海洋浮游微食物网结构及其影响因素*
2016-03-15张武昌
张武昌 陈 雪 赵 苑 赵 丽 肖 天
(1. 中国科学院海洋研究所 海洋生态与环境科学重点实验室, 青岛 266071; 2. 青岛海洋科学与技术国家实验室 海洋生态与环境科学功能实验室, 青岛 266071; 3. 中国科学院大学, 北京 100049)
海洋浮游微食物网结构及其影响因素*
张武昌1,2①陈 雪1,2,3赵 苑1,2赵 丽1,2肖 天1,2
(1. 中国科学院海洋研究所 海洋生态与环境科学重点实验室, 青岛 266071; 2. 青岛海洋科学与技术国家实验室 海洋生态与环境科学功能实验室, 青岛 266071; 3. 中国科学院大学, 北京 100049)
微食物环是海洋生态系统中重要的物质和能量过程, 是传统食物链的有效补充。微食物环研究是当前海洋生态学研究的热点之一, 但对其结构的系统研究较少,海洋微食物网结构在2000年才被Garrison提出。尽管微食物网各个类群的丰度在不同海洋环境中存在相对变化, 但是这些变化都处于一定的范围之内, 其丰度结构约为纤毛虫10cell/mL、鞭毛虫103cell/mL、微微型真核浮游生物104cell/mL、蓝细菌104~ 105cell/mL、异养细菌 106cell/mL、病毒 107particle/mL。海洋浮游食物链中捕食者和饵料生物粒径的最佳比值为 10︰1, 实际研究中该比值会略低, 如纤毛虫与其饵料的粒径比值为8︰1, 鞭毛虫为3︰1。微微型(pico-)和微型(nano-)浮游植物的丰度比(pico︰nano)是研究微食物网结构的指数之一, 该指数具有不受研究尺度影响的优点, 可用于研究区域性和全球性微食物网结构。近年来, 学者们从多角度对海洋微食物网的结构开展了研究, 针对不同海区微食物网各类群丰度、生物量时间和空间变化的研究有很多报道, 微食物网的结构受空间、季节、摄食、营养盐等多种因素影响。在对不同空间微食物网的研究中, 众多学者往往研究不同物理性质的水团中各类群生物丰度的不同,以此来表征微食物网结构的不同; 同一海区微食物网结构的季节变化也是使用各个类群丰度和生物量的变化来表示, 该变化主要受水文环境因素影响。摄食者对微食物网各类生物的影响通过 3种途径: ①中型浮游动物摄食; ②中型浮游动物摄食微型浮游动物, 通过营养级级联效应影响低营养级生物; ③中型浮游动物通过释放溶解有机物、营养盐影响细菌和低营养级生物。浮游植物通过产生化感物质和溶解有机物影响微食物网结构, 而营养盐的浓度及变化则可以对微食物网产生直接或间接影响。
微食物网; 结构; 微微型浮游生物; 鞭毛虫; 纤毛虫
1983年, 微食物环(microbial loop)概念首次被提出(Azam et al., 1983)。海洋是地球上三大有机碳库之一, 这些有机碳大部分以溶解有机碳(dissolved organic carbon, DOC)的形式存在, 海洋异养细菌(heterotrophic bacteria, HB)通过“二次生产”(Fuhrman and Azam, 1980)将DOC转化为自身物质以生长和繁殖, 随着原生生物(如鞭毛虫、纤毛虫等)摄食异养细菌,细菌回收的碳被再次传递返回经典食物链,从而构成微食物环(Azam et al., 1983; Pomeroy and Wiebe, 1988)。
海洋微食物网(microbial food web)是微食物环概念的延伸。微食物环概念最初仅用于解释以细菌为基础的摄食关系, 科学家发现海洋中存在着大量与细菌粒径相近的自养生物, 包括聚球藻属蓝细菌(Synechococcus) (Waterbury et al., 1979; Murphy and Haugen, 1985)和原绿球藻属蓝细菌(Prochlorococcus)(Chisholm et al., 1988, 1992), 以及海洋微微型真核浮游生物(picoeukaryotes)(Fenchel 1982; Johnson and Sieburth, 1982)。这些自养生物通过光合作用进行自身生长, 随后同样被原生动物和微型浮游生物所摄食, 从而进入经典食物链。Sherr等(1988)对微食物环两条路径进行了总结, 认为这些微小生物之间的营养关系不再是一条线, 而是交织成一个网状, 因此提出海洋微食物网概念。海洋浮游病毒作为分解者, 裂解藻类和细菌向水中释放溶解有机物(dissolved organic matter, DOM), 而这部分DOM又再被细菌利用, 形成了微食物网的病毒回路(viral shunt)(Wilhelm and Suttle, 1999), 这方面的研究进一步完善了微食物网的概念。
长久以来, 人们就认识到食物网结构可以对海洋生态系统产生重要影响, 如鱼产量(fish yield)(Ryther, 1969)、海洋碳通量(carbon flux)(Azam et al., 1983; Michaels and Silver, 1988)、营养盐再生(nutrient regeneration) (Caron, 1991)等。随着对海洋微型生物研究的逐渐开展, 科学家也开始重视海洋微食物网的结构(microbial food web structure)及其主要影响因素的研究, 以期进一步了解和预测海洋生态系统变化和生物地球化学循环过程。
海洋微食物网结构这一概念最早是由Garrison在 2000年提出的, 但是他并没有给出结构的定义, 而是测定微食物网不同生物类群的丰度和生物量, 通过聚类分析的方法,判断不同调查时间的微食物网是否相同(Garrison et al., 2000)。近年来, 学者们从不同角度对海洋微食物网的结构进行了深入研究。本文将从以下几个方面介绍微食物网结构:微食物网的丰度和生物量结构、微食物网的粒级结构和微食物网结构的变化和影响因素。
1 微食物网的丰度和生物量结构
1.1 各类群的丰度结构
尽管微食物网各个类群的丰度在不同海洋环境中有相对变化, 但是这些变化都处于一定的范围之内。一般说来, 每毫升海水中含有浮游病毒107个、细菌106个、蓝细菌104~105个、真核藻类104个( Azam and Malfatti, 2007),海洋浮游鞭毛虫的普遍丰度约为每毫升103个(Fenchel, 2008), 近岸海区的纤毛虫丰度约为每毫升 10个(Leakey et al., 1992)。Miki和Jacqnet(2008)指出, 在典型的海水中, 异养鞭毛虫、细菌、病毒的丰度比例为1︰103︰104。
在微食物网各类群中, 异养鞭毛虫(heterotrophic nanoflagellates, HNF)与细菌丰度的关系被研究得最多。有部分研究表明, 异养鞭毛虫与细菌丰度之间并没有良好的相关性(McManus and Fuhrman, 1990; Van Duyl et al., 1990; Weinbauer and Peduzzi, 1995), 另外一些研究则发现了二者之间的相关关系。Sanders等(1992)集合了之前多篇文献的数据,研究了在各种水生环境(包括海洋、河口以及淡水)中异养鞭毛虫和细菌丰度的数量关系,结果表明, 二者呈显著正相关(log HNFabundance=0.90×log bacteria abundance-2.4, R2=0.5, P<0.001, n=600) [图1(a)], 异养鞭毛虫和细菌丰度的数量级比例大约为 1︰103。Gasol和Vaque(1993)采用同样的方法, 得到了不同水生环境中异养鞭毛虫和细菌丰度的数量关系(log HNF abundance=0.79×log bacteria abundance-1.67, R2=0.31, P<0.001, n=41) [图1(b)], 这个结果整合了季节变化, 因此是一个平均值(mean realized abundance, MRA)。以上结果虽然表明异养鞭毛虫和细菌丰度之间存在显著的相关性, 但是相关关系较弱。在上述研究的基础上, Gasol(1994)提出了最大可达到丰度(maximum attainable abundance, MAA)线的概念, 即对于一个给定的细菌丰度, 鞭毛虫所能达到的最大丰度。基于收集的数据, 这个最大值边界为log HNFmax=-2.47+1.07×log HB,但是在自然海区中测得的数据很难达到MAA线。此外, Gasol(1994)还提出用一个定性模型来分析上行控制和下行控制对鞭毛虫丰度的影响(图2), 即当鞭毛虫和纤毛虫的比例高于MRA时, 上行控制为主要因素, 反之下行控制为主要因素。
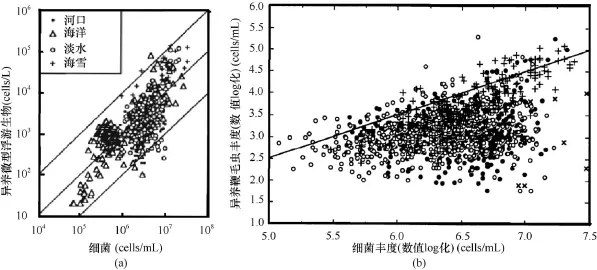
