中国沿海6个花鲈群体的形态差异分析
2016-03-06侯吉伦赵雅贤任建功王桂兴陈睿毅骆明飞王玉芬刘海金
侯吉伦,赵雅贤,任建功,王桂兴,陈睿毅,骆明飞,王玉芬,刘海金
(1.中国水产科学研究院北戴河中心实验站,河北秦皇岛 066100;2.浙江省海洋水产研究所,舟山 316021;3.珠海市现代农业发展中心,广东珠海 519000)
中国沿海6个花鲈群体的形态差异分析
侯吉伦1,赵雅贤1,任建功1,王桂兴1,陈睿毅2,骆明飞3,王玉芬1,刘海金1
(1.中国水产科学研究院北戴河中心实验站,河北秦皇岛 066100;2.浙江省海洋水产研究所,舟山 316021;3.珠海市现代农业发展中心,广东珠海 519000)
通过聚类分析、主成分分析和判别分析等多元分析方法,对东港、绥中、秦皇岛、青岛、舟山以及珠海等6个花鲈(Lateolabrax maculatus)地理群体的8个形态比例性状进行了研究。聚类分析和主成分分析表明,6个花鲈群体可被分为两支:来自黄海、渤海海域的东港、秦皇岛、绥中和青岛群体为一支(北方群体);东海海域的舟山群体和南海海域的珠海群体为另一支(南方群体)。南北群体间有一定程度的形态分化,和舟山群体相比,珠海群体和北方群体的亲缘关系更近。主成分分析获得的3个主成分方差贡献率分别为31.726%、27.744%和14.075%,累计贡献率为73.545%。利用8个变量构建的6个地理种群的判别公式,判别准确率在63.6%~84.4%之间,综合判别准确率72.7%。本研究结果为花鲈地理种群判别、种质资源评价以及良种选育等提供了基础资料。
花鲈;多元分析;形态差异;逃逸
形态是生物体的重要性状。通过对形态数据的测量,并进行相应的统计分析,可以比较个体或群体间的异同。形态差异分析常用的方法是多元分析法,包含多元方差分析、聚类分析、因子分析、主成分分析和判别分析等分析方法。多元分析能够在多个指标和对象相互关联的情况下,分析它们的统计规律。在鱼类研究中,多元分析被广泛地应用于不同地理群体形态差异[1-5]、杂交种后代与亲本形态差异[6-9]以及品种判别[10-11]等领域。
花鲈(Lateolabrax maculatus)俗称七星鲈鱼、板鲈、鲈鱼、青鲈、花寨等,隶属于鲈形目(Perciformes),鮨科(Serranidae),花鲈属,是一种广温性近岸中下层鱼类,广泛分布于中国沿海及朝鲜半岛。花鲈具有较强的盐度适应能力,无论在海淡水中都能生存[12]。随着人工繁殖、饵料等关键技术的突破,花鲈的养殖业迅速发展,并形成了池塘、海水网箱、工厂化等多种养殖模式。近年来,我国学者针对花鲈开展了营养学[13-15]、繁殖生理学[16]、遗传学[17-19]等相关研究。关于花鲈不同地理种群形态差异的研究,目前尚未见报道。本文利用聚类分析、主成分分析和判别分析等3种分析方法,对来东港、绥中、秦皇岛、青岛、舟山以及珠海海域的6个花鲈地理群体的形态差异进行了研究,旨在为花鲈的地理种群判别、种质资源评价以及良种选育等提供基础资料。
