多年生黑麦草对Pb、Cd胁迫的响应及富集能力研究
2016-02-23冯鹏孙力申晓慧姜成李如来李增杰郑海燕
冯鹏,孙力,申晓慧,姜成,李如来,李增杰,郑海燕,
张华1,郭伟1,韩旭东1,洪亚南1
(1.黑龙江省农业科学院佳木斯分院,黑龙江 佳木斯 154007;2.黑龙江省农业科学院博士后工作站,
黑龙江 哈尔滨150086;3.佳木斯大学生命科学学院,黑龙江 佳木斯 154003)
多年生黑麦草对Pb、Cd胁迫的响应及富集能力研究
冯鹏1,2*,孙力1,申晓慧1,姜成3,李如来1,李增杰1,郑海燕1,
张华1,郭伟1,韩旭东1,洪亚南1
(1.黑龙江省农业科学院佳木斯分院,黑龙江 佳木斯 154007;2.黑龙江省农业科学院博士后工作站,
黑龙江 哈尔滨150086;3.佳木斯大学生命科学学院,黑龙江 佳木斯 154003)
摘要:研究了铅(Pb)、镉(Cd)及Pb-Cd复合胁迫对多年生黑麦草生长发育的影响,了解多年生黑麦草修复Pb、Cd污染土壤的潜能,为农田土壤的污染修复工作提供技术支撑和应用借鉴。以人工盆栽的方法,Pb设0, 300, 500, 1000, 1500 mg/kg 5个处理,Cd设0, 0.3, 3, 10, 50, 100 mg/kg 6个处理,Pb-Cd复合胁迫设Pb0Cd0, Pb300Cd0.3, Pb500Cd0.3, Pb1000Cd0.3, Pb1500Cd0.3, Pb300Cd10, Pb500Cd10, Pb1000Cd10, Pb1500Cd109个处理。测定Pb、Cd单一及复合胁迫下,多年生黑麦草种子发芽势、发芽率、发芽指数、简化活力指数,叶片叶绿素含量,净光合效率(Pn)、蒸腾速率(Tr)、光合有效辐射(PAR)、光能利用率(LUE),根、茎、叶各器官Pb、Cd含量,并计算各处理富集系数。结果表明,低浓度Pb离子(300,500 mg/kg)对多年生黑麦草种子发芽影响不显著,高浓度Pb离子(1000,1500 mg/kg)抑制种子发芽,低浓度Cd离子对种子发芽势及发芽率具有促进作用,Cd离子浓度超过10 mg/kg,则表现显著抑制种子发芽;Pb、Cd单一胁迫下,随Pb离子浓度的增加,叶片叶绿素含量呈递增趋势,随Cd离子浓度增加,叶绿素含量呈递减趋势。Pb离子浓度在0~1500 mg/kg范围,Cd离子浓度在0~100 mg/kg范围,Pb、Cd离子单一胁迫下,净光合效率均表现先降低,后升高的变化趋势,Pb浓度为1000 mg/kg时Pn达到最大值,为21.59 μmol CO2/(m2·s);Cd浓度为10 mg/kg时,Pn达到最大值,为22.67 μmol CO2/(m2·s);Pb、Cd单一胁迫处理,随Pb、Cd浓度的增加,多年生黑麦草富集系数表现降低趋势。Pb-Cd复合胁迫对种子发芽抑制作用大于Pb、Cd单一元素胁迫;Pb、Cd及Pb-Cd复合胁迫,多年生黑麦草根、茎、叶吸收Pb、Cd离子顺序均为根>叶>茎;多年生黑麦草对Cd离子吸收富集效应较Pb离子更为显著,对轻度Pb、Cd污染土壤修复效果更为明显。
关键词:铅;镉;胁迫;多年生黑麦草;富集能力
植物修复(phyto-remediation)是指直接或间接利用植物或植物组织培养物在原位去除或控制耕地土壤、大气水体沉积物、固体废弃物、地表水、地下水、污泥和大气等环境介质中的如重金属、类金属、放射性元素和有机污染物等有毒有害物质,从而最大限度地降低或消除其环境风险的环境友好修复技术[1]。具有修复成本低,操作技术简单;绿色环保;不造成“二次污染”;并可以提高土壤肥力,有利于后期土壤利用等优势[2]。