图1 各浮游生态系统中细菌和异养鞭毛虫(HNF)丰度的数量关系Fig.1 Relationship between abundance of bacterial and heterotrophic nanoflagellates (HNF) from a variety of planktonic ecosystems (a) 各浮游生态系统中细菌和异养微型浮游生物丰富的数量关系(Sanders et al., 1992); (b) 细菌和异养鞭毛虫丰度的数量关系(Gasol and Vaque, 1993) (a) 中黑色实线自上到下分别代表细菌丰度︰异养鞭毛虫=102︰1, 103︰1, 104︰1; (b) 中黑色实线表示异养鞭毛虫丰度的理论最大值, 估算基于鞭毛虫对细菌的摄食率为14cells/L和细菌每天分裂一次的假设
目前, 关于微食物网中其他生物类群丰度关系的报道还非常少, Chen等(2016)报道了我国荣成桑沟湾微食物网中 5个生物类群(聚球藻蓝细菌、微微型真核生物、异养鞭毛虫、自养鞭毛虫和纤毛虫)与异养细菌丰度的关系,在每个丰度关系中分别给出了MAA线, 并类推出了最小可达到丰度(minimum attainable abundance, MAA)线(图3)。
1.2 各类群相对生物量的变化

图2 上行控制(bottom-up)和下行控制(top-down)影响异养鞭毛虫(HNF)丰度(数值log化)的定性模型(Gasol, 1994)Fig.2 Framework for the analysis of the mechanisms of control (bottom-up and top-down) of the abundance of heterotrophic nanoflagellates (HNF) with the MAA and MRA lines, Note that the scale is logarithmic (Gasol, 1994)
虽然目前对微食物网各类群变化的研究较多, 但是对于相对生物量变化的研究非常少。对于如何判断不同海区和时间的微食物网是否相同, Garrison等(2000)使用聚类分析的方法研究得出, 阿拉伯海不同季风季节微食物网结构存在差异, 但仅用聚类分析的方法无法指出具体差异是什么。Chen等(2016)提出了归一化的方法来寻找具体差异, 即将异养细菌的生物量归为1, 其他类群生物量等比例变化, 得出各类群的相对生物量。用这种方法计算的相对生物量可以反映微食物网的生物量结构, 使得不同海区和不同季节的微食物网之间可以进行比较。Chen等(2016)用这种方法研究了桑沟湾海区微食物网的生物量结构,通过聚类分析方法表明, 桑沟湾微食物网结构存在季节变化和湾内外的地域性差异, 进而使用归一化的方法表明, 桑沟湾微食物网结构的季节变化主要源于聚球藻蓝细菌、微微型真核生物和鞭毛虫的相对生物量在夏秋季升高, 而纤毛虫的相对生物量在冬季升高;湾内、外微食物网结构的差异主要是由聚球藻蓝细菌、微微型真核生物和自养鞭毛虫引起的。
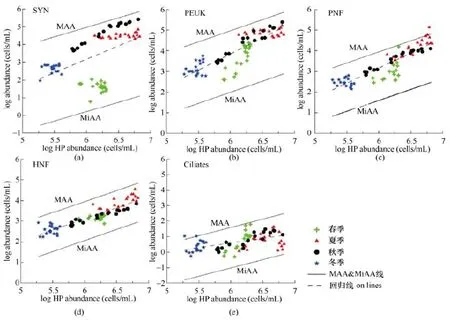
图3 异养原核生物与其他生物类群丰度(数值log化)的数量关系(Chen et al., 2016)Fig.3 Scatter graphs between HP abundance (cells/ml) and abundance (cells/ml or ind./ml) of the other five biological groups (SYN, PEUK, PNF, HNF, ciliates) with MAA and MiAA lines, Note that the scale is logarithmic (Chen et al., 2016) SYN(Synechococcus): 聚球蓝细菌; PEUK(picoeukaryotes): 微微型真核生物; PNF(pigmented nanoflagellates): 混合营养鞭毛虫; HP(heterotrophic prokaryotes): 异养原核生物; HNF(heterotrophic nanoflagellates): 异养鞭毛虫; ciliates: 纤毛虫。MAA(maximum attainable abundance): 最大可达到丰度; MiAA(minimum attainable abundance): 最小可达到丰度
2 微食物网的粒级结构
2.1 微食物网营养级的粒级结构
Azam等(1983)根据Sheldon等(1972)的海洋浮游生物的粒径模型, 建议海洋浮游食物链中捕食者和饵料生物粒径的最佳比值为10︰1,体积的最佳比值为 103︰1, 营养级间碳的输送效率为10%(Sheldon et al., 1972; Azam et al., 1983)。随着研究的深入, 多位学者发现, 在微食物网中摄食者和饵料生物粒级比值可能更小(Goldman and Caron, 1985; Hansen et al., 1994)。Goldman和 Caron(1985)用实验证明,异养鞭毛虫(Paraphysomonas imperforata)的饵料粒级变化较大, 以细菌为食时粒径比例为 7, 以浮游植物为饵料时, 粒径比例则降低至2。Hansen等(1994)总结了摄食试验中饵料和摄食者的粒径, 给出甲藻与其饵料的最佳粒径比值为 1︰1, 其他鞭毛虫(不含甲藻)为3︰1, 纤毛虫为8︰1(Hansen et al., 1994)。在后来的研究中, 有学者建议鞭毛虫与其饵料的粒径比值为3︰1(Guillou et al., 2001)。
海洋浮游鞭毛虫的主要粒径范围为 1~ 20μm, 按照 3︰1的鞭毛虫与饵料粒径比例,在自然海区的鞭毛虫应该存在多个营养级,这一现象在自然海区海水多级过滤培养中得到证实(Rassoulzadegan and Sheldon, 1986; Wikner and Hagström, 1988; Chen et al., 2009),不同学者对微型浮游动物的营养级划分略有不同, 如 Wikner和 Hagström(1988)将鞭毛虫分为1~5μm、5~8μm、8~10μm和>10μm的营养级, Chen等(2009)则把微型浮游动物划分为5个营养级<2μm、2~5μm、5~10μm、10~60μm和60~200μm。
因为微型浮游动物存在多个营养级, 所以微食物网内部也会存在营养级级联摄食。Calbet等(2001)在研究细菌与鞭毛虫之间的营养级联作用时, 发现如果 2~3μm粒级的异养鞭毛虫被去除, 小于 2μm粒级的异养鞭毛虫的丰度则会增加, 进而导致细菌丰度降低, 在细菌有较高生物量(8~14μgC/L)时, 这种营养级联现象会更加显著。分粒级稀释培养实验的结果也反映了营养级级联现象, 去除>10μm和>3μm的微型浮游动物后, 微微型浮游生物(picoplankton, 粒径≤2μm)的内禀增长率和被摄食率都有所增加, 被摄食率的增加可能是因为微型浮游动物中存在营养级联作用, 较大个体的微型浮游动物被过滤去除后, 较小个体的微型浮游动物因为失去了摄食者而生长, 进而增加了对微微型浮游生物的摄食率(Reckermann and Veldhuis, 1997)。Chen和Liu (2010)使用分粒级稀释培养也发现, 去除粒径>20μm 以上的浮游动物后, 微微型浮游生物的被摄食率增加。
2.2 微微型浮游生物和微型浮游生物的比例
微微型浮游生物和微型浮游生物(nanoplankton, 2~20μm 的浮游生物)是根据粒径来进行划分的(Sieburth et al., 1978), 微微型浮游植物(picophytoplankton)和微型浮游植物(nanophytoplankton)的丰度比(pico︰nano)是研究微食物网结构的指数之一, 该指数具有不受研究尺度影响的优点, 可用于研究区域性和全球性微食物网结构(Li et al., 2013; López-Urrutia, 2013)。海洋环境中pico︰nano比值范围为 1~1000, 在海洋上层水体的比值高于底层, 在低纬度海区高于高纬度海区。pico︰nano比值常应用于“空间代替时间”(spacefor-time substitution studies)的研究方法中,可以用来预测海洋微食物网的长期变化(López-Urrutia, 2013)。
3 微食物网结构的变化和影响因素
3.1 空间和季节的影响
空间和季节对微食物网的结构及变化有重要影响, 这些影响更多的是通过温度、盐度等环境因子对微食物网各不同类群产生影响而实现的。
在对不同空间微食物网的研究中, 人们往往研究不同物理性质的水团中各类群生物丰度的不同, 以此来表征微食物网结构的不同。Jacquet等(2010)研究了地中海西南部Alboran海的微微型浮游生物的丰度及空间分布, 他们将研究海区分为3个水团: 北部的地中海水团、南部的大西洋水团, 以及二者之间的锋面。调查发现, 低温高盐的地中海水团原绿球藻蓝细菌丰度较高, 高温低盐的大西洋水团聚球藻蓝细菌丰度较高, 在锋面海区这两类蓝细菌的丰度都较高, 此外微微型真核浮游生物和异养细菌在大西洋水团和锋面区域丰度较高(Jacquet et al., 2010)。Christaki等(2011)研究了地中海中部3个涡内外微食物网的差异, 发现在涡内部异养生物的丰度低于外部。在比斯开湾的Urdaibai河口, 受潮汐影响较大的站位其细菌、异养鞭毛虫和纤毛虫比受潮汐影响较小的站位生物量大(Iriarte et al., 2003)。