1 材料与方法
1.1 实验材料
实验用花鲈于2015年9月至12月间,利用拖网法,分别采自于辽宁东港、绥中、河北秦皇岛、山东青岛、浙江舟山以及广东珠海等6个海域(图1)。这6个海域覆盖了我国渤海、黄海、东海以及南海等海区,具有一定的代表性。测定时,先用吸水纸将样品体表的水分吸干,再用电子天平称量体重(精确到0.01 g),最后用数码相机对每一尾鱼进行标准图片采集。每个海域的采样尾数、体重和体长范围见表1。
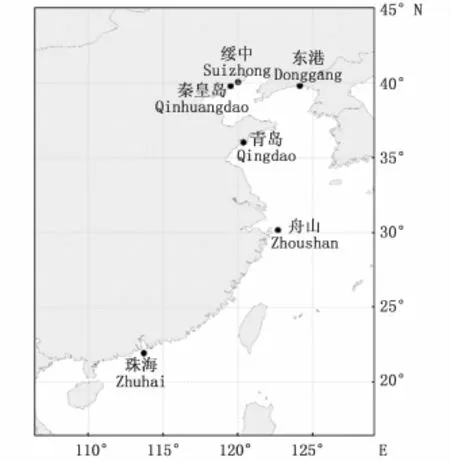
图1 本研究采样点地理位置Fig.1 Locations of sampling sites in this study
1.2 数据测量
采用了13个解剖学坐标点,对全长(TL)、体长(BL)、头长(HL)、尾柄长(CPL)、体高(WH)、头高(HH)、眼径(ED)、第一背鳍下体高(FWH)和尾柄高(CPH)等9个形态参数进行了测量(图2)。测量在Motic Images Plus 2.0软件中进行,并用比例尺对测量数据进行校正(精确到0.01 cm)。共测量了253 ind样品的2 277个形态数据。
1.3 数据分析
为消除样品规格大小差异对形态参数的影响,将每尾鱼测量获得的全长、头长、尾柄长、体高、头高、眼径、第一背鳍下体高和尾柄高等8个形态参数分别除以体长,转换成8个形体比例参数:全长/体长(TL/BL)、头长/体长(HL/BL),尾柄长/体长(CPL/BL)、体高/体长(WH/BL)、头高/体长(HH/BL)、眼径/体长(ED/BL)、第一背鳍下体高/体长(FWH/BL)、尾柄高/体长(CPH/BL)。
采用SPSS 19.0对各地理群体8个形态比例性状进行方差分析、聚类分析、主成分分析和判别分析。
1.3.1 聚类分析
计算各地理群体8个形态比例性状的平均值,并用平均值进行基于欧氏距离的最短距离系统聚类[20],计算群体间的欧氏距离,绘制聚类图。
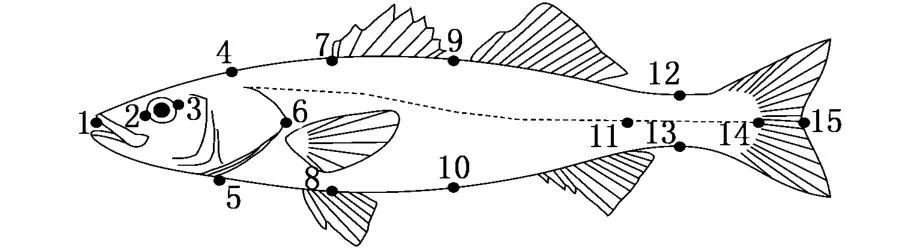
图2 花鲈形态性状测量部位示意图Fig.2 Sketch of measuration positions of morphological traits of spotted sea bass(Lateolabrax maculatus)
1.3.2 主成分分析
先对8个形态比例参数进行校正,然后用校正值进行主成分分析。
1.3.3 判别分析
判别准确率的计算公式为:

式中,Ai为第i个群体判别正确的尾数,Bi为第i个群体的实际尾数,k为群体数。
2 结果与分析
2.1 形态比例参数统计量分析
花鲈6个地理群体的8个形态比例性状单因素方差分析结果显示,各群体之间既有差异显著的性状,又有差异不显著的比例性状,无特定规律(表2)。例如,全长/体长这一比例参数上,绥中、舟山和珠海3个群体显著大于其它3个群体(P<0.05),青岛群体的值最小。头长/体长,秦皇岛群体和舟山群体差异不显著,但小于其它4个群体(4个群体之间差异不显著)。而在尾柄高/体高这一性状上,绥中群体显著高于其他群体,秦皇岛群体居其次,随后是东港和青岛群体,最小的是舟山和珠海群体。