多年生黑麦草(Loliumperenne)为禾本科多年生草本植物,是一种多用途、抗性强的植物,它对贫瘠地、有机物、污水等都表现出较强的抗性。而且,多年生黑麦草在生境恶劣、寸草不生的尾矿地都能生存,甚至能正常生长[3]。通过对多年生黑麦草反复修剪刈割,可以逐步减少土壤中重金属离子含量,达到植物修复污染土壤的目的。侯伶龙等[4]研究得出,多年生黑麦草、大花月见草(Oenotherarosea)、早熟禾(Poaannua)、羽衣甘蓝(Brassicaoleracea)、曼陀罗(Daturastramonium)等对土壤Cd有较强的富集作用。Alvarenga等[5]发现多年生黑麦草因其大量的根系和分布广泛的生长模式可应用于植物固定。徐卫红等[6]研究指出,多年生黑麦草吸收的锌、镉主要集中在地上部,锌、镉复合污染植株对重金属离子的蓄集量最大。徐佩贤等[7]研究4种草坪植物对镉的耐受性与积累特性,结果表明,高羊茅(Festucaarundinacea)对Cd的耐受性最好,其次为多年生黑麦草,再次是草地早熟禾,匍匐剪股颖(Agrostisstolonifera)对Cd的耐受性最差;在相同Cd浓度处理下,多年生黑麦草地上部的Cd浓度最低,根系Cd浓度最高。
重金属对植物的毒害是深远的,从植物种子萌发、幼苗生长和成熟植株开花结实等都会有很强的毒害作用,致使植物的生长发育受到抑制[8]。李慧芳等[9]研究指出,随着镉浓度的增加,多年生黑麦草的相对存活率和相对地下生物量显著降低;相对株高、相对分蘖数和相对地上生物量呈先上升后下降的趋势。王晨等[10]研究发现,Cd 使黑麦草根系发育受阻导致生物量下降,Cd、Zn、Pb 复合污染造成黑麦草叶绿体结构的破坏,使叶绿素含量减少。陈伟等[11]在研究Cd2+、Cu2+、Zn2+、Pb2+对多年生黑麦草荧光特性的影响时得出,Cd2+胁迫对多年生黑麦草PSⅡ系统的活性影响不大;多年生黑麦草表现出低浓度Zn2+促进PSⅡ系统活性的作用,但随着浓度升高,表现出对PSⅡ系统越来越强的抑制作用。尽管重金属胁迫对植物生长发育的研究已有报道,但有关多年生黑麦草修复Pb、Cd及Pb-Cd复合污染土壤报道仍较少。本文通过分析Pb、Cd及Pb-Cd复合胁迫对多年生黑麦草生长发育的影响,找出多年生黑麦草对Pb、Cd污染的临界抗性浓度,并分析多年生黑麦草对Pb、Cd离子吸收积累特性,深入了解修复Pb、Cd污染土壤的潜力,为植物修复重金属污染土壤提供理论依据。
1材料与方法
1.1材料
供试材料为多年生黑麦草“匹克威”品种种子,由中国林业科学院提供。供试金属离子:Pb(NO3)2、CdCl2·2.5H2O均为分析纯试剂。盆栽用土为草甸黑土,有机质含量为2.49%(Walkley-Black method);碱解氮含量86.3 mg/kg,有效磷含量64.6 mg/kg,速效钾含量79.9 mg/kg,全氮含量0.14%,全磷含量0.14%,全钾含量3.12%,pH 6.5;土壤Pb,Cd背景值分别为0.24 mg/kg,0.031 mg/kg。
1.2实验设计
2014年5月1日按照牧草检验规程标准方法[12]进行发芽试验。每天更换Pb,Cd离子培养液,以保证重金属离子胁迫浓度一致。将实验室发芽的种子培育成苗,6月10日移栽于相同Pb,Cd浓度塑料盆中,规格为34 cm×21 cm×27 cm(上径×下径×高),每盆装土10 kg,平衡1个月后,供移栽。以灌蒸馏水为对照(CK),每个处理3盆,每盆50株。9月1日测定植株光合指标,并取样测定叶片叶绿素含量及植株根、茎、叶Pb、Cd含量。
1.3Pb、Cd及Pb-Cd复合胁迫处理设置
Pb离子浓度设置0,300,500,1000,1500 mg/kg 5个处理,Cd离子浓度设置0,0.3,3,10,50,100 mg/kg 6个处理,Pb-Cd复合胁迫设置Pb0Cd0,Pb300Cd0.3,Pb500Cd0.3,Pb1000Cd0.3,Pb1500Cd0.3,Pb300Cd10,Pb500Cd10,Pb1000Cd10,Pb1500Cd109个处理(下标即为对应Pb,Cd离子浓度)。
1.4方法
1.4.1标准发芽率、发芽势、发芽指数和活力指数的测定采用培养皿纸上发芽,4℃预冷7 d后移入发芽箱,每日光照8 h,温度30℃,黑暗16 h,温度20℃,第5天初次计数,末次计数在第14天,共18个处理,重复4次,各100粒种子;从种子萌发开始计数到第14天为止,种苗长度在第14天测试;计算种子发芽势、发芽率、发芽指数和简化活力指数[13]。