同一海区微食物网结构的季节变化也使用各个类群丰度和生物量的变化来表示。微食物网中微微型浮游生物的季节变化主要受水文环境因素影响, 在温带海域其丰度的季节性变动通常为夏季和秋季最大、冬季最小(Agawin et al., 1998; Li, 1998; Liu et al., 2002)。在比斯开湾, 聚球藻和原绿球藻蓝细菌丰度在夏、秋季占优势, 其他季节则是微微型真核浮游生物占优势(Calvo-Díaz and Morán, 2006)。在我国南海, 原绿球藻丰度在夏季最高, 并且在各个季节均比聚球藻和微微型真核浮游生物丰度高 1~2个数量级, 聚球藻和微微型真核浮游生物的丰度虽然较低, 但变化幅度很大, 在冬季至早春达到丰度最大值,可能与研究海域冬季混合层深度的变化有关(Liu et al., 2007)。原生动物类群(鞭毛虫、纤毛虫)丰度和生物量的变化受到自养生物类群丰度和生物量的影响。Dupuy等(2007)研究了大西洋近岸浮游生物群落结构, 发现原生动物类群的生物量在夏季较高、冬季最低; 但不论什么季节, 鞭毛虫生物量所占比例都很高(53%~66%)。此外, 季节变化会影响微食物网的粒径级组成。冬、春季粒级较大(micro-级)的类群占优势, 夏秋季则以粒级较小的类群(pico-和nano-级)为主(Savenkoff et al., 2000; Shinada et al., 2005)。
3.2 摄食者对微食物网结构的影响
摄食者通过 3种途径对微食物网各类生物产生影响: ①中型浮游动物摄食; ②中型浮游动物摄食微型浮游动物, 通过营养级级联效应影响低营养级生物; ③中型浮游动物通过释放溶解有机物、营养盐影响细菌和低营养级生物。摄食者对微食物网结构的影响研究中, 研究最多的是桡足类和水母。
3.2.1 桡足类对微食物网结构的影响
研究桡足类对微食物网的影响通常采用自然海水添加桡足类实验的方法。桡足类摄食的饵料最小粒级是 5μm, 因此桡足类能摄食微食物网中的纤毛虫、鞭毛虫, 其中纤毛虫是桡足类最优先摄食的饵料, 几乎所有的摄食培养实验中都显示桡足类对纤毛虫有很高的清滤率(张武昌等, 2014), 但是其他类群对添加桡足类的反应(增加或减少)并没有一致的结果, 添加中型浮游动物对微食物网底部的生物影响很小(Calbet and Landry, 1999)。
按照微食物环细菌-异养鞭毛虫-纤毛虫的营养结构, 从理论上来看, 添加桡足类既然导致纤毛虫减少, 就应该会引起异养鞭毛虫增加, 但是在有的实验中观测到异养鞭毛虫增加, 有的实验中异养鞭毛虫反而降低, 说明细菌-异养鞭毛虫-纤毛虫的营养级级联效应较弱。对以上现象的解释分为3个方面: ①异养鞭毛虫的摄食者(桡足类、纤毛虫和食鞭毛虫的鞭毛虫)之间互相摄食使得理论上的线性摄食关系不成立; ②异养鞭毛虫的控制因子主要是上行控制(即受饵料控制), 被摄食不是主要的限制因子; ③虽然在实验过程中纤毛虫丰度降低了, 但是由此释放的对异养鞭毛虫的摄食压力造成的后果在短时间内来不及显现(Samuelsson and Andersson, 2003; Sipura et al., 2003)。
3.2.2 水母对微食物网结构的影响
近年来, 水母暴发(Jellyfish Blooms)现象在世界近岸区域频发, 其范围和严重程度有可能继续增加, 因此水母对微食物网的影响得到科学家的重视。水母添加实验证明, 添加水母后, 水母释放的胶体态和溶解态有机物质会增加细菌的丰度, 并改变其优势类群(Condon et al., 2011)。研究发现, 含虫黄藻的水母(Catostylus mosaicus)和不含虫黄藻的水母(Phyllorhiza punctata)都能摄食中型浮游动物, 通过营养级级联作用导致异养甲藻(Protoperidinium sp.)的增加(West et al., 2009)。在一个中型围隔(300 L)进行水母(Mnemiopsis leidyi)添加培养的实验中, 通过 168h的培养,发现这种水母降低了中型浮游动物的生物量,甲藻的生物量增加, 小型浮游植物和纤毛虫丰度降低, 细菌丰度的变化较小, 这个结果说明水母对中型浮游动物的影响比较明显, 而对细菌等低营养级生物的影响不明显(Dinasquet et al., 2012)。
3.2.3 贝类对微食物网结构的影响
双壳类养殖生物对微食物网的影响体现在两个方面: 一是对微食物网各生物类群的摄食, 二是释放营养盐影响微食物网各生物类群的生长。
贝类对微食物网各生物类群的不同截留效率(retention efficiency)会影响微食物网的结构(Dupuy et al., 2000a)。目前, 对贝类滤食微食物网组分的研究多集中在实验室内或海上原位实验, 对自然海区的研究还未见报道。已有的贝类添加实验证明, 双壳类(包括贻贝、牡蛎和扇贝等)可以滤食或截留(retain)微微型浮游生物(包括异养细菌、微微型自养生物、微微型真核生物等)、鞭毛虫和纤毛虫(Kreeger and Newell, 1996; Le Gall et al., 1997; Dupuy et al., 1999, 2000a, 2000b; Fournier et al., 2012),但是对不同生物类群的截留效率不同, 并且截留效率随生物粒径级的增加而增加。Kryger和Riisgård(1988)通过用6种双壳贝类摄食不同粒径级(2~9μm)饵料(细菌和单胞藻)的实验发现, 有大型前侧纤毛的贝类可以完全截留4μm以上的饵料, 对于 2μm的饵料截留效率下降至 35%~70%; 有小型前侧纤毛或者没有的种类可以完全保留 5~6μm 的饵料, 对于2μm 的饵料, 截留效率分别下降为 50%和15%。Dupuy等(2000b)的摄食实验表明, 牡蛎可以摄食>5μm的鞭毛虫、微型浮游植物、甲藻、纤毛虫和大型浮游动物, 对<5μm的鞭毛虫和微微型真核生物的截留效率为 45%和2%。虽然微型藻类(硅藻和甲藻)、鞭毛虫和纤毛虫均能被贝类有效滤食, 但从碳贡献率分析,异养或混合营养的原生动物(hetero/mixotrophic protists)的贡献率较高(主要为鞭毛虫、其次为纤毛虫), 硅藻和甲藻等微藻的碳贡献率很低,因此异养或混合营养的原生动物可以将微食物环中的细菌生产力(bacterioplanktonic production)向更高营养级(双壳类等)传递(Dupuy et al., 2000a)。
除了摄食的作用, 贝类也可以通过释放无机营养盐影响微食物网。Lu等(2015)在桑沟湾进行的围隔培养实验表明, 栉孔扇贝(Chlamys farreri)增加了海水中无机磷酸盐的浓度, 从而刺激了各生物类群的生长(Lu et al., 2015)。
3.2.4 病毒对微食物网结构的影响
病毒在海洋中数量巨大, 是微食物环的组成成分之一。病毒具有宿主特异性, 通过病毒的特异性侵染和裂解, 可以有效地调节细菌、藻类等微食物环组分的丰度和群落结构,每天平均有 20%~40%的海洋细菌死亡是由病毒介导的, 与浮游动物摄食造成的影响相当(Suttle, 2007)。在部分研究海区, 病毒的调节作用更为明显, 如在夏季南海西部调查中, 有26.44%~96.08%(平均值 77.82%)的细菌生产被病毒裂解(Chen et al., 2011)。
3.3 浮游植物对微食物网结构的影响
有些浮游植物会产生化感物质, 由于这些化感物质对不同生物的作用不同, 这些浮游植物的存在对微食物网的结构会产生影响。例如, 甲藻塔玛亚历山大藻(Alexandrium tamarense)会释放化感物质, 这种物质对细菌影响不大,但是会对异养鞭毛虫和纤毛虫产生抑制作用,使异养鞭毛虫和纤毛虫产生自溶现象, 增加了水体中的溶解有机物质, 从而增加了水体中的细菌生物量(Weissbach et al., 2011)。
不同种类的浮游植物产生的 DOM 的组成不同, 可能会影响细菌群落结构, 进而影响微食物网结构。在一个中型围隔(1000L)的培养实验中, 3个围隔分别为接种球形棕囊藻(Phaeocystis globosa)和硅藻(Chaetoceros decipiens和 Thalassiosira pseudonana)的实验组, 以及不接种任何生物的对照组, 以观察水华对微食物网的影响(Passow et al., 2007)。在培养过程中, 3个围隔中的浮游植物发生明显不同的变化: 对照组中没有发生水华, 接种球形棕囊藻的围隔中发生了小规模的水华, 接种硅藻的围隔中发生了较大规模的水华。结果表明,不同水华过程中细菌丰度的变化趋势相似,但是变化幅度不同; 不同种类浮游植物会对细菌的群落组成产生影响; 接种浮游植物后异养鞭毛虫丰度比对照组升高, 在接种球形棕囊藻的围隔中粒径大于 6μm的异养鞭毛虫丰度较大(Passow et al., 2007; Murray et al.,2007)。
3.4 营养盐对微食物网结构的影响
营养盐对微食物网的影响既有直接影响也有间接影响: 直接影响是细菌会利用营养盐, 营养盐浓度和结构的不同会导致细菌丰度的不同; 间接影响是营养盐浓度和结构的不同会导致浮游植物的类群组成和DOC的不同, 从而间接影响细菌。然而, 这些影响交织在一起, 很难辨别是哪个途径占主要作用。一般来说, 高营养盐浓度下, 浮游植物生物量增加, 释放较多的 DOC, 细菌有了营养盐和DOC, 其生长会得到促进, 但是营养盐升高是否导致异养鞭毛虫和纤毛虫的生物量也升高却有很大的不确定性。在人工添加营养盐的情况下, 细菌的生物量升高, 异养鞭毛虫和纤毛虫的生物量也升高(Andersson et al., 2006);但也有研究发现, 在亚得里亚海(Adriatic Sea),随着营养盐浓度的降低, 细菌丰度则会降低, 但异养鞭毛虫的丰度反而增加(Šolić et al., 2010)。
大气沉降到海里的沙尘对微食物网的影响复杂多变。在地中海进行的两次中型围隔实验的结果完全不同: 第一次添加沙尘实验中,细菌群落发生了明显变化, 但病毒和异养鞭毛虫的丰度没有明显变化; 第二次添加沙尘实验中, 细菌群落没有变化, 但是细菌的丰度降低, 病毒的丰度增加(Pulido-Villena et al., 2014)。Romero等(2011)的沙尘添加实验中,细菌的丰度先升高, 48 h后细菌丰度降低, 鞭毛虫丰度升高(Romero et al., 2011)。在南海寡营养海区添加大气气溶胶后, 细菌的丰度并没有明显增加, 但是不同细菌类群的反应不同, 聚球藻蓝细菌的丰度降低, 但是群落组成的变化很小(Guo et al., 2012)。
3.5 其他因素对微食物网结构的影响
除以上内容外, 还有一些因素也会导致微食物网结构发生变化。扰动可以使鞭毛虫和纤毛虫对细菌的摄食压力减小, 转而摄食比细菌大的颗粒, 从而使得细菌的丰度增加(Peters et al., 2002)。分散剂常被用来治理原油泄漏事故导致的海上漂浮原油, Ortmann等(2012)用围隔方法研究了原油泄露和分散剂对浮游微食物网的影响, 在自然海水中单独加入原油导致纤毛虫生物量增加, 但是单独加入分散剂或同时加入分散剂和原油会导致细菌生物量增加, 纤毛虫生物量则受到抑制。近年来, 缺氧是备受关注的海洋灾害之一, 它会对微食物网产生影响。在缺氧区, 浮游纤毛虫的群落结构会发生很大变化, 超过 80%的纤毛虫是摄食细菌的Uronema spp.(Stauffer et al., 2013)。
4 结语
迄今为止, 海洋微食物网已经历了 30余年的研究, 随着研究手段的进步, 人们逐步认识到微食物网对海洋生态系统产生的重要影响。近年来, 学者们从多角度对海洋微食物网的结构开展了研究, 不同海区微食物网各类群丰度、生物量的时间和空间变化研究有很多报道, 微食物网的结构可受空间、季节、摄食、浮游植物、营养盐等多种因素影响。国际上关于微食物网结构的系统研究还有不足, 关于自然海区微食物网结构的动态变化研究还需要继续深入开展。
张武昌, 陈雪, 李海波, 等. 2014. 海洋浮游桡足类摄食纤毛虫的研究. 海洋与湖沼, 45(4): 764-775
Agawin N S R, Duarte C M, Agustí S. 1998. Growth and abundance of Synechococcus sp. in a Mediterranean Bay: seasonality and relationship with temperature. Marine Ecology Progress Series, 170: 45-53
Andersson A, Samuelsson K, Haecky P, et al. 2006. Changes in the pelagic microbial food web due to artificial eutrophication. Aquatic Ecology, 40(3): 299-313
Azam F, Fenchel T, Field J G, et al. 1983. The ecological role of water-column microbes in the sea. Marine Ecology-Progress Series, 10(3): 257-263
Azam F, Malfatti F. 2007. Microbial structuring of marine ecosystems. Nature Reviews Microbiology, 5(10): 782-791
Calbet A, Landry M R, Nunnery S. 2001. Bacteriaflagellate interactions in the microbial food web of the oligotrophic subtropical North Pacific. Aquatic Microbial Ecology, 23: 283-292
Calbet A, Landry M R. 1999. Mesozooplankton influences on the microbial food web: direct and indirect trophic interactions in the oligotrophic open ocean. Limnology and Oceanography, 44(6): 1370-1380
Calvo-Díaz A, Morán X A G. 2006. Seasonal dynamics of picoplankton in shelf waters of the southern Bay of Biscay. Aquatic Microbial Ecology, 42(2): 159-174
Caron D A. 1991. Evolving role of protozoa in aquatic nutrient cycles//Reid P C, Turley C M, Burkill P H. Protozoa and Their Role in Marine Processes. Berlin Heidelberg: Springer: 387-415
Chen B Z, Liu H B, Wang Z L. 2009. Trophic interactions within the microbial food web in the South China Sea revealed by size-fractionation method. Journal of Experimental Marine Biology and Ecology, 368(1): 59-66
Chen B Z, Liu H B. 2010. Trophic linkages between grazers and ultraplankton within the microbial food web in subtropical coastal waters. Marine Ecology Progress Series, 407: 43-53
Chen X H, Liu H B, Weinbauer M, et al. 2011. Viral dynamics in the surface water of the western South China Sea in summer 2007. Aquatic Microbial Ecology, 63(2): 145-160
Chen X, Zhang W C, Denis M, et al. 2016. The pelagic microbial food web structure in Sanggou Bay, Yellow Sea: spatial variation over four successive seasons. Biogeosciences doi:10.5194/bg-2015-665
Chisholm S W, Frankel S L, Goericke R, et al. 1992. Prochlorococcus marinus nov. gen. nov. sp.: an oxyphototrophic marine prokaryote containing divinyl chlorophyll a and b. Archives of Microbiology, 157(3): 297-300