2.2 聚类分析
8个形态比例性状平均值的计算欧式距离并进行聚类分析结果显示,6个地理群体聚为两支:东港、秦皇岛、绥中和青岛为一支;舟山和珠海为另一支。在东港、秦皇岛、绥中和青岛这一支中,东港、秦皇岛和绥中群体聚为一支,这三者再和青岛群体聚为一支。舟山群体和绥中群体在形态比例性状上距离最远(表3,图3)。
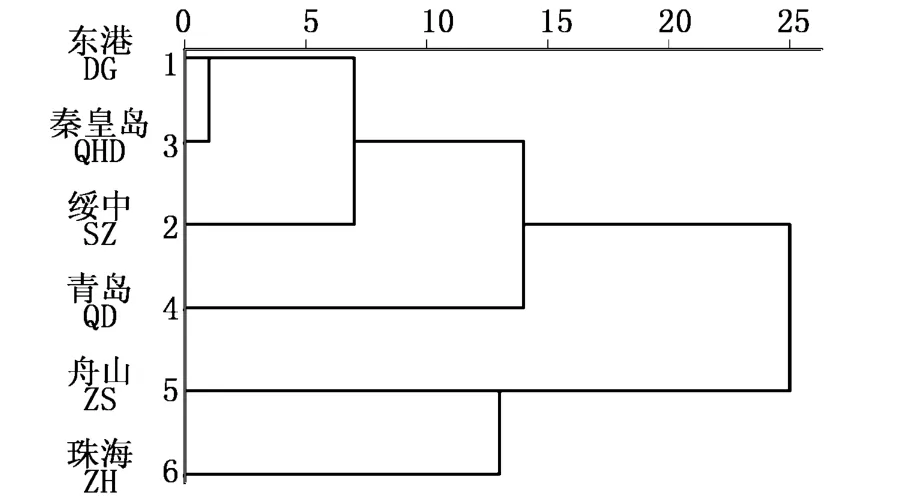
图3 花鲈6个地理群体的聚类分析图Fig.3 Dendrogram of cluster analysis of 6 geographic populations of spotted sea bass(Lateolabrax maculatus)

表1 不同花鲈群体采样地点、数量、体质量及体长信息Tab.1 Sampling location,number,body weight and body length of spotted sea bass(Lateolabrax maculatus)
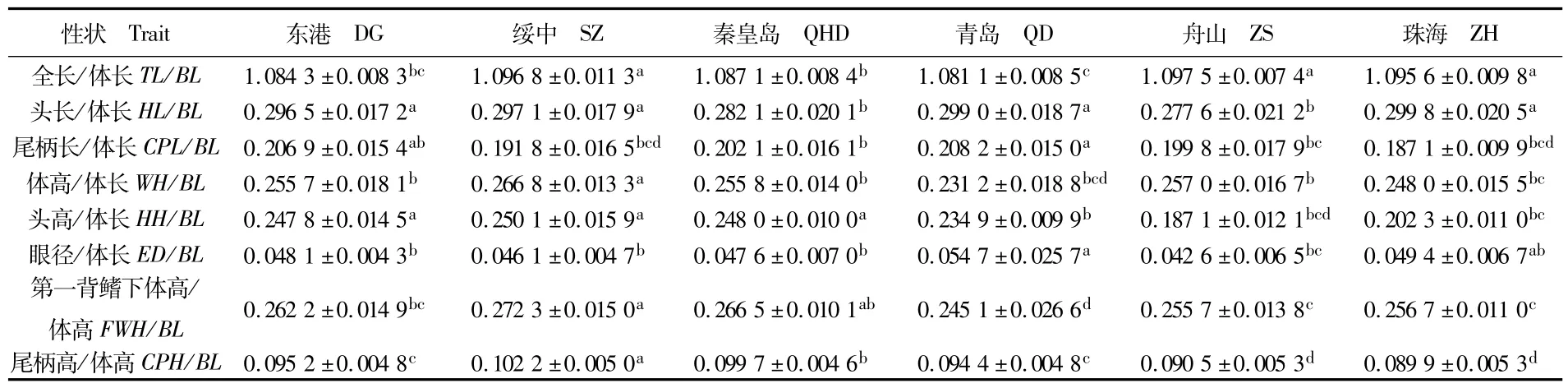
表2 花鲈6个不同地理群体8个可量性状方差分析结果Tab.2 ANOVA results of 8 proportional characters of 6 geographical sea bass(Lateolabrax maculatus)populations

表3 花鲈6个地理群体的欧式距离Tab.3 Euclidean distances of 6 geographic populations of spotted sea bass(Lateolabrax maculatus)
2.3 主成分分析
分别对花鲈不同地理种群的8个形态比例性状进行主成分分析,获得了3个主成分。各主成分的负荷值、方差贡献率和累计贡献率见表4。结果显示,3个主成分的方差贡献率分别为31.726%、27.744%和14.075%,累计贡献率73.545%。此3个主成分包含变异的大部分,对其进行分析可基本反映综合指标所隐含的信息。主成分1起主要作用的是第一背鳍下体高/体长、体高/体长,负荷值分别为0.883和0.835;主成分2起主要作用的是头高/体长,负荷值为0.908;主成分3起主要作用的是眼径/体长,负荷值为0.692。主成分1和2、2和3的散布图分别见图4和图5。从图中可以发现,主成分1和2以及2和3均可将东港、绥中、秦皇岛和青岛4个群体分为一簇,且东港、绥中和秦皇岛三个群体个体间重叠明显,表面遗传距离较近;舟山和珠海群体为另一簇。珠海群体有部分个体和东港、绥中、秦皇岛和青岛群体一簇相重叠,说明珠海群体的部分个体的遗传距离和上述4个群体较近。
2.4 判别分析
对所有8个形态比例性状进行判别分析。F检验结果显示,6个花鲈群体间形态差异显著(P<0.05),判别效果较好。判别准确率在63.6%~84.4%之间,综合判别率为72.7%。其中舟山群体的判别准确率为84.4%,在所有6个地理种群中最高(表5),说明和其它几个群体的形态差异较大,其次分别是珠海群体、青岛群体、绥中群体、秦皇岛群体和东港群体。此结果和聚类分析结果一致。
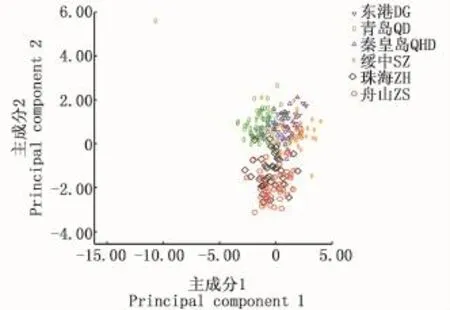
图4 花鲈6个地理群体第1、2主成分散布图Fig.4 Scatter diagram of principal component 1 and 2 of 6 geographic populations of spotted sea bass(Lateolabrax maculatus)
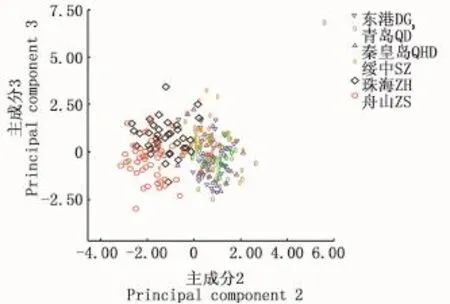
图5 花鲈6个地理群体第2、3主成分散布图Fig.5 Scatter diagram of principal component 2 and 3 of 6 geographic populations of spotted sea bass(Lateolabrax maculatus)

表4 花鲈主成分分析因子负荷值及主成分贡献率Tab.4 The load values,contribution rates of 3 principal components of spotted sea bass(Lateolabrax maculatus)by principal components analysis

表5 花鲈6个地理群体的判别结果Tab.5 Discriminant results of 6 geographic populations of spotted sea bass(Lateolabrax maculatus)
利用逐步判别法,对8个形态比例性状的特征值进行了分析,建立了花鲈6个地理种群的判别函数。函数中X1-X8分别代表全长/体长、头长/体长,尾柄长/体长、体高/体长、头高/体长、眼径/体长、第一背鳍下体高/体长、尾柄高/体长等8个形态比例性状。各判别公式如下:
东港群体:
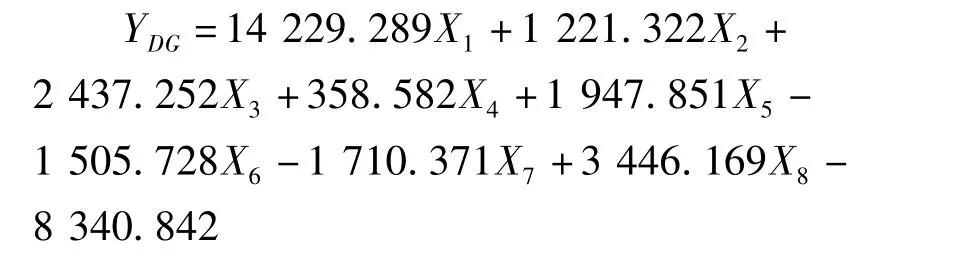
绥中群体:
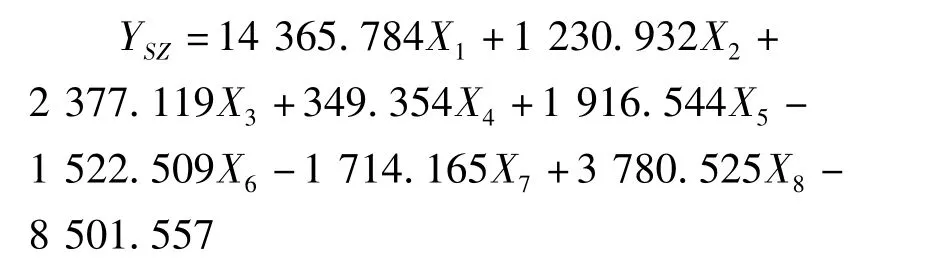
秦皇岛群体:
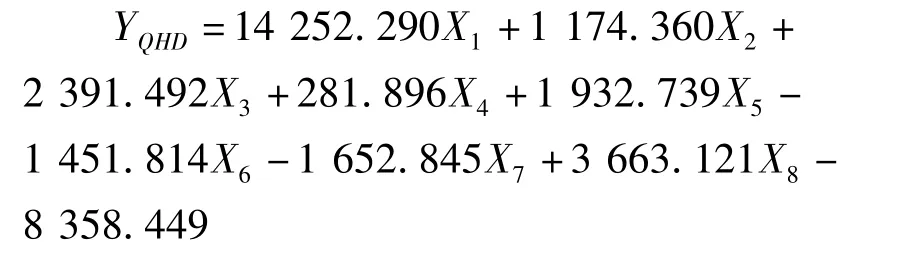
青岛群体:
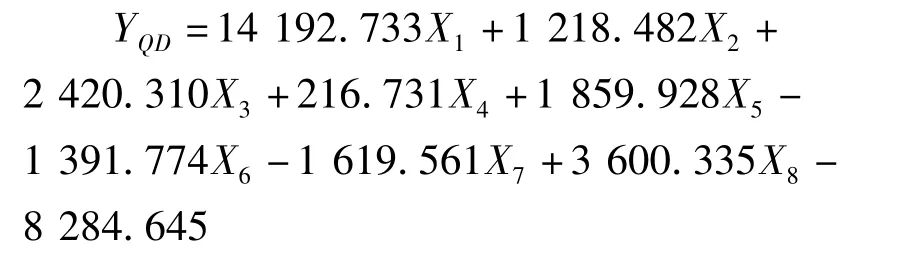
舟山群体:

珠海群体:
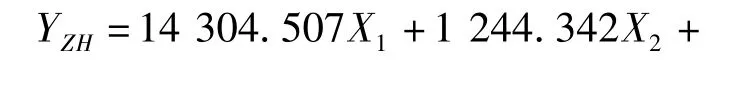
3 讨论
本研究利用聚类分析、主成分分析和判别分析等多元分析方法,对中国沿海花鲈6个地理种群的形态差异进行了分析。研究结果显示,这3种分析方法通过不同角度进行分析,虽然所获得的结果具有一定的相似性,但彼此间不能代替。聚类分析和主成分分析表明,按照地理距离的远近,6个花鲈群体可被分为两支:来自黄海、渤海海域的东港、秦皇岛、绥中和青岛群体为一支;东海海域的舟山和南海海域的珠海群体为另一支。南北群体间有一定程度的形态分化。主成分分析获得的 3个主成分的方差贡献率分别为31.726%、27.744%和14.075%,累计贡献率73.545%,包含了总变量的大部分,说明可以用几个相互独立的因子来概括花鲈不同地理种群的形态差异。利用8个变量构建的6个地理种群的判别公式,综合判别准确率72.7%。
本结果基本体现了各群体地理关系的相关性,地理距离较近的群体的亲缘关系一般也较近。基因流是致使基因从一个群体到另一个群体或从一个亚群到另一个亚群的运转的机制[21]。地理距离可能是导致不同地理种群出现基因流的主要因素。胡自明等[17]利用RAPD方法,发现花鲈舟山群体与威海群体的遗传距离要小于同花鲈北海群体的遗传距离,作者认为这可能是因为花鲈舟山群体与威海群体之间地理距离较近,它们之间存在基因流动所致。本研究从形态学的角度证明了胡自明等[17]的观点。
珠海群体位于南海,而舟山群体位于东海,珠海和东港、秦皇岛、绥中以及青岛等地的地理距离要远于舟山和上述群体。按照地理距离远近和群体间亲缘关系的一般规律,珠海群体花鲈和4个北方群体的亲缘关系应为最远。但实际上,珠海群体和北方群体的亲缘关系要较舟山群体的近。这个结果可能与我国目前花鲈养殖业发展的地域特征具有一定的关系。珠海是我国最重要的花鲈养殖产区,2014年的人工养殖面积约为1 367 hm2,占当地海水养殖面积的2/3[22]。当地养殖苗种主要来源是北苗南养模式[12],即利用山东等地的野生花鲈亲鱼在当地或者福建等地进行人工繁殖,受精卵孵化培育成鱼苗时再运往珠海养殖;或者直接将受精卵运至珠海进行苗种培育和养成。在养殖过程中,是否因自然灾害或操作不当等原因,发生养殖花鲈逃逸到自然海域中,存活并繁殖,对自然海域野生群体的种群结构和遗传多样性产生了影响,由此导致本研究中珠海海域野生花鲈的亲缘关系和北方群体更近的结果?本研究中,珠海群体的采样点位于珠海万山岛附近,距离花鲈养殖主产地—斗门距离较近,因此,不能排除所采样品中含有逃逸个体或其所繁殖的后代。养殖逃逸是水产养殖中存在的一种现象,挪威沿海1989~1996年捕获的大西洋鲑中有34%~54%是养殖逃逸的鲑鱼,在峡湾捕获的鲑鱼中逃逸鱼占10%~21%[23]。现在已经有研究证明,养殖逃逸鱼对野生种群的遗传多样性会造成副作用[24-25]。因此,渔业主管部门加强对养殖户的宣传教育,杜绝人为原因造成的逃逸事件,切实保护好野生花鲈的遗传多样性是十分重要的。
[1] 明俊超,董在杰,梁政远,等.6个不同鲤群体的形态差异分析[J].广东海洋大学学报,2009,29(6):1-6.
MING JC,DONG Z J,LIANG Z Y,et al.Analysis of morphological variation in six common carp(Cyprinus carpio)populations[J].Journal of Guangdong Ocean University,2009,29(6):1-6.
[2] 孙 鹏,马 波,赵文阁.北方须鳅4个地理种群的形态差异与判别分析[J].水产学杂志,2014,27(2):14-19.
SUN P,MA B,ZHAO W G.Morphological variation and discriminant analysis in four geographical populations of loachBarbatula nuda[J].Chinese Journal of Fisheries,2014,27(2):14-19.
[3] 于 飞,张庆文,孔 杰,等.大菱鲆4个进口群体的形态差异分析[J].海洋水产研究,2008,29(5):27-32.
YU F,ZHANG Q W,KONG J,et al.Morphological variations and analyses of four imported turbot stocks,Scophthalmus maximus[J].Marine Fisheries Research,2008,29(5):27-32.
[4] 刘建勇,杨廷宝.我国沿海鲻鱼(Mugil cephalus)不同地理群体形态差异研究[J].海洋与湖沼,2009,40(5):572-576.
LIU J Y,YANG T B.Morphological variation of grey mulletMugil cephalusin different geographical populations along China coast[J].Oceanologia Et Limnologia Sinica,2009,40(5):572-576.
[5] 赵 建,朱新平,陈永乐,等.珠江卷口鱼不同地理种群的形态变异[J].动物学报,2007,53(5):921-927.
ZHAO J,ZHU X P,CHEN Y L,et al.Morphological variation of the ratmouth barbellPtychidio jordanifrom different geographic populations in the pearl river basin[J].Acta Zoologica Sinica,2007,53(5):921-927.
[6] 刘 苏,朱新平,陈昆慈,等.斑鳢、乌鳢及其杂交种形态差异分析[J].华中农业大学学报,2011,30(4):488-493.
LIU S,ZHU X P,CHEN K C,et al.Morphological variations ofChanna maculate,Channaargusand their hybrid(C.maculate♀×C.argus♂)[J].Journal of Huazhong Agricultural University,2011,30(4):488-493.
[7] 张 颖,刘晓勇,曲秋芝,等.达氏鳇、施氏鲟及其杂交种(施氏鲟♂×达氏鳇♀)形态差异与判别分析[J].淡水渔业,2012,42(6):27-32.
ZHANG Y,LIU X Y,QU Q Z,et al.Comparative discriminate analysis of morphological traits amongAcipenserschrenckii,Husodauricusand their hybrid(A.schrenckii♂×H.dauricus♀)[J].Freshwater Fisheries,2012,42(6):27-32.
[8] 马海涛,韩承慧,滕照军,等.大菱鲆(Scophthalmus maximus)、星斑川鲽(Platichthys stellatus)及其杂交种的形态学分析[J].海洋与湖沼,2014,45(6):1380-1386.
MA H T,HAN C H,TENG Z J,et al.Morphological analysis toScophthalmus maximus,Platichthys stellatus and their hybrid[J].Oceanologia Et Limnologia Sinica,2014,45(6):1380-1386.
[9] 李 冰,柴学森,张成锋,等.建鲤和黑龙江野鲤自交和正反交F1形态差异分析[J].中国水产科学,2014,21(1):59-66.
LIB,CHAIX S,ZHANG C F,et al.Morphometric variations analysis ofCyprinuscarpiovar.jian,Cyprinus carpio haermatopterus and the reciprocal F1hybrid(Cyprinus carpiovar.jian♀×Cyprinus carpio haermatopterus♂,Cyprinus carpiovar.jian♂×Cyprinus carpio haermatopterus♀)[J].Journal of Fishery Sciences of China,2014,21(1):59-66.
[10] 李雅娟,张明昭,于 卓,等.黑龙江泥鳅、北方泥鳅和泥鳅的形态差异分析[J].大连海洋大学学报,2010,25(5):397-401.
LIY J,ZHANG M Z,YU Z,et al.Morphological variations in Amur weather fishMisgurnus mohoity,northern weather fishMisgurnus bipartitusand oriental weather fishMisgurnus anguillicaudatusin China[J].Journal of Dalian Ocean University,2010,25(5):397-401.
[11] 霍堂斌,袁美云,马 波,等.白斑狗鱼与黑斑狗鱼的形态差异与判别分析[J].大连海洋大学学报,2011,26(3):253-259.
HUO T B,YUAN M Y,MA B,et al.Morphological variations and discrimination between northern pikeEsox Luciusand Amur pikeEsox reicherti[J].Journal of Dalian Ocean University,2011,26(3):253-259.
[12] 温海深,钱 焜,张美昭,等.花鲈繁殖生理及人工养殖模式探析[J].现代农业科技,2013(5):281-282.WEN H S,QIAN K,ZHANGM Z,etal.Discusses on the reproductive physiology and artificial culturing modes of spotted sea bass[J].Modern Agricultural Science and Technology,2013(5):281-282.
[13] 常 青,梁萌青,王家林,等.花鲈对不同饲料原料的表观消化率[J].水生生物学报,2005,29(2):172-176.
CHANG Q,LIANG M Q,WANG J L,et al.Apparent digestibility coefficients of various feed ingredients for Japanese sea bass(Lateolabrax japonicus)[J].Acta Hydrobiologica Sinica,2005,29(2):172-176.
[14] 张志勇,薛 敏,王 嘉,等.混合植物蛋白质替代鱼粉对花鲈和西伯利亚鲟生长和肉质影响的比较研究[J].动物营养学报,2013,25(6):1260-1275.
ZHANG Z Y,XUE M,WANG J,et al.Effects of fish meal replacement by plant protein blend on growth and flesh quality of Japanese sea bass(Lateolabrax japonicus)and Siberian sturgeon(Acipenserbaeri Brandt)[J].Chinese Journal of Animal Nutrition,2013,25(6):1260-1275.
[15] 王 艳,胡先成,罗 颖.盐度对鲈鱼稚鱼的生长及脂肪酸组成的影响[J].重庆师范大学学报(自然科学版),2007,24(2):62-66.
WANG Y,HU X C,LUO Y.Effects of salinity on growth and fatty acids composition of juvenileLateolabrax japonicus[J].Journal of Chongqing Normal University(Natural Science Edition),2007,24(2):62-66.
[16] 张春丹,李明云.花鲈繁殖生物学及繁育技术研究进展[J].宁波大学学报(理工版),2005,18(3):400-403.
ZHANG C D,LI M Y.Advance of study on reproductive biology and breeding technology ofLateolabrax japonicus[J].Journal of Ningbo University(NSEE),2005,18(3):400-403.
[17] 胡自民,高天翔,韩志强,等.花鲈和鲈鱼群体的遗传分化研究[J].中国海洋大学学报,2007,37(3):413-418.
HU ZM,GAO T X,HAN Z Q,et al.Studies on genetic differentiation of the spotted sea bass(Lateolabrax maculatus)and Japanese sea bass(Lateolabrax japonicus)[J].Periodical of Ocean University of China,2007,37(3):413-418.
[18] 刘明月,蒋琦辰,杨家新.不同海域中国花鲈的细胞色素b序列的遗传分析[J].南京师大学报(自然科学版),2010,33(1):102-106.
LIU M Y,JIANG Q C,YANG J X.Analysis on mitochondrial DNA cytochrome b gene ofLateolabrax japonicusfrom different seas[J].Journal of Nanjing Normal University(Natural Science Edition),2010,33(1):102-106.
[19] 江 鑫,杨官品,危起伟,等.中国沿海花鲈群体遗传结构分析[J].中国海洋大学学报,2009,39(2):271-274.
JIANG X,YANGG P,WEIQW,et al.Analysis of the genetic structure of spotted sea bass(Lateolabrax maculatus)inhabiting the Chinese coast[J].Periodical of Ocean University of China,2009,39(2):271-274.
[20] 张尧庭,方开泰.多元统计分析引论[M].北京:科学出版社,1982:393-401.
ZHANG Y T,FANG K T.Introduction ofmultivariate statistical analysis[M].Beijing:Science Press,1982:393-401.
[21] SLATKIN M.Gene flow in natural populations[J].Annual Review of Ecology and Systematics,1985(16):393-430.
[22] 韦成昱,刘西磊,杜玉兰,等.我国海鲈养殖现状及养殖技术分析[J].安徽农业科学,2015,43(28):132-133.
WEIC Y,LIU X L,DU Y L,et al.Status of breeding sea bass and breeding technical analysis[J].Journal of Anhui Agricultural Science,2015,43(28):132-133.
[23] 董双林,潘克厚,UWE B.海水养殖对沿岸生态环境影响的研究进展[J].青岛海洋大学学报,2000,30(4):575-582.
DONG SL,PAN K H,UWE B.Review on effects of mariculture on coastal environment[J].Journal of Ocean University of Qingdao,2000,30(4):575-582.
[24] 黎中宝,方 秀,陈 锦,等.大黄鱼(Pseudosciaena crocea)养殖群体遗传多样性的降低[J].海洋与湖沼,2009,40(4):446-450.
LI Z B,FANG X,CHEN J,et al.Loss of the genetic diversity in cultivated populations ofPseudosciaena croceaby AFLP[J].Oceanologia Et Limnologia Sinica,2009,40(4):446-450.
[25] CLIFFORD SL,McGINNITY P,FERGUSON A.Genetic changes in an Atlantic salmon population resulting from escaped juvenile farm salmon[J].Journal of Fish Biology,1998,52(1):118-127.
Morphological variations of six spotted sea bass(Lateolabrax maculatus)geographical populations along China coast
HOU Ji-lun1,ZHAO Ya-xian1,REN Jian-gong1,WANG Gui-xing1,CHEN Rui-yi2,LUO Ming-fei3,WANG Yu-fen1,LIU Hai-jin1
(1.Beidaihe Central Experiment Station,Chinese Academy of Fishery Sciences,Qinhuangdao066100;2.Marine Fisheries Research Institute of Zhejiang,Zhoushan 316021;3.Zhuhai Modern Agriculturaldevelopment center,Zhuhai519000)
Spotted sea bass(Lateolabrax maculatus)is an eurythermic fish inhabiting in the coastal lower layers,and widely distributed along the coast of China and the Korean peninsula.Spotted sea bass is euryhalinous,and can survive in fresh water as well as marine water.With the breakthroughs in key technologies such as artificial breeding and feeds,the culture of spotted sea bass has been rapidly developed.Recent years,many Chinese researchers have studied the nutrition,reproductive physiology and population genetics of spotted sea bass.Morphology is an important character of organisms.Through the measurement of morphological data,and corresponding statistical analysis,the similarities and differences between individuals or groups can be compared.The common method for analyzing morphological differences is the multivariate statistical analysis,which contains multiple variance analysis,cluster analysis,factor analysis,principal component analysis,discriminant analysis and others.Multivariate analysis is able to analyze the statistical regularity of the multiple indexes and the objects that are related to each other.In the study of fish,multivariate analysis is widely used in the comparison of morphological differences of different geographic populations,morphological differences between hybrids and their parents,and discrimination of varieties.However,the morphological variation analysis of different geographical populations has not been reported yet.In this paper,the morphological variations of six spotted sea bass geographical populations along China coast were analyzed using multivariate analysis with eight proportional characters.Cluster analysis and principal component analysis showed that these six populations could be divided into two groups:the northern group thatincluded Donggang,Qinhuangdao,Suizhong and Qingdao population,which were from the Yellow Sea and the Bohai Sea;the southern group that included Zhoushan(from the East China Sea)and Zhuhai population(from the South China Sea).There were a certain morphological differences between the north and the south group,while the relationship between Zhuhai and the northern population was closer than that between Zhoushan population and the northern group.Principal component analysis constructed 3 principal components with contributions of31.726%,27.744%and 14.075%,respectively,and the cumulative contribution rate was 73.545%.Finally,discriminant formulas were constructed by 8 variables to discriminate the six geographical populations.The discriminant accuracy varied from 63.6%to 84.4%,and synthetic discriminant accuracy rate was 72.7%.The results of this study provide basic data for the identification of geographical populations,evaluation of germplasm resources as well as the selection of fine variety.
spotted sea bass(Lateolabrax maculatus);multivariate analysis;morphological variations;escape
Q 145;S 917
A
1004-2490(2016)05-0459-09
2016-03-22
现代农业产业技术体系建设专项资金(CARS-50-G02,CARS-50-Z03)
侯吉伦(1984-),男,博士,浙江舟山人,助理研究员,研究方向:鱼类遗传育种。Tel:0335-4260806,E-mail:jilunhou@hotmail.com
刘海金,研究员。Tel:0335-4260860,E-mail:liuhaijin2005@126.com