发芽势(%)=(前5 d正常种苗数/供试种子数)×100
发芽率(%)=(前14 d正常种苗数/供试种子数)×100
发芽指数GI=∑(Gt/Dt)
简化活力指数=G×S
式中,Gt为t日的发芽数,Dt为相应的发芽日数,G为发芽率,S为萌发第14天种苗长。
1.4.2叶绿素含量测定取植株中部成熟叶片0.2 g。80%丙酮+95%乙醇20 mL水浴,50℃避光浸提24 h[14], 645和663 nm波长处测定光密度值,以提取液为空白溶液。设重复3次。叶绿素含量用 Arnon法[12]的修正公式:
Ca=(12.71A663-2.59A645)×(V/M)×1000
Cb=(22.88A645-4.67A663)×(V/M)×1000
Ct=(8.04A663+20.29A645)×(V/M)×1000
式中,Ca、Cb、Ct分别为叶绿素a、b及叶绿素总量浓度;V为提取液体积;M为称取叶片质量;A645、A663分别为645和663 nm吸光度值[12]。
1.4.3光合特性测定用美国CID公司生产的CI-310便携式光合作用分析仪在开路系统下测定各处理植株瞬时净光合速率(Pn)、蒸腾速率(Tr)以及光合有效辐射(PAR)。测定标准为:每一处理测定2株,每一植株分别测定上、中、下部向阳面成熟叶片。叶室为12 cm×12 cm×25 cm。测定在晴天进行。光能利用效率(LUE)=Pn/PAR[15]。
1.4.4植株根、茎、叶Pb、Cd含量测定取各处理植株根、茎、叶,烘干并粉碎,过20目(0.074 mm)筛;称取1.00 g试样于坩埚中,电炉炭化无烟后,移入马弗炉500℃灰化6 h,冷却。0.5 mol/L硝酸溶解,定容25 mL。用日立Z-2000原子吸收分光光度计,石墨炉原子化法测定植株各部分Pb、Cd含量[16-17]。
仪器条件:测定波长Pb、Cd分别为283.3和228.8 nm,狭缝均为1.0 nm,灯电流分别为7和10 mA;干燥温度120℃,20 s;灰化温度分别为450和350℃,20 s;原子化温度均为2300℃,5 s。
1.4.5富集系数富集系数(enrichment coefficient,EC)是指植物体内某种重金属含量与土壤中该种重金属含量的比值,是评价植物富集重金属能力的重要指标之一,反映植物对某种重金属元素的富集能力[18]。
计算公式:EC=a/b,式中,a为植物体内重金属元素含量,单位mg/kg;b为土壤中的重金属元素含量,单位mg/kg。
1.5数据分析
采用SAS 8.1 (SAS Institute Inc.,2006) 统计软件,在P<0.05水平对试验结果进行方差分析和Duncan多重比较。
2结果与分析
2.1Pb、Cd及Pb-Cd复合胁迫对种子发芽特性的影响
多年生黑麦草种子发芽势,随Pb浓度的增加,表现递减趋势,但各处理与对照差异不显著(表1);高浓度Pb对发芽率影响较明显,1000,1500 mg/kg Pb浓度处理发芽率最低,分别为83.68%,84.14%,与对照差异显著(P<0.05),说明重金属Pb降低多年生黑麦草种子发芽能力,抑制种子发芽。Cd离子对多年生黑麦草种子发芽势及发芽率表现单峰曲线,低浓度Cd处理(0.3,3 mg/kg)组对种子发芽势及发芽率具有促进作用,当Cd浓度超过10 mg/kg,则显著抑制种子发芽。各处理简化活力指数均低于对照,说明Pb、Cd胁迫降低种子发芽活力。Pb-Cd复合胁迫,同浓度Cd处理组,随Pb浓度增加,多年生黑麦草种子发芽率及发芽势均表现下降趋势,且均低于同浓度Cd单一胁迫,说明Pb-Cd复合胁迫对种子发芽抑制作用大于单一Cd离子胁迫。
表中同列不同字母表示差异显著(P<0.05)。下同。
Means in the same column with different letters differ significantly (P<0.05). The same below.
