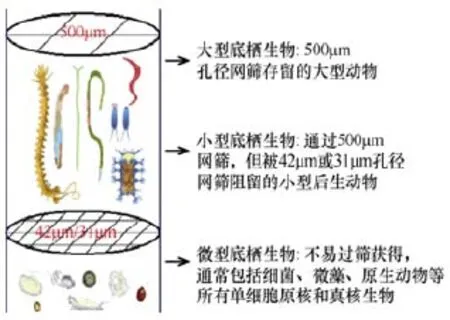
图1 底栖生物过筛分选获得的各粒级组成示意图(见书后彩图15)Fig.1 Diagram showing major benthos groups sorted by different particle size fraction
大型底栖动物(macrofauna)通常是指能被0.5mm 孔径网筛截留的底栖动物, 其主要类群包括刺胞动物(Cnidaria)、多毛类环节动物(Annelida)、软体动物(Mollusca)、甲壳动物(Crustacea)及棘皮动物(Echinodermata), 此外还包括常见的海绵(Porifera)、苔藓虫(Bryozoa)、星虫(Sipuncula)、螠虫(Echiura)、纽虫(Nemertea)和底栖鱼类等。
小型底栖生物又称小型底栖动物(meiofauna),是指分选时可通过 0.5mm 网筛, 但被0.042mm 网筛蓄留的动物, 深海生态研究中采用 0.031mm 作为小型底栖生物的下限(Giere, 2009)。小型底栖生物是底栖环境中数量最为优势的后生动物类群, 包含暂时性和永久性小型底栖生物, 至少有 22个门类包含小型底栖生物的分类阶元, 其中动吻动物(Kinorhyncha)、颚咽动物(Gnathostomulida)、缓步动物(Tardigrada)和有甲动物(Cingulata)等门类是小型底栖生物特有的。线虫(Nematoda)通常是丰度上最占优势的类群, 其次是以猛水蚤(Harpacticoida)为主的底栖桡足类(Copepods) (McIntyre, 1969; Coull, 1999)。
微型底栖生物是指生活于沉积物表面和底内的所有单细胞原核和真核微型生物, 包括光合自养和异养(含混合营养)的类群。按个体大小划分, 微型底栖生物涵盖粒径0.2~2μm的微微型、2~20μm的微型和>20μm的微小型底栖生物; 按功能类群划分, 微型底栖生物包括底栖原核微生物、真核微藻及原生动物等(类彦立和徐奎栋, 2011; 徐奎栋, 2011)。此外,低等真菌及营寄生生活的毫微微型(<0.2μm)病毒也是微型底栖生物的组成部分, 还是海洋底栖生态系统的重要组分。
1 底栖生物的生态功能
底栖生物对底层生态系统的能量流动和物质循环过程, 如营养物的循环, 污染物质的分解、扩散和掩埋, 以及次级生产力等有重要作用(Snelgrove, 1998; Ellingsen and Gray, 2002; 李新正, 2011)。微型底栖生物、小型底栖生物和大型底栖生物各司其职, 构成了完整的海洋底栖生物食物网(图2), 并完成海洋底层的物质循环和能量流动过程。因此, 对底栖生物的研究将有助于加深我们对海洋底层生物多样性和生态系统的了解。
不同种类底栖生物对环境条件的适应性及对污染等不利因素的耐受力和敏感程度不同, 海洋大型底栖生物生活史较长、活动性差,而微型和小型底栖生物的生活周期较短、繁殖力强, 根据这些特点, 利用底栖生物的种群结构、优势种类、数量等参量可以确切反映水体和底层环境的质量状况。底栖生物在环境污染监测中通常被用作环境变化的指示生物。
通常, 大型底栖生物因其具有较大的经济意义而更受关注。大型底栖生物中许多种类是水产养殖和捕捞的对象, 有些是经济鱼虾蟹的天然饵料, 具有重要的经济价值; 多数大型底栖生物长期生活在底表或底层沉积物中, 具有区域性强、迁移能力弱等特点, 对于环境污染及变化通常少有回避能力, 其群落破坏后的重建需要相对较长的时间(Snelgrove, 1998; Ellingsen and Gray, 2002; 李新正, 2011)。小型底栖生物作为微型底栖生物和大型底栖生物的中间桥梁, 在整个底栖生态系统中的物质循环和能量流动过程中起着承上启下的作用。小型底栖生物主要摄食沉积环境中的有机碎屑和细菌、硅藻、纤毛虫等微型生物, 调节微型生物的生产过程; 同时, 又是大型底栖生物的重要食物, 是许多经济鱼类、虾类和贝类等幼体的重要饵料, 其变化会直接影响到这些重要经济类群的数量(Coull, 1999; 张志南等, 2004)。微型底栖生物数量极为庞大, 在生态系统的初级生产、有机质利用及营养物质循环中起着基础作用, 是底栖微食物网中初级生产力向高营养级转换的枢纽 (徐奎栋, 2011)。

图2 海洋底栖生物食物网示意图(仿自徐奎栋, 2011)Fig.2 Schematic diagram of benthic food web (from Xu, 2011)
2 海洋底栖生物学研究简史
2.1 国际研究进展
海洋生态学开始于18世纪初英国的海洋生物学家零星的底栖生物调查。19世纪, 西欧各国相继进行多次大范围的海洋生物调查。20世纪初, 海洋大型底栖生物的研究主要包括对群落组成及其稳定性和多样性、群落演替、能量转换等方面的基础性研究。在此期间, 国际上才开始出现对小型底栖生物的系统研究,比大型底栖生物的研究晚了将近一个半世纪(Higgins and Thiel, 1988)。这些对小型底栖生物的研究主要还处于发现和分类鉴定阶段。20世纪50年代后, 大型底栖生物进入定量研究、生物群落及生物多样性研究阶段, 并通过对底栖生物群落进行长期调查以进行环境监测,小型底栖生物也进入系统分类学加速发展和群落定性描述阶段。20世纪60年代初, 人们才普遍使用大型底栖动物区系、物种的出现、丰度、生物量等来评价海洋生物环境状况。到20世纪80年代, 提出了有关底栖生物的种种假说, 各类假说的验证成为研究的主要驱动力, 实验生态学和生物海洋学迅速发展。进入21世纪, 底栖生物研究的时空尺度已经扩大到全球, 并以年际和 10年际的时间系列为重点, 联系全球变化, 特别是以气候异常和人类活动的影响为切入点, 通过现场观测和受控试验相结合, 探讨底栖生物的功能响应越来越受到学者的重视。随着生态调查范围的扩大,多元统计技术和计算机的广泛应用, 海洋底栖生物群落的研究已由定性描述发展到了定量解析阶段。底栖群落生态学的研究在经历了一个多世纪的发展过程后, 逐步由单纯的野外观测和描述转入实验生态、对系统演化的考虑及污染和扰动对群落结构的影响评价。
目前, 海洋底栖生物多样性主要的进展和趋势如下: ①从局域尺度和区域尺度到全球尺度的海洋底栖生物多样性格局及其控制机制已取得明显进展。②现场观测和受控实验相结合, 探讨底栖生物多样性对气候异常和人类扰动的响应是另一个热点。③底栖生物在大的时间尺度下发生变化的相关因素也备受关注。特别是近些年来, 人类活动和全球气候变化加剧了生态环境的改变, 外部因素对种群和群落的影响更受关注。④随着深海探测技术的成熟和应用, 越来越多的研究开始涉及深海和极端生境的底栖生物生态研究, 包括揭示新物种和新生命现象、探讨其生物地理学特点、评估底栖生物在区域和全球海域的生物多样性。⑤分子生物学技术在系统发育分析和遗传多样性研究中得到广泛应用, 对底栖生物各类群的系统发育地位和关系进行了重新审视, 获得了新的认识。⑥国际合作的全球化和新技术应用的大趋势。
2.2 国内研究进展
我国底栖生物研究起步较晚, 经过数十年的发展, 已取得许多研究进展和成果, 但目前学科发展仍存在很多不足。刘瑞玉(1990)、李新正(2011)、张志南等(2004)及徐奎栋(2011)分别对大型、小型和微型底栖生物研究的进展做过详细综述, 在此不做详细赘述, 仅简要回顾其发展历史并介绍重要进展。
2.2.1 大型底栖生物研究进展
我国首次海洋生物调查从 1935~1936年“胶州湾海产动物采集团”的调查开始, 先后发表四期采集报告及食用软体动物报告(张玺, 1935; 张玺和相里矩, 1936; 张玺和马绣同, 1936, 1949)。我国海洋底栖生态学的研究起步较晚, 是以 1956年中苏黄海、东海和海南岛潮间带生态学开始的, 俄罗斯学者古丽亚诺娃(1958)和我国底栖生态学奠基人刘瑞玉等对黄海潮间带的大型底栖生物进行了研究。1957年, 我国第一艘海洋综合调查船“金星号”开始服役, 展开了全国海洋多学科的综合调查, 即全国近海海洋综合调查(1958~1960年)(刘瑞玉和徐凤山, 1963)。中越北部湾海洋综合调查研究也获得了多学科资料和标本,填补了学科领域的空白(1959~1960年, 1962年)。我国在这一阶段的研究主要是调查底栖生物多样性的本底资料并开展分类工作, 从而对各海区大型底栖动物区系的特点有了基本了解。
20世纪80年代以来, 通过开展国际合作,我国的大型底栖生物学的研究取得很大发展。将底栖生物群落与沉积类型相联系, 进行定性定量的描述, 并进行东海大陆架调查(1978~1979年)、中国海岸带和海涂资源综合调查(1980~1986年)。Liu等(1983)、刘瑞玉等(1986)研究了黄、东海大型底栖动物的生态特点, 并对黄东海底栖生物区系进行了划分, 讨论了底栖生物与温度、盐度、海流及沉积物的关系。唐质灿和徐凤山(1978)、江锦祥等(1985)对东海大陆架底栖生物的数量分布和群落进行了研究。Boesch等 (1986) 讨论了东海大陆架底栖生物在沉积过程中的作用。后来, 还在长江口及其邻近水域进行了长江三峡工程对河口生物学和渔业资源影响的调查(刘瑞玉等, 1987)、中国海岛资源综合调查(1990~1994年)、中韩黄海及其附近水域的调查(刘瑞玉等, 1996)、126大陆架专项调查(1997~2000年)、东黄海生态系统动力学与生物资源可持续利用(1999~2004年)、中国近海海洋综合调查与评价专项(即908专项调查, 2004~2009年) 和正在进行中的中国近海水母暴发的关键过程、机理及生态环境效应等。
其中, 对黄海大型底栖生态研究相对较多, 刘瑞玉(1990)、李新正(2011)对黄海的海洋大型底栖生态学进行了综述。张均龙等对比了 50年间黄海大型底栖动物群落的变化, 结果显示, 黄海软体动物多样性降低, 群落受到了较为严重的干扰, 群落结构发生了较大变化, 冷水种种群萎缩衰退, 分布范围缩小, 有的甚至已经枯竭, 暖水性种类分布范围北移,从东海扩布到了黄海南部; 黄海软体动物在近 50年中在向小型化和种数减少趋势发展,现在的种类以生长周期短、个体小、数量多的物种为主(Zhang et al., 2016a, 2016b)。
另外, 对渤海(吴耀泉和张宝林, 1990; 孙道元和刘银城, 1991; 于子山等, 2001; 韩洁等, 2004; Zhou et al., 2007)、长江口(孙道元等, 1992; 李宝泉等, 2007; 章飞军等, 2007; Meng et al., 2007; 刘录三等, 2008; Liu and Zheng, 2010)、胶州湾(刘瑞玉, 1992; 李新正等, 2005, 2006; 于海燕等, 2006)、福建和浙江沿海(李荣冠和江锦祥, 1989; 鲍毅新等, 2006; 寿鹿等, 2009; 赵永强等, 2009)等地的底栖生物资源的调查也较多, 并研究了影响海洋底栖生物分布的生态因子, 为进一步研究底栖生态系统的结构和功能提供了重要参考。
2.2.2 小型底栖生物研究进展
我国小型底栖生物的研究起步较晚、研究历史较短。陆鼎恒(1934)记述了黄海山东半岛北岸的动吻类新种是我国关于海洋小型底栖生物最早的报道, 但此后直到20世纪80年代几乎没有相关的研究报道。20世纪70年代末和 80年代初, 在开放政策和学者外出访问及国际合作引入的推动下, 包括“中美”(1985~ 1987年), “中英”(1995~1997年、1998~2000年),“中法” (2004~2006年)和“中日” (1997~2006年)等国际联合调查, 我国的小型底栖生物研究取得了快速发展 (张志南等, 2004; 黄勇, 2005)。在张志南教授的带领下, 我国的小型底栖生物研究从黄渤海域的潮间带、养虾池和浅海陆架开始, 逐步扩展到其他海域, 并在分类学、多样性和生态学研究领域取得了一系列较为显著的进展。
渤海是我国陆架区海域小型底栖生物生态系统研究的开端, 也是我国目前研究较为全面系统的海域。此后, 研究涉及了渤海的小型底栖生物的类群组成、丰度、生物量、生产量、空间分布及其与环境因子的关系等, 对渤海的小型底栖生物生态学及线虫和底栖桡足类的群落结构和多样性做了较为全面系统的研究(张志南等, 1989, 1993; 张志南, 2000; 慕芳红等, 2001a, 2001b; Guo et al., 2001a, 2001b, 2002; 郭玉清等, 2002, 2003; Hua et al., 2009; Zhang et al., 2010)。
黄海的小型底栖生物生态学研究是迄今为止所有海域中最多的。黄海的小型底栖生物生态学研究始于钱国珍等(1992)对青岛湾潮间带有机质污染区的调查; 之后在青岛潮间带的砂质、泥砂质和泥质沉积物中陆续开展了大量研究(张志南等, 1993; 范士亮等, 2006, 2011; Du et al., 2009; 杜永芬等, 2011; 华尔等, 2012), 而黄海其他沿海地区潮间带则鲜有研究报道; 此外, 张志南等(1993)和张艳等(2005)还对潮间带的附生于海藻上的小型底栖生物进行了研究。黄海潮下带的小型底栖生物研究始于张志南等(2001)对胶州湾的调查,之后在黄海陆架区展开了一些研究(林岿璇等, 2004; 张志南和周红, 2004; 黄勇, 2005; 刘晓收, 2005; 黄勇和张志南, 2007; 吴绍渊和慕芳红, 2009;张艳, 2009; 吴秀芹等, 2010; 王家栋等, 2011;于婷婷和徐奎栋, 2013), 这些研究丰富了对黄海潮下带, 尤其是陆架区小型底栖生物类群组成和数量分布的认识, 为进一步了解其动态和功能提供了必要的背景资料。
与渤海和黄海相比, 东海海域的小型底栖生物研究仍然偏少, 研究范围主要集中在滩涂虾池和红树林等潮间带(蔡立哲和李复雪, 1998; 蔡立哲等, 2000a; 郭玉清, 2008; 郭玉清和叶兆弘, 2008)、台湾海峡 (蔡立哲等, 2000b, 2001; 方少华等, 2000)和长江口及其邻近海域(Fleeger et al., 1986; 华尔等, 2005; Hua et al., 2006, 2009; 王小谷等, 2010; 于婷婷和徐奎栋, 2013; 史本泽等, 2015), 涉及小型底栖动物的丰度、生物量及粒径谱等。迄今为止, 我国对该海域的研究偏少, 缺乏季节和年际变化的资料, 对其数量分布及变动尚缺乏足够的认识。
总体上, 南海的开始最晚, 也是目前小型底栖生物研究最少的海域, 且已有研究主要集中于香港沿岸(Zhou, 2001; Liu et al., 2008, 2009, 2011a, 2011b; Zhou and Zhang, 2008)、北部湾(蔡立哲等, 2000b; 王彦国等, 2011; Cai et al., 2012)和南海北部陆架海域(杜永芬等, 2010), 而关于大陆坡及深海海域则尚无研究资料。
2.2.3 微型底栖生物研究进展
相较于大型和小型底栖生物, 微型底栖生物的个体微小、多样性更高、功能更复杂,且大部分底栖微型生物个体脆弱, 不易从沉积物中提取和培养, 研究极为欠缺。类彦立和徐奎栋编制完成的《海洋微型底栖生物调查规范》(HY/T 140—2011)填补了现行《海洋调查规范》(GB/T 12763.6—2007)中尚缺的微型底栖生物部分。该规范为微型底栖生物各类群的定量提取与定性分析提供了技术方案, 提出了用于海洋微型底栖生物分析的 Ludox-QPS法 (Xu et al., 2010), 该方法可以同时定量及定性分析海洋底栖微型生物多样性, 尤其是底栖原生动物和真核微藻的多样性。Xu等(2010)和 Du等(2009)通过测试不同类型的沉积物发现, 该方法对原生动物和真核微藻的提取率可达97%以上。
以此为基础, 开展了潮间带和近海的微型底栖生物研究, 揭示了黄海近岸潮间带和陆架海区的微型底栖生物群落结构特点。在黄海陆架海区共鉴定底栖纤毛虫等近200种, 发现前口纲纤毛虫为该海区最为优势的类群,功能群上以肉食性纤毛虫占优势, 不同于黄海近岸潮间带以藻食性为主的功能群构成(孟昭翠和徐奎栋, 2013; 周百灵等, 2015)。从微型底栖生物垂直分布来看, 底栖原生动物垂直分层明显, 80%左右的丰度和生物量由表层0~2 cm贡献 (Meng et al., 2012), 底栖真核微藻和小型鞭毛虫同样呈现明显的垂直分层分布; 而粒级较小的底栖微型生物, 如底栖细菌、微型鞭毛虫则分层不明显(Meng et al., 2011)。总体上, 黄海微型底栖生物现存量十分庞大, 单位体积内的底栖细菌、底栖微藻和底栖原生动物的现存量高出水体中相应类群2~3个数量级(Meng et al., 2011)。在同一海域,底栖细菌和底栖原生动物在生产量和新陈代谢率上明显高于小型底栖生物。黄海单位面积沉积物中各微型生物类群的总生物量和约100m深的水体相当, 数量如此丰富无疑联系着功能上的巨大作用 (徐奎栋, 2011)。
海洋微型底栖生物多样性研究主要侧重于物种多样性及群落结构与分子多样性, 而生境多样性的研究则相对较少。显著的进展体现在原核微生物的分子多样性, 以及真核微藻的物种多样性, 而底栖原生动物则因采样、保存及分类鉴定等诸多限制而研究滞后。随着我国海洋微型底栖生物行业标准的发布实施,越来越多的海洋调查和研究开始关注这些个体微小、数量和生物量巨大的类群的生物多样性和生态功能, 以及其在底栖生态系统和底栖-浮游耦合中的作用。
3 展望
我国自海洋普查至今的数十年间, 海洋底栖生物的研发力量已在国内绝大多数涉海单位发展起来, 并逐渐在海洋的生物生态调查、海洋生物多样性、海洋环境监测等领域占有一席之地。然而, 总体来讲, 海洋底栖生物学在我国仍是一个相对弱势的学科, 研究力量相对薄弱和分散, 研究人员间缺少必要的交流和沟通, 尤其缺乏一个交流与合作的平台。即将成立的中国海洋湖沼学会海洋底栖生物学分会, 将整合业内的研究力量和资源, 建立一个底栖生物学科合作与交流的平台, 推动海洋底栖生物学科的繁荣发展, 为国家海洋战略的深入实施提供重要的技术支撑与保障, 促进我国底栖生物学的发展。
在全国各涉海单位的推动及海洋科学工作者的积极努力下, 已基本掌握了我国海域和河口主要底栖动物类群的种类、分布、资源利用情况, 以及主要的生态学和生物多样性特点。但仍存在几个方面需要加强。
(1)加强分类学研究, 开展对尚未进行编目类群的分类研究, 尤其是对稀有类群和分类难度较大优势类群的分类研究, 海洋底栖生物的分类研究依然任重道远。当前基于形态学特征的分类鉴定仍然是国际间分类学研究的主流, 每年大量的新种、新属甚至新科被发现和描述。然而, 仅仅基于形态学特征已不能满足对目前分类研究的要求, 因此将分子生物学等新的技术手段引入到海洋底栖生物分类学和系统学研究已势在必行。
(2)之前研究大多涉及底栖动物多样性、次级生产力和群落结构及其变化的动态分析,但主要停留在对数据的观测及现象的描述,缺乏深入、系统的生态学分析和机理的探究,亟待展开深入的生态学和多样性研究, 全面理解海洋生态系统动力学的运行机制。
(3)需加强时间序列的数据资料积累和新技术手段的应用, 以及对国际前沿的跟踪, 拓展研究视角和研究领域, 增强底栖生物研究的广度和深度。另外, 现代研究应将微微型的细菌、微藻、微型动物、小型底栖动物及大型底栖动物组成的底栖食物网视为一个结构完整的复合体而进行研究。
(4)加强对尚未探知的生境, 尤其是深海(如海山、热液、冷泉、深渊等)底栖生物的研究, 将中国的底栖生物学研究从近岸向远洋、从浅海向深海拓展, 并使其在全球海洋底栖生物的生物多样性和地理分布研究中占有一席之地。
海洋底栖生物无论是在生物多样性还是生态功能方面, 均是整个海洋生态系统不可缺少的组成部分。加强对这一较为薄弱领域的研究, 研究其生物多样性构成、分布与变动及驱动变化的因子, 将有助于加深对海洋生物多样性整体的认识, 为海洋生态系统结构和功能及全球碳的源汇等重大问题的解析提供新的视角。
致谢 谨以此文献给即将成立的中国海洋与湖沼学会海洋底栖生物学分会。
鲍毅新, 葛宝明, 郑祥, 等. 2006. 温州湾天河滩涂大型底栖动物群落分布与季节变化. 动物学报, 52(1): 45-52
蔡立哲, 厉红梅, 邹朝中. 2000a. 深圳河口福田泥滩海洋线虫的种类组成及季节变化. 生物多样性, 8(4): 385-390
蔡立哲, 洪华生, 邹朝中, 等. 2001. 台湾海峡中北部海洋线虫群落的种类组成及分布. 台湾海峡, 20(2): 228-235
蔡立哲, 李复雪. 1998. 厦门潮间带泥滩和虾池小型底栖动物类群的丰度. 台湾海峡, 17(1): 91-95
蔡立哲, 洪华生, 邹朝中, 等. 2000b. 台湾海峡南部海洋线虫种类组成及其取食类型. 台湾海峡, 19(2): 212-217
杜永芬, 徐奎栋, 类彦立, 等. 2011. 青岛湾小型底栖生物周年数量分布与沉积环境. 生态学报, 31(2): 431-440
杜永芬, 徐奎栋, 孟昭翠, 等. 2010. 南海小型底栖动物生态学的初步研究. 海洋与湖沼, 41(2): 199-207
范士亮, 刘海滨, 张志南, 等. 2006. 青岛太平湾砂质潮间带小型底栖生物丰度和生物量的研究. 中国海洋大学学报, 36(S1): 98-104
范士亮, 王宗兴, 徐宗军, 等. 2011. 南黄海冬季小型底栖生物分布特征. 海洋环境科学, 30(2): 185-188
方少华, 吕小梅, 张跃平, 等. 2000. 台湾海峡小型底栖生物数量的量分布. 海洋学报, 22(6): 136-140
古丽亚诺娃 E Ф, 刘瑞玉, 斯卡拉脱 O A, 等. 1958.黄海潮间带生态学研究. 北京: 科学出版社
郭玉清, 叶兆弘. 2008. 虾池小型底栖动物类群组成和数量分布. 集美大学学报(自然科学版), 13(1): 18-23
郭玉清, 张志南, 慕芳红. 2002. 渤海小型底栖动物丰度的分布格局. 生态学报, 22(9): 1463-1469
郭玉清, 张志南, 慕芳红. 2003. 渤海自由生活海洋线虫多样性的研究. 海洋学报, 25(2): 106-113
郭玉清. 2008. 厦门凤林红树林湿地自由生活海洋线虫群落的研究. 海洋学报, 30(4): 147-153
韩洁, 张志南, 于子山. 2004. 渤海中、南部大型底栖动物的群落结构. 生态学报, 24(3): 531-537
华尔, 李佳, 董洁, 等. 2012. 砂质潮间带自由生活海洋线虫对缺氧的响应——微型受控生态系研究.生态学报, 32(13): 3975-3986
华尔, 张志南, 张艳. 2005. 长江口及邻近海域小型底栖生物丰度和生物量. 生态学报, 25(9): 2234-2242
黄勇. 2005. 南黄海小型底栖生物生态学和海洋线虫分类学研究. 青岛: 中国海洋大学博士学位论文
黄勇, 张志南. 2007. 南黄海自由生活线虫的分类学研究. 聊城大学学报(自然科学版), 20(2): 14-19
江锦祥, 黄立强, 孟凡, 等. 1985. 东海大陆架及其邻近海区底栖生物种类组成和分布特点. 台湾海峡, 4(1): 89-98
类彦立, 徐奎栋. 2011. 海洋微型底栖生物调查方法与操作规程. 海洋与湖沼, 42(1): 157-164
李宝泉, 李新正, 王洪法, 等. 2007. 长江口附近海域大型底栖动物群落特征. 动物学报, 53(1): 76-82
李荣冠, 江锦祥. 1989. 厦门西部海域大型底栖生物群落变化. 台湾海峡, 8(2): 144-149
李新正, 李宝泉, 王洪法, 等. 2006. 胶州湾潮间带大型底栖动物的群落生态. 动物学报, 52(3): 612-618
李新正, 王洪法, 张宝琳. 2005. 胶州湾大型底栖动物次级生产力初探. 海洋与湖沼, 36(6): 527-533
李新正. 2011. 我国海洋大型底栖生物多样性研究及展望: 以黄海为例. 生物多样性, 19(6): 676-684
林岿璇, 张志南, 王睿照. 2004. 东、黄海典型站位底栖动物粒径谱研究. 生态学报, 24(2): 241-245
刘录三, 孟伟, 田自强, 等. 2008. 长江口及毗邻海域大型底栖动物的空间分布与历史演变. 生态学报, 28(7): 3027-3034
刘瑞玉, 崔玉珩, 徐凤山, 等. 1986. 黄海、东海底栖生物的生态特点. 海洋科学集刊, (27): 153-173
刘瑞玉, 罗秉征, 崔玉珩, 等. 1987. 三峡工程对河口生物及渔业资源的影响//中国科学院三峡工程生态与环境科研项目领导小组. 长江三峡工程对生态与环境影响及其对策研究论文集. 北京: 科学出版社: 403-446
刘瑞玉, 徐凤山. 1963. 黄、东海底栖动物区系的特点.海洋与湖沼, 5(4): 306-321
刘瑞玉, 赵一阳, 翁学传, 等. 1996. 中韩黄海海洋学联合研究新进展. 海洋与湖沼, 27(6): 657-662
刘瑞玉. 1990. 中国关于黄海海洋生态学的研究. 黄海研究, 3: 51-79
刘瑞玉. 1992. 胶州湾生态学和生物资源. 北京: 科学出版社
刘晓收. 2005. 南黄海鳀鱼产卵场小型底栖动物生态学研究. 青岛: 中国海洋大学硕士学位论文
陆鼎恒. 1934. 动吻类一新种在烟台之发现//国立北平研究院动物研究所丛刊. 北平: 国立北平研究院出版部: 1-9
孟昭翠, 徐奎栋. 2013. 长江口及东海春季底栖硅藻、原生动物和小型底栖生物的生态特点. 生态学报, 33(21): 6813-6824
慕芳红, 张志南, 郭玉清. 2001a. 渤海底栖桡足类群落结构的研究. 海洋学报, 23(6): 120-127
慕芳红, 张志南, 郭玉清. 2001b. 渤海小型底栖生物的丰度和生物量. 青岛海洋大学学报, 31(6): 897-905
钱国珍, 倪铮, 曲云龙. 1992. 用海洋线虫监测潮间带有机质污染的调查研究. 海洋湖沼通报, (3): 48-55
史本泽, 于婷婷, 徐奎栋. 2015. 长江口及东海夏季小型底栖动物丰度和生物量变化. 生态学报, 35(9): 3093-3103
寿鹿, 廖一波, 徐晓群, 等. 2009. 福清核电站邻近潮间带大型底栖生物数量分布与群落结构. 海洋学研究, 27(2): 42-50
孙道元, 刘银城. 1991. 渤海底栖动物种类组成和数量分布. 黄渤海海洋, 9(1): 42-50
孙道元, 徐凤山, 崔玉衍, 等. 1992. 长江口区枯、丰水期后底栖生物分布特点. 海洋科学集刊, (33): 217-235
唐玲, 李恒翔, 严岩. 2012a. 大亚湾春季小型底栖动物初步研究. 海洋环境科学, 31(3): 405-409
唐玲, 张洪波, 李恒翔, 等. 2012b. 大亚湾秋季小型底栖生物初步研究. 热带海洋学报, 31(4): 104-111
唐质灿, 徐凤山. 1978. 东海大陆架区底栖生物数量分布和群落的初步分析//中国科学院海洋研究所.东海大陆架论文集. 青岛: 中国科学院海洋研究所: 156-164
王家栋, 类彦立, 徐奎栋, 等. 2011. 黄海冷水团及周边海域夏初小型底栖动物现存量及空间分布研究. 海洋与湖沼, 42(3): 359-366
王彦国, 王春光, 陈小银, 等. 2011. 北部湾海域小型底栖动物丰度和生物量. 生态科学, 30(4): 375-382
吴绍渊, 慕芳红. 2009. 山东南部沿海冬季小型底栖生物的初步研究. 海洋与湖沼, 40(6): 682-691
吴秀芹, 徐奎栋, 于子山, 等. 2010. 2008年浒苔大暴发末期黄海小型底栖动物现存量及空间分布. 应用生态学报, 21(8): 2140-2147
吴耀泉, 张宝林. 1990. 渤海经济无脊椎动物生态特点的研究. 海洋科学, (2): 48-52
徐奎栋. 2011. 海洋微型底栖生物的多样性与地理分布. 生物多样性, 19(6): 661-675
于海燕, 李新正, 李宝泉, 等. 2006. 胶州湾大型底栖动物生物多样性现状. 生态学报, 26(2): 416-422
于婷婷, 徐奎栋. 2013. 长江口及邻近海域秋冬季小型底栖动物类群组成与分布. 生态学报, 33(15): 4556-4566
于子山, 张志南, 韩洁. 2001. 渤海大型底栖动物次级生产力的初步研究. 青岛海洋大学学报, 31(6): 867-871
张玺. 1935. 胶州湾海产动物采集团第一期采集报告//国立北平研究院动物学研究所中文报告汇刊. 北平: 国立北平研究院出版部: 1-95
张玺, 马绣同. 1936. 胶州湾海产动物采集团第二期及第三期采集报告//国立北平研究院动物学研究所中文报告汇刊. 北平: 国立北平研究院出版部: 1-176
张玺, 马绣同. 1949. 胶州湾海产动物采集团第四期采集报告//国立北平研究院动物学研究所中文报告汇刊. 北平: 国立北平研究院出版部: 1-113
张玺, 相里矩. 1936. 胶州湾及其附近海产食用软体动物之研究//国立北平研究院动物学研究所中文报告汇刊. 北平: 国立北平研究院出版部: 1-94
张艳. 2009. 胶州湾典型站位小型底栖生物丰度和生物量的季节变化研究. 中国农学通报, 25(17): 296-301
张艳, 张志南, 邓可, 等. 2005. 青岛岩礁附植小型底栖动物 ATP含量的研究. 中国海洋大学学报, 35(5): 799-806
张志南. 2000. 水层-底栖耦合生态动力学研究的某些进展. 青岛海洋大学学报, 30(1): 115-122
张志南, 党宏月, 于子山. 1993. 青岛湾有机质污染带小型底栖生物群落的研究. 青岛海洋大学学报, 23(1): 83-91
张志南, 李永贵, 图立红, 等. 1989. 黄河口水下三角洲及其邻近水域小型底栖动物的初步研究. 海洋与湖沼, 20(3): 197-208
张志南, 林岿旋, 周红, 等. 2004. 东、黄海春秋季小型底栖生物丰度和生物量研究. 生态学报, 24(5): 997-1005
张志南, 周红, 于子山. 2001. 胶州湾小型底栖生物的丰度和生物量. 海洋与湖沼, 32(2): 139-147
张志南, 周红. 2004. 国际小型底栖生物研究的某些进展. 中国海洋大学学报, 34(5): 799-806
章飞军, 童春富, 张衡, 等. 2007. 长江口潮下带春季大型底栖动物的群落结构. 动物学研究, 28(1): 47-52
赵永强, 曾江宁, 高爱根, 等. 2009. 椒江口滩涂大型底栖动物群落格局与多样性. 生物多样性, 17(3): 303-309
周百灵, 孟昭翠, 赵峰, 等. 2015. 长江口外海域微型和小型底栖生物群落结构和时空变化. 生态学报, 35(15): 5050-5063
Boesch D F, 唐质灿, 徐凤山, 等. 1986. 东湖大陆架沉积物内大型底栖生物和生源结构. 海洋科学集刊, (27): 187-197
Cai L Z, Fu S J, Yang J. et al. 2012. Distribution of meiofaunal abundance in relation to environmental factors in Beibu Gulf, South China Sea. Acta Oceanologica Sinica, 31(6): 92-103
Coull B C. 1999. Role of meiofauna in estuarine softbottom habitats. Australian Journal of Ecology, 24(4): 327-343
Du Y F, Xu K D, Lei Y L. 2009. Simultaneous enumeration of diatom, protozoa and meiobenthos from marine sediments using Ludox-QPS method. Chinese Journal of Oceanology and Limnology, 27(4): 775-783
Ellingsen K E, Gray J S. 2002. Spatial patterns of benthic diversity: is there a latitudinal gradient along the Norwegian continental shelf? Journal of Animal Ecology, 71(3): 373-389
Fleeger J W, 唐质灿, Higgins R P. 1986. 长江口及邻近海域小型底栖桡足类和动吻类生态的初步研究. 海洋科学集刊, (27): 199-208
Giere O. 2009. Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments. 2nd ed. Berlin Heidelberg: Springer-Verlag
Gray J S, Elliott M. 2009. Ecology of Marine Sediments: from Science to Management. Oxford: Oxford University Press
Guo Y Q, Somerfield P J, Warwick R M, et al. 2001a. Large-scale patterns in the community structure and biodiversity of freeliving nematodes in the Bohai Sea, China. Journal of the Marine Biological Association of the United Kingdom, 81(5): 755-763
Guo Y Q, Warwick R M, Zhang Z N, et al. 2002. Freeliving marine nematodes as a pollution indicator of the Bohai Sea. Journal of Environmental Sciences, 14(4): 558-562
Guo Y Q, Zhang Z N, Mu F H. 2001b. Biomass of meiobenthic in the Bohai Sea, China. Acta Oceanologica Sinica, 20(2): 435-442
Higgins R P, Thiel H. 1988. Introduction to the Study of Meiofauna. Washington D C: Smithsonian Institution Press
Hua E, Zhang Z N, Zhang Y. 2006. Meiofauna distributions at the oxygen minimum zone in Changjiang (Yangtze) River Estuary waters. Acta Oceanologica Sinica, 25(5): 120-134
Hua E, Zhang Z N, Zhang Y. 2009. Environmental factors affecting nematode community structure in the Changjiang Estuary and its adjacent waters. Journal of the Marine Biological Association of the United Kingdom, 89(1): 109-117
Liu L S, Zheng B H. 2010. Secondary production of macrobenthos in the Yangtze River Estuary ant its adjacent waters. Chinese Journal of Applied & Environmental Biology, 16(5): 667-671
Liu R Y, Cui Y H, Xu F S, et al. 1983. Ecology of macrobenthos of the East China Sea and adjacent waters//Proceedings of International Symposium on the Sedimentation on the Continental Shelf, with Special Reference to the East China Sea. Beijing: China Ocean Press: 795-818
Liu X S, Cheung S G, Shin P K S. 2009. Meiofauna with special reference to nematodes in trawling ground of subtropical Hong Kong. Marine Pollution Bulletin, 58(4): 607-615
Liu X S, Xu W Z, Cheung S G, et al. 2008. Subtropical meiobenthic nematode communities in Victoria Harbour, Hong Kong. Marine Pollution Bulletin, 56(8): 1491-1497
Liu X S, Xu W Z, Cheung S G, et al. 2011a. Marine meiobenthic and nematode community structure in Victoria Harbour, Hong Kong upon recovery from sewage pollution. Marine Pollution Bulletin, 63(5-12): 318-325
Liu X S, Xu W Z, Cheung S G, et al. 2011b. Response of meiofaunal community with special reference to nematodes upon deployment of artificial reefs and cessation of bottom trawling in subtropical waters, Hong Kong. Marine Pollution Bulletin, 63(5-12): 376-384
McIntyre A D. 1969. Ecology of marine meiobenthos. Biological Reviews, 44(2): 245-288
Meng W, Liu L S, Zheng B H, et al. 2007. Macrobenthic community structure in the Changjiang Estuary and its adjacent waters in summer. Acta Oceanologica Sinica, 26(6): 62-71
Meng Z C, Xu K D, Dai R H, et al. 2012. Ciliate community structure, diversity and trophic role in offshore sediments from the Yellow Sea. European Journal of Protistology, 48(1): 73-84
Meng Z C, Xu K D, Lei Y L. 2011. Community composition, distribution, and contribution of microbenthos in offshore sediments from the Yellow Sea. Continental Shelf Research, 31(14): 1437-1446
Snelgrove P V R. 1998. The biodiversity of macrofaunal organisms in marine sediments. Biodiversity & Conservation, 7(9): 1123-1132
Xu K D, Du Y F, Lei Y L, et al. 2010. A practical method of Ludox density gradient centrifugation combined with protargol staining for extracting and estimating ciliates in marine sediments. European Journal of Protistology, 46(4): 263-270
Zhang J L, Xiao N, Zhang S P, et al. 2016a. A comparative study on the macrobenthic community over a half century in the Yellow Sea, China. Journal of Oceanography, 72(2): 189-205
Zhang J L, Zhang S P, Zhang S Q, et al. 2016b. What has happened to the benthic mollusks of the Yellow Sea in the near half century? Comparison on molluscan biodiversity between 1959 and 2007. Continental Shelf Research, 113: 21-29
Zhang Q T, Wang X H, Hu G K. 2010. Evaluation of estimation methods for meiofaunal biomass from a meiofaunal survey in Bohai Bay. Chinese Journal of Oceanology and Limnology, 28(1): 82-87
Zhou H, Zhang Z N, Liu X S, et al. 2007. Changes in the shelf macrobenthic community over large temporal and spatial scales in the Bohai Sea, China. Journal of Marine Systems, 67(3-4): 312-321
Zhou H, Zhang Z N. 2008. Nematode assemblages from submarine caves in Hong Kong. Journal of Natural History, 42(9-12): 781-795
Zhou H. 2001. Effects of leaf litter addition on meiofaunal colonization of azoic sediments in a subtropical mangrove in Hong Kong. Journal of Experimental Marine Biology and Ecology, 256(1): 99-121
Progress and Prospect in Marine Benthology in China
ZHANG Jun-Long, SHI Ben-Ze, ZHAO Feng, XU Kui-Dong*
(Department of Marine Organism Taxonomy and Phylogeny, Institute of oceanology, Chinese Academy of Sciences, Qingdao 266071, China) *Corresponding author, E-mail: kxu@qdio.ac.cn
Marine benthos are important components in marine ecosystem, serving vital functions in energy flow and material cycling of marine bottom environment. During the past decades, studies on Marine Benthology in China have achieved major progress, but there are still some weaknesses in many research fields. In general, research activities on Marine Benthology in China are fragmented and deficient. It is in urgent need of establishing a specialized society to facilitate cooperation, exchange ideas, and integrate and share research resources. Based on the analysis of current status, we put forward proposals for future study and elaborate the function of the society in future development of Marine Benthology.
benthos; Marine Benthology Branch of Chinese Society for Oceanology and Limnology; ecology; taxonomy; biodiversity
Q958.8
10.12036/hykxjk20160725005
* 资助项目: 中国科学院战略性先导科技专项(A类)项目(XDA11030201); 国家自然科学基金项目(41476144)。张均龙, 男, 副研究员, E-mail: zhangjl@qdio.ac.cn
① 通讯作者: 徐奎栋, 男, 研究员, 主要从事海洋生物分类与多样性研究, E-mail: kxu@qdio.ac.cn
2016-07-25, 收修改稿日期: 2016-07-31