Chisholm S W, Olson R J, Zettler E R, et al. 1988. A novel free-living prochlorophyte abundant in the oceanic euphotic zone. Nature, 334(6180): 340-343
Christaki U, Van Wambeke F, Lefevre D, et al. 2011. Microbial food webs and metabolic state across oligotrophic waters of the Mediterranean Sea during summer. Biogeosciences, 8(7): 1839-1852
Condon R H, Steinberg D K, del Giorgio P A, et al. 2011. Jellyfish blooms result in a major microbialrespiratory sink of carbon in marine systems. Proceedings of the National Academy of Sciences of the United States of America, 108(25): 10225-10230
Dinasquet J, Titelman J, Møller L F, et al. 2012. Cascading effects of the ctenophore Mnemiopsis leidyi on the planktonic food web in a nutrientlimited estuarine system. Marine Ecology Progress Series, 460: 49-61
Dupuy C, Le Gall S, Hartmann H J, et al. 1999. Retention of ciliates and flagellates by the oyster Crassostrea gigas in French Atlantic coastal ponds: protists as a trophic link between bacterioplankton and benthic suspension-feeders. Marine Ecology Progress Series, 177: 165-175
Dupuy C, Pastoureaud A, Ryckaert M, et al. 2000a. Impact of the oyster Crassostrea gigas on a microbial community in Atlantic coastal ponds near La Rochelle. Aquatic Microbial Ecology, 22(3): 227-242
Dupuy C, Ryckaert M, Le Gall S, et al. 2007. Seasonal variations in planktonic community structure and production in an Atlantic coastal pond: the importance of nanoflagellates. Microbial Ecology, 53(4): 537-548
Dupuy C, Vaquer A, Lam-Höai T, et al. 2000b. Feeding rate of the oyster Crassostrea gigas in a natural planktonic community of the Mediterranean Thau Lagoon. Marine Ecology Progress Series, 205: 171-184
Fenchel T. 1982. Ecology of heterotrophic microflagellates. IV. Quantitative occurrence and importance as bacterial consumers. Marine Ecology Progress Series, 9(1): 35-42
Fenchel T. 2008. The microbial loop-25 years later. Journal of Experimental Marine Biology and Ecology, 366(1-2): 99-103
Fournier J, Dupuy C, Bouvy M, et al. 2012. Pearl oysters Pinctada margaritifera grazing on natural plankton in Ahe atoll lagoon (Tuamotu Archipelago, French Polynesia). Marine Pollution Bulletin, 65(10-12): 490-499
Fuhrman J A, Azam F. 1980. Bacterioplankton secondary production estimates for coastal waters of British Columbia, Antarctica, and California. Applied and Environmental Microbiology, 39(6): 1085-1095
Garrison D L, Gowing M M, Hughes M P, et al. 2000. Microbial food web structure in the Arabian Sea: a US JGOFS study. Deep Sea Research Part II: Topical Studies in Oceanography, 47(7-8): 1387-1422
Gasol J M, Vaque D. 1993. Lack of coupling between heterotrophic nanoflagellates and bacteria: a general phenomenon across aquatic systems. Limnology and Oceanography, 38(3): 657-665
Gasol J M. 1994. A framework for the assessment of top-down vs. bottom-up control of heterotrophic nanoflagellate abundance. Marine Ecology Progress Series, 113(3): 291-300
Goldman J C, Caron D A. 1985. Experimental studies on an omnivorous microflagellate: implications for grazing and nutrient regeneration in the marine microbial food chain. Deep Sea Research Part A. Oceanographic Research Papers, 32(8): 899-915
Guillou L, Jacquet S, Chrétiennot-Dinet M J, et al. 2001. Grazing impact of two small heterotrophic flagellates on Prochlorococcus and Synechococcus. Aquatic Microbial Ecology, 26(2): 201-207
Guo C, Yu J, Ho T Y, et al. 2012. Dynamics of phytoplankton community structure in the South China Sea in response to the East Asian aerosol input. Biogeosciences, 9(4): 1519-1536
Hansen B, Bjornsen P K, Hansen P J. 1994. The size ratio between planktonic predators and their prey. Limnology and Oceanography, 39(2): 395-403
Iriarte A, Madariaga I, Revilla M, et al. 2003. Short-term variability in microbial food web dynamics in a shallow tidal estuary. Aquatic Microbial Ecology, 31(2): 145-161
Jacquet S, Prieur L, Nival P, et al. 2010. Structure and variability of the microbial community associated to the Alboran Sea frontal system (Western Mediterranean) in winter. Journal of Oceanography, Research and Data, 3(4): 47-75
Johnson P W, Sieburth J M. 1982. In-situ morphology and occurrence of eucaryotic phototrophs of bacterial size in the picoplankton of estuarine and oceanic waters. Journal of Phycology, 18(3): 318-327
Kreeger D A, Newell R. 1996. Ingestion and assimilation of carbon from cellulolytic bacteria and heterotrophic flagellates by the mussels Geukensia demissa and Mytilus edulis (Bivalvia, Mollusca). Aquatic Microbial Ecology, 11(3): 205-214
Kryger J, Riisgård H U. 1988. Filtration rate capacities in 6 species of European freshwater bivalves. Oecologia, 77(1): 34-38
Le Gall S, Bel Hassen M, Le Gall P. 1997. Ingestion of a bacterivorous ciliate by the oyster Crassostrea gigas: protozoa as a trophic link between picoplankton and benthic suspension-feeders. Marine Ecology Progress Series, 15(1-3): 301-306
Leakey R J G, Burkill P H, Sleigh M A. 1992. Planktonic ciliates in Southampton Water: abundance, biomass, production, and role in pelagic carbon flow. Marine Biology, 114(1): 67-83
Li W K W, Carmack E C, McLaughlin F A, et al. 2013. Space-for-time substitution in predicting the state of picoplankton and nanoplankton in a changing Arctic Ocean. Journal of Geophysical Research: Oceans, 118(10): 5750-5759
Li W K W. 1998. Annual average abundance of heterotrophic bacteria and Synechococcus in surface ocean waters. Limnology and Oceanography, 43(7): 1746-1753
Liu H B, Chang J, Tseng C M, et al. 2007. Seasonal variability of picoplankton in the Northern South China Sea at the SEATS station. Deep Sea Research Part II: Topical Studies in Oceanography, 54(14-15): 1602-1616
Liu H B, Suzuki K, Minami C, et al. 2002. Picoplankton community structure in the subarctic Pacific Ocean and the Bering Sea during summer 1999. Marine Ecology Progress Series, 237: 1-14
López-Urrutia A. 2013. Macroscopic patterns in marine plankton//Levin S A. Encyclopedia of Biodiversity. 2nd ed. Waltham, Massachusetts: Academic: 667-680
Lu J C, Huang L F, Xiao T, et al. 2015. The effects of Zhikong scallop (Chlamys farreri) on the microbial food web in a phosphorus-deficient mariculture system in Sanggou Bay, China. Aquaculture, 448: 341-349
McManus G B, Fuhrman J A. 1990. Mesoscale and seasonal variability of heterotrophic nanoflagellate abundance in an estuarine outflow plume. Marine Ecology Progress Series, 61(3): 207-213
Michaels A F, Silver M W. 1988. Primary production, sinking fluxes and the microbial food web. Deep Sea Research Part A. Oceanographic Research Papers, 35(4): 473-490
Miki T, Jacquet S. 2008. Complex interactions in the microbial world: underexplored key links between viruses, bacteria and protozoan grazers in aquatic environments. Aquatic Microbial Ecology, 51(2): 195-208
Murphy L S, Haugen E M. 1985. The distribution and abundance of phototrophic ultraplankton in the North Atlantic. Limnology and Oceanography, 30(1): 47-58
Murray A E, Arnosti C, De La Rocha C L, et al. 2007. Microbial dynamics in autotrophic and heterotrophic seawater mesocosms. II. Bacterioplankton community structure and hydrolytic enzyme activities. Aquatic Microbial Ecology, 49(2): 123-141
Ortmann A C, Anders J, Shelton N, et al. 2012. Dispersed oil disrupts microbial pathways in pelagic food webs. PLoS One, 7(7): e42548
Passow U, De La Rocha C L, Arnosti C, et al. 2007. Microbial dynamics in autotrophic and heterotrophic seawater mesocosms. I. Effect of phytoplankton on the microbial loop. Aquatic Microbial Ecology, 49(2): 109-121
Peters F, Marrasé C, Havskum H, et al. 2002. Turbulence and the microbial food web: effects on bacterial losses to predation and on community structure. Journal of Plankton Research, 24(4): 321-331
Pomeroy L R, Wiebe W J. 1988. Energetics of microbial food webs. Hydrobiologia, 159(1): 7-18
Pulido-Villena E, Baudoux A C, Obernosterer I, et al. 2014. Microbial food web dynamics in response to a Saharan dust event: results from a mesocosm study in the oligotrophic Mediterranean Sea. Biogeosciences, 11(19): 5607-5619
Rassoulzadegan F, Sheldon R W. 1986. Predator-prey interactions of nanozooplankton and bacteria in an oligotrophic marine environment. Limnology and Oceanography, 31(5): 1010-1029
Reckermann M, Veldhuis M J W. 1997. Trophic interactions between picophytoplankton and microand nanozooplankton in the western Arabian Sea during the NE monsoon 1993. Aquatic Microbial Ecology, 12(3): 263-273
Romero E, Peters F, Marrasé C, et al. 2011. Coastal Mediterranean plankton stimulation dynamics through a dust storm event: an experimental simulation. Estuarine, Coastal and Shelf Science, 93(1): 27-39
Ryther J H. 1969. Photosynthesis and fish production in the sea. Science, 166(3901): 72-76
Samuelsson K, Andersson A. 2003. Predation limitation in the pelagic microbial food web in an oligotrophic aquatic system. Aquatic Microbial Ecology, 30(3): 239-250
Sanders R W, Caron D A, Berninger U G. 1992. Relationships between bacteria and heterotrophic nanoplankton in marine and fresh waters: an inter-ecosystem comparison. Marine Ecology Progress Series, 86(1): 1-14
Savenkoff C, Vézina A, Roy S, et al. 2000. Export of biogenic carbon and structure and dynamics of the pelagic food web in the Gulf of St. Lawrence Part 1. Seasonal variations. Deep Sea Research Part II: Topical Studies in Oceanography, 47(3): 585-607
Sheldon R W, Prakash A, Sutcliffe Jr W H. 1972. The size distribution of particles in the ocean. Limnology and Oceanography, 17(3): 327-340
Sherr B F, Sherr E B, Hopkinson C S. 1988. Trophic interactions within pelagic microbial communities: indications of feedback regulation of carbon flow. Hydrobiologia, 159(1): 19-26
Shinada A, Ban S, Yamada Y, et al. 2005. Seasonal variations of plankton food web structure in the coastal water off Usujiri southwestern Hokkaido, Japan. Journal of Oceanography, 61(4): 645-654
Sieburth J M, Smetacek V, Lenz J. 1978. Pelagic ecosystem structure: heterotrophic compartments of the plankton and their relationship to plankton size fractions. Limnology and Oceanography, 23(6): 1256-1263
Sipura J, Lores E, Snyder R A. 2003. Effect of copepods on estuarine microbial plankton in short-term microcosms. Aquatic Microbial Ecology, 33(2): 181-190
Šolić M, Krstulović N, Kušpilić G, et al. 2010. Changes in microbial food web structure in response to changed environmental trophic status: a case study of the Vranjic Basin (Adriatic Sea). Marine Environmental Research, 70(2): 239-249
Stauffer B A, Schnetzer A, Gellene A G, et al. 2013. Effects of an acute hypoxic event on microplankton community structure in a coastal harbor of southern California. Estuaries and Coasts, 36(1): 135-148
Suttle C A. 2007. Marine viruses-major players in the global ecosystem. Nature Reviews Microbiology, 5(10): 801-812
Van Duyl F C, Bak R P M, Kop A J, et al. 1990. Bacteria, auto- and heterotrophic nanoflagellates, and their relations in mixed, frontal and stratified waters of the North Sea. Netherlands Journal of Sea Research, 26(1): 97-109
Waterbury J B, Watson S W, Guillard R R L, et al. 1979. Widespread occurrence of a unicellular, marine, planktonic, cyanobacterium. Nature, 277(5694): 293-294
Weinbauer M G, Peduzzi P. 1995. Significance of viruses versus heterotrophic nanofiagellates for controlling bacterial abundance in the northern Adriatic Sea. Journal of Plankton Research, 17(9): 1851-1856
Weissbach A, Rudström M, Olofsson M, et al. 2011. Phytoplankton allelochemical interactions change microbial food web dynamics. Limnology and Oceanography, 56(3): 899-909
West E J, Pitt K A, Welsh D T, et al. 2009. Top-down and bottom-up influences of jellyfish on primary productivity and planktonic assemblages. Limnology and Oceanography, 54(6): 2058-2071
Wikner J, Hagström Å. 1988. Evidence for a tightly coupled nanoplanktonic predator-prey link regulating the bacterivores in the marine environment. Marine Ecology Progress Series, 50(1): 137-145
Wilhelm S W, Suttle C A. 1999. Viruses and nutrient cycles in the sea. Viruses play critical roles in the structure and function of aquatic food webs. Bioscience, 49(10): 781-788
The Microbial Food Web Structure and Influencing Factors: A Review
ZHANG Wu-Chang1,2*, CHEN Xue1,2,3, ZHAO Yuan1,2, ZHAO Li1,2, XIAO Tian1,2
(1. Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. Laboratory of Marine Ecology and Environmental Science, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China; 3. University of Chinese Academy of Sciences, Beijing 100049, China) *Corresponding author, E-mail: wuchangzhang@qdio.ac.cn
The microbial food web, which is comprised of phototrophic picoplankton, heterotrophic bacteria, virus and protozoa, is ubiquitous in marine waters. Members of the microbial food web may play important roles in the energy and nutrients cycle in the ocean. In microbial food webs, different types of interactions occur between microorganisms themselves and with microzooplankton. The functioning of the microbial food webs in different ecosystems has been well illustrated and explained. However, little has been known about the structure of the microbial food web since the first study in 2000 by Garrison et al, The abundances of different groups in the microbial food web are changing under different environments, but they mostly fall into a certain range: Ciliate 10cell/mL, nanoflagellates 103cell/mL, picoplankton 104cell/mL, cyanobacteria 104~105cell/mL, heterotrophic bacteria 106cell/mL, virus 107particle/ml. The optimum linear size ratio between predators and their optimal prey is 10︰1, although this ratio is lower in field studies. For example, the ratio is 3︰1 for nanoflagellates and 8︰1 for ciliates. The ratio of picoplankton to nanoplankton abundance (pico︰nano) is a scale-free index to study the microbial food web structure. The structure of the microbial food web depends, on one hand, on the type of environments and, on the other hand, on forcing factors exerting on these environments such as predation, phytoplankton and nutrient supplement. When studying the spatial variation of microbial food web structure, researchers usually use observation results of the abundance of microbial food web components from different water masses as an alternative plan. Regarding to the seasonal variation, the abundances and biomasses of microbial food web components are affected by the hydrological environment. Predation can affect the microbial food web structure through the following pathways: ①Mesozooplankton feeding; ②Mesozooplankton consumes microzooplankton and affecting lower trophic level through cascading predation effects;③Mesozooplankton could affect bacteria and other lower trophic level component by releasing dissolved organic matter and nutrients. Phytoplankton could release allelochemical matter and dissolved organic matter to change microbial food web dynamics. The concentration and variation of nutrients can have a direct or indirect impact on microbial food web structure.
microbial food web; structure; picoplankton; nanoflagellates; ciliates
P735
10.12036/hykxjk20160725002
* 资助项目: 国家自然科学基金(41576164, 41306161); 国家重点基础研究发展计划资助项目 (2014CB441504);中国科学院战略性先导科技专项(XDA11030202.2)。
① 通讯作者: 张武昌, 男, 博士, 研究员, 主要从事海洋微食物网生态学研究。E-mail: wuchangzhang@ qdio.ac.cn
2016-07-25, 收修改稿日期: 2016-07-29