2.2 Pb、Cd及Pb-Cd复合胁迫对植株叶绿素含量的影响
Pb、Cd及Pb-Cd复合胁迫对多年生黑麦草叶绿素a、叶绿素b、叶绿素总量影响趋势基本相同(表2)。各处理叶绿素含量均显著高于对照(P<0.05),说明重金属离子促进植株叶片叶绿素合成,叶绿素含量增加。Pb、Cd单一胁迫下,随Pb离子浓度的增加,叶片叶绿素含量呈递增趋势;随Cd离子浓度增加,叶片叶绿素含量呈递减趋势。Pb-Cd复合胁迫下,同浓度Cd离子处理组,叶片叶绿素含量均表现先增加后降低趋势,Pb离子浓度为1000mg/kg、Cd离子浓度为10mg/kg处理最大(P<0.05),叶绿素a、叶绿素b、叶绿素总量分别为1.381,0.511,1.892mg/g。
2.3 Pb、Cd及Pb-Cd复合胁迫对植株光合特性的影响
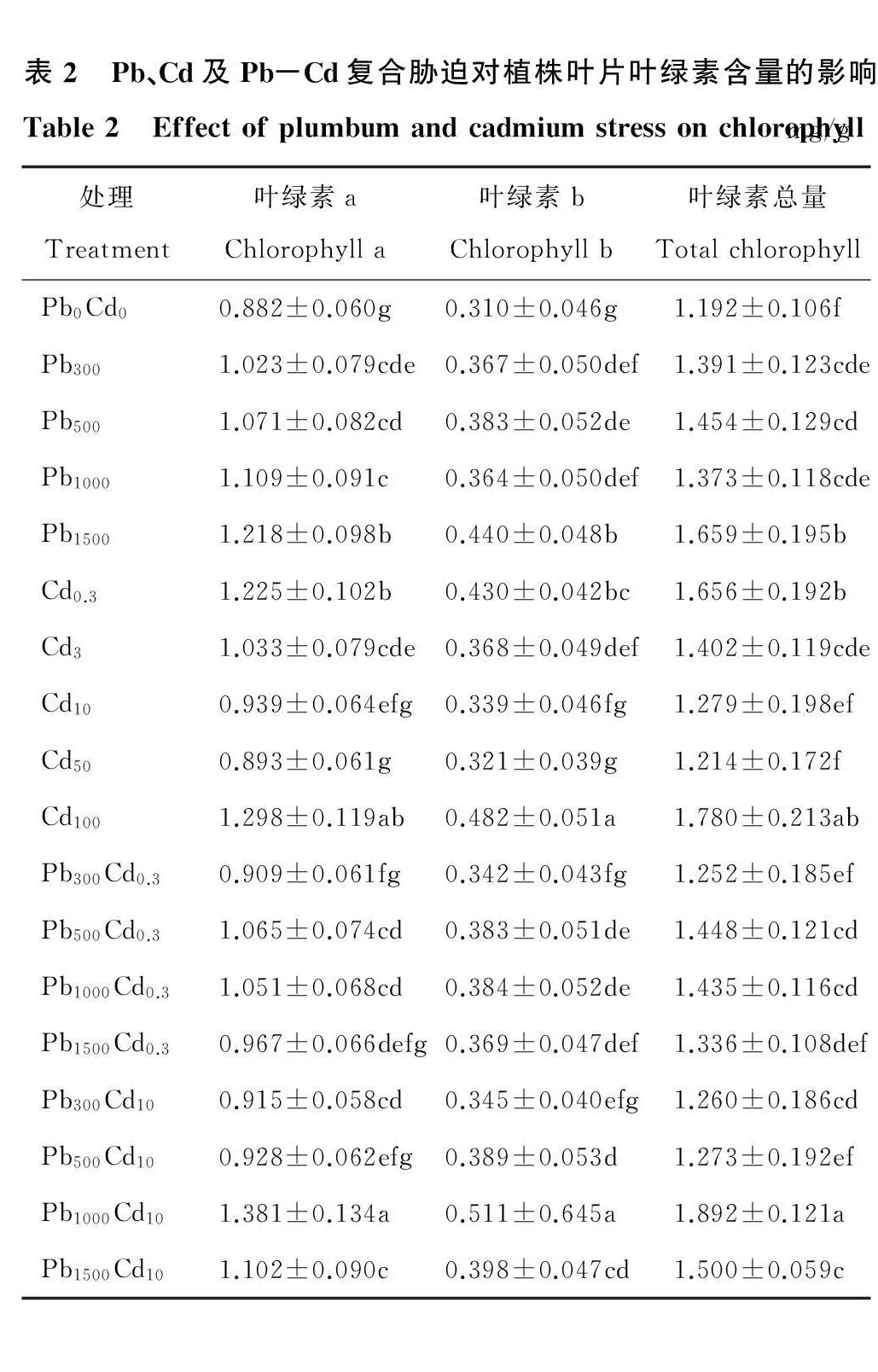
表2 Pb、Cd及Pb-Cd复合胁迫对植株叶片叶绿素含量的影响Table2 Effectofplumbumandcadmiumstressonchlorophyllmg/g处理Treatment叶绿素aChlorophylla叶绿素bChlorophyllb叶绿素总量TotalchlorophyllPb0Cd00.882±0.060g0.310±0.046g1.192±0.106fPb3001.023±0.079cde0.367±0.050def1.391±0.123cdePb5001.071±0.082cd0.383±0.052de1.454±0.129cdPb10001.109±0.091c0.364±0.050def1.373±0.118cdePb15001.218±0.098b0.440±0.048b1.659±0.195bCd0.31.225±0.102b0.430±0.042bc1.656±0.192bCd31.033±0.079cde0.368±0.049def1.402±0.119cdeCd100.939±0.064efg0.339±0.046fg1.279±0.198efCd500.893±0.061g0.321±0.039g1.214±0.172fCd1001.298±0.119ab0.482±0.051a1.780±0.213abPb300Cd0.30.909±0.061fg0.342±0.043fg1.252±0.185efPb500Cd0.31.065±0.074cd0.383±0.051de1.448±0.121cdPb1000Cd0.31.051±0.068cd0.384±0.052de1.435±0.116cdPb1500Cd0.30.967±0.066defg0.369±0.047def1.336±0.108defPb300Cd100.915±0.058cd0.345±0.040efg1.260±0.186cdPb500Cd100.928±0.062efg0.389±0.053d1.273±0.192efPb1000Cd101.381±0.134a0.511±0.645a1.892±0.121aPb1500Cd101.102±0.090c0.398±0.047cd1.500±0.059c
净光合效率、蒸腾速率、光合有效辐射是植株光合特性的主要指标。结果表明,单一Pb、Cd离子胁迫下,净光合效率均表现先降低,后升高的变化趋势(表3),Pb浓度为1000mg/kg时Pn达到最大值,为21.59μmolCO2/(m2·s),Cd浓度为10mg/kg时,Pn达到最大值,为22.67μmolCO2/(m2·s)(P<0.05);Pb-Cd复合胁迫,低浓度Cd离子(0.3mg/kg)与Pb复合胁迫时,随Pb浓度增加,Pn先升后降;高浓度Cd离子(10 mg/kg)与Pb复合胁迫,随Pb浓度增加,Pn明显持续降低,说明较高浓度Cd离子处理对多年生黑麦草净光合效率影响更为显著。Pb、Cd及Pb-Cd复合胁迫对各处理蒸腾速率、光合有效辐射影响趋势基本相同,随重金属离子Pb、Cd浓度增加,叶片Tr、PAR均表现先升高后降低趋势。
2.4多年生黑麦草根、茎、叶Pb、Cd吸收积累特性
Pb、Cd单一胁迫多年生黑麦草根、茎、叶均表现相同趋势,随着Pb、Cd浓度的增加,植株根、茎、叶Pb、Cd离子含量增加(表4);Pb-Cd复合胁迫下,Cd离子(0.3 mg/kg)处理组,多年生黑麦草根系Cd离子含量显著高于同浓度Cd离子单一胁迫根系Cd离子含量(P<0.05),说明Pb离子存在促进植株根系对Cd离子吸收;Pb-Cd复合胁迫下,Cd离子浓度0.3 mg/kg处理组植株根、茎、叶Pb离子显著高于Cd离子浓度为10 mg/kg处理组,说明高浓度Cd离子对植株根、茎、叶Pb离子吸收具有一定的抑制作用。Pb、Cd单一胁迫及Pb、Cd复合胁迫,多年生黑麦草根、茎、叶吸收Pb离子能力顺序均为根>叶>茎。
2.5多年生黑麦草对Pb、Cd离子生物富集效应
为了反映植物对重金属离子的富集能力,Chamber-lain定义富集系数为植物体中的污染物含量与生长基质中该污染物含量的比值;富集系数越大,表明植物对该金属元素的吸收能力越强[18]。Pb、Cd单一胁迫处理,随Pb、Cd浓度的增加,多年生黑麦草富集系数表现降低趋势(表5),说明多年生黑麦草对低浓度Pb、Cd污染土壤富集效应大于高浓度处理。Pb-Cd复合胁迫,低浓度Cd处理组(0.3 mg/kg)富集系数均显著高于高浓度Cd(10 mg/kg)(P<0.05),说明Pb-Cd复合胁迫下,Pb离子促进多年生黑麦草对低浓度Cd离子富集效应。由表5可知,Pb、Cd及Pb-Cd复合胁迫下,多年生黑麦草对Cd离子富集系数均大于Pb离子富集系数,表明多年生黑麦草对Cd离子吸收富集效应较Pb离子更为显著。
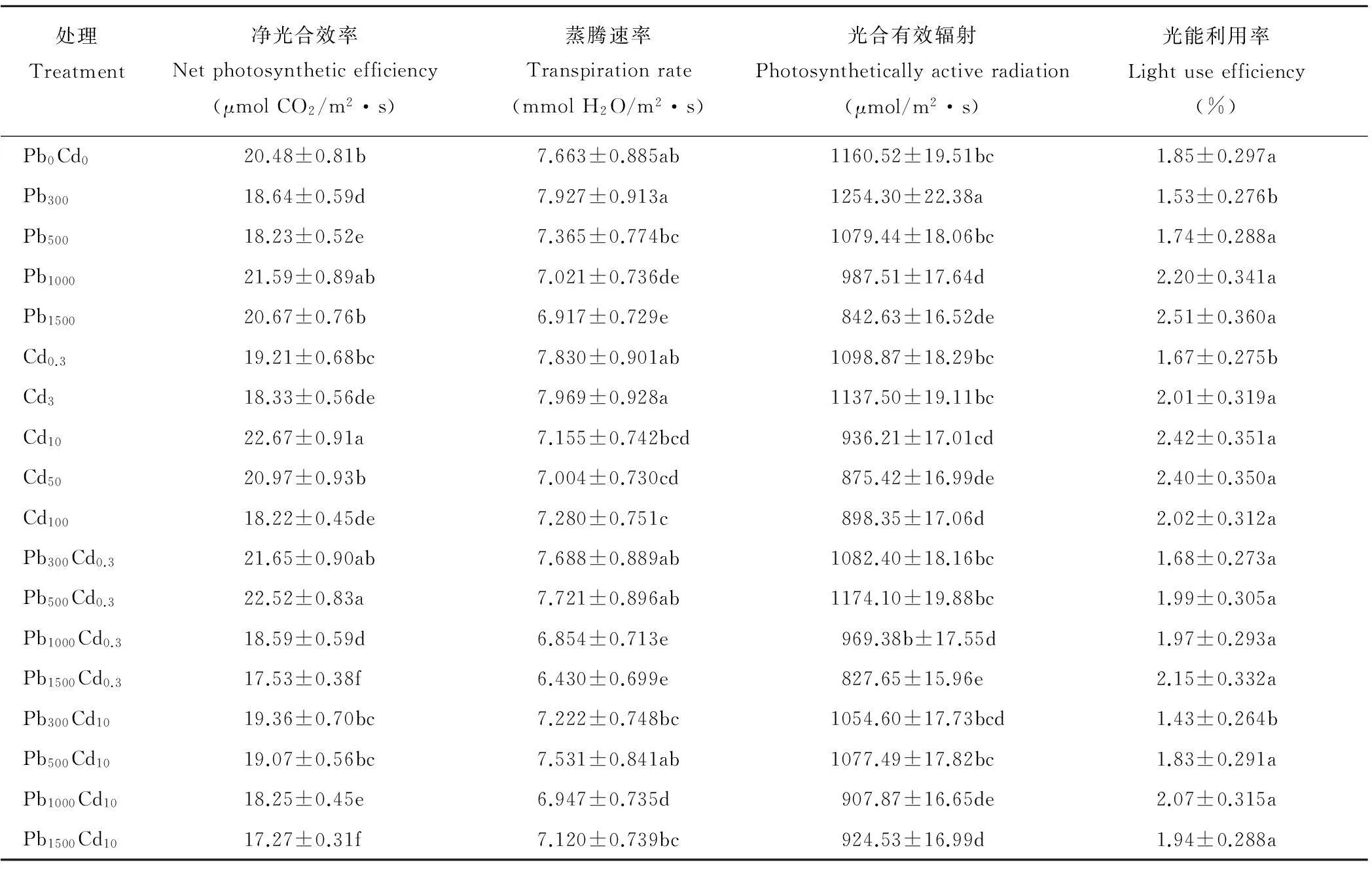
表3 Pb、Cd及Pb-Cd复合胁迫对植株光合特性的影响

表4 多年生黑麦草根、茎、叶Pb、Cd吸收积累特性

表5 多年生黑麦草对重金属Pb、Cd离子生物富集效应
3讨论
铅、镉是农田土壤中主要的重金属污染物,进入植物体内并积累到一定程度后,严重影响植物的生长发育,甚至枯萎死亡[19]。刘明美等[20]研究发现,Pb2+处理浓度<800 mg/kg时, 多花黑麦草种子的萌发和幼苗生长受到促进, 出现增效效应;大于800 mg/kg, 种子萌发及幼苗生长受到显著抑制。高浓度Pb2+处理对根生长的抑制大于对芽生长的抑制。王锦文等[21]研究了不同浓度铅、镉对水稻(Oryzasative)种子发芽、幼苗生长的影响,结果表明,铅、镉胁迫下,水稻发芽率降低,芽和根的伸长受到抑制,尤其是根。王丽燕和郑世英[22]通过水培实验发现低浓度的镉、铅处理对小麦(Triticumaestivum)种子发芽率、发芽势、发芽指数、活力指数及根长均有一定的促进作用,随着处理浓度的提高其数值逐渐下降。本研究中,低浓度Pb(300,500 mg/kg)处理对多年生黑麦草种子发芽影响不显著;低浓度Cd离子对种子发芽势及发芽率具有促进作用,Cd离子浓度超过10 mg/kg,则表现显著抑制种子发芽。这与刘明美等[20]、王丽燕和郑世英[22]的研究结果相一致。造成这一现象的原因可能是低浓度的Pb、Cd离子可提高胚的生理活性,促进萌发;而高浓度Pb、Cd离子对胚、芽等产生了伤害作用,并且高浓度Pb、Cd胁迫抑制淀粉酶、蛋白酶活性,即抑制种子内贮藏淀粉和蛋白质的分解,从而影响种子萌发所需要的物质和能力,致使种子萌发受到抑制[23]。
植物受重金属毒害的外部症状表现为叶色黄化,甚至枯萎死亡[24]。夏红霞等[25]研究了重金属铅对黑麦草叶绿素荧光特性的影响,结果表明,低浓度铅(<200 mg/kg)可促进黑麦草叶绿素的合成,促进植株的光合作用;高浓度铅(200~3000 mg/kg)抑制叶绿素的合成,降低植株光合作用,抑制植株生长。周朝彬等[26]对铅胁迫下草木樨(Melilotusofficinalis)叶中叶绿素含量及光合特性进行研究,表明铅胁迫下叶绿素含量、气孔导度和光合速率变化明显,铅处理浓度与叶绿素含量变化呈明显正相关。Drziewicz和Baszynski[27]研究结果表明Pb、Cd 处理可使叶绿体结构发生明显变化,破坏了叶绿体的膜系统。本实验中Pb、Cd离子胁迫下,各处理叶片叶绿素含量均高于对照,这与周朝彬等[26]的研究结果相似,但与夏红霞等[25]研究结果不同,可能因为本试验Pb离子浓度最高值为1000 mg/kg所限,Pb离子浓度>1000 mg/kg叶片叶绿素含量变化趋势,有待进一步研究证实。我们还发现Pb-Cd复合胁迫下,低浓度Cd离子(0.3 mg/kg)与Pb复合胁迫,随Pb浓度增加,Pn先升高后降低,高浓度Cd离子(10 mg/kg)与Pb复合胁迫,随Pb浓度增加,Pn明显持续降低,这说明较高浓度Cd离子对多年生黑麦草净光合效率影响更为显著。
植物体内不同部位重金属含量不同[28]。廖敏和黄昌勇[29]研究发现,根系中 Cd含量明显高于茎叶中 Cd 含量,进而得出,黑麦草根系对 Cd 有较强的富集作用,并阻止 Cd 向茎叶中迁移。杨明琰等[30]研究了铅胁迫对黑麦草Pb富集特性,结果表明,黑麦草不同部位对铅的富集规律表现为根>茎>叶,黑麦草根、茎、叶中的铅含量均随土壤铅含量的增大而显著增加,并呈明显的剂量效应关系。张尧等[31]研究指出黑麦草在Cd 浓度为10,20 mg/kg时,富集系数>1,对Cd 有较强的富集能力,转移系数<1,富集的Cd 主要累积在根部,表明黑麦草能够有效富集土壤中的Cd,且富集的Cd 主要积累在根部;根、茎、叶部分Cd 积累量:根>茎>叶。本实验得出Pb、Cd及Pb-Cd复合胁迫,多年生黑麦草根、茎、叶吸收重金属Pb、Cd离子能力顺序均为根>叶>茎的结论与此相一致。多年生黑麦草根系生物量大,分蘖再生能力强,生长速度快,同时能深植土壤,对重金属离子具有较好的富集能力。
由于黑麦草具有先锋植物特性和可以多次刈割并再生的特点,可以在对污染土壤的治理,恢复土壤再利用过程中发挥重要作用。且黑麦草对重金属具有很强的抗性,对重金属有蓄集作用[32]。李松克等[33]研究了黄壤种植多年生黑麦草土壤和植株重金属As、Hg、Pb、Cr、Cd含量的变化,结果表明,多年生黑麦草从根部向地上部运输重金属的能力顺序为Cd>Cr>As>Pb>Hg,地上部对重金属的富集能力顺序为Cd>Cr>Pb>As>Hg,对Cr、Pb、As、Hg的富集能力弱,对Cd为中等富集能力。杨卓等[34]研究了高羊茅(Festucaelata)、黑麦草对复合污染土壤Cd、Pb、Zn的富集特点,结果表明,其重金属的富集能力顺序为:Zn>Cd>Pb,本研究得出多年生黑麦草对Cd离子吸收富集效应较Pb离子更为显著的结论与此相似。同时,我们还发现Pb、Cd单一胁迫处理,随Pb、Cd浓度的增加,多年生黑麦草富集系数表现降低趋势,说明多年生黑麦草对低浓度Pb、Cd污染土壤富集效应大于高浓度处理,进一步表明多年生黑麦草对轻度Pb、Cd污染土壤修复效果更为明显。
4结论
低浓度Pb(<500 mg/kg)、Cd(<10 mg/kg)离子胁迫对多年生黑麦草种子发芽势及发芽率具有促进作用,高浓度则抑制种子发芽;Pb-Cd复合胁迫对种子发芽抑制作用大于Pb、Cd单一离子胁迫;Pb、Cd单一胁迫下,随Pb离子浓度的增加,叶片叶绿素含量呈递增趋势,随Cd离子浓度增加,叶绿素含量呈递减趋势。Pb离子浓度在0~1500 mg/kg范围,Cd离子浓度在0~100 mg/kg范围,Pb、Cd离子单一胁迫下,净光合效率均表现先降低,后升高的变化趋势;Pb、Cd及Pb-Cd复合胁迫下,多年生黑麦草根、茎、叶吸收重金属Pb、Cd离子能力顺序均为根>叶>茎;多年生黑麦草对Cd离子吸收富集效应较Pb离子更为显著,对轻度Pb、Cd污染土壤修复效果更为明显。
References:
[1]Rengasamy B, Pauline M. Organic acid complexation, heavy metal distribution and the effect of ATPase inhibition in hairy roots of hyperaccumulator plant species. Journal of Biotechnology, 2003, 101: 131-146.
[2]Shariful I, Takeshi S, Masaaki K. Phytofiltration of arsenic and cadmium by using an aquatic plant,Micranthemumumbrosum: Phytotoxicity, uptake kinetics, and mechanism. Ecotoxicology and Environmental Safety, 2015, 112: 193-200.
[3]Wu Q L, Wang W C, He S Y. Enhancement of GA3 and EDTA onLoliumperenneto remediate Pb contaminated soil and its detoxification mechanism. Chinese Journal of Applied Ecology, 2014, 25(10): 2999-3005.
[4]Hou L L, Huang R, Zhou L R,etal. The accumulation of Cd inHuttuyniacordataand its promotion by rhizospheric microbes. Ecology and Environmental Sciences, 2010, 19(4): 817-821.
[5]Alvarenga P, Gonalves A P, Fernandes R M. Evaluation of composts and liming materials the phytostabilization of a mine sio using perennial ryegrass. Science of the Total Environment, 2008, 406: 43-56.
[6]Xu W H, Wang H X, Wang Z Y,etal. Effects of zinc, cadmium and their combined pollution on nutrient uptake and Zn, Cd, accumulation in ryegrass (LoliumperenneL.). Asian Journal of Eeotoxieology, 2006, 11(1): 70-74.
[7]Xu P X, Fei L, Chen X B,etal. Cadmium tolerance and accumulation in four cool-season turfgrass. Acta Prataculturae Sinica, 2014, 23(6): 176-188.
[8]Sun R L, Jin C X, Zhou Q X. Characteristics of cadmium accumulation and tolerance inRorippaspecies with some characteristics of cadmium hyperaccumulation. Plant Growth Regulation, 2010, 67: 67-74.
[9]Li H F, Yuan Q H, Zhao G Q. Effects of cadmium stress on seeding growth of perennial ryegrass germplasm. Pratacultural Science, 2014, 31(5): 898-904.
[10]Wang C, Wang H Y, Zhao K,etal. Effects of silicon on physiological and biochemical properties of ryegrass under the compound pollution of Cd, Zn and Pb. Ecology and Environment, 2008, 17(6): 2240-2245.
[11]Chen W, Zhang M M, Song Y Y,etal. Impacts of heavy metals on the fluorescence characteristics and root morphology of 2 turfgrass species. Acta Prataculturae Sinica, 2014, 23(3): 333-342.
[12]Xu B M. The Seedling Evaluation and Vigour Test[M]. Beijing: China Agricultural University Press, 2001: 53-68.
[13]Chen W, Zhang M M, Song Y Y,etal. Inhibitory effect of heavy metal stress on the seed germination of four turfgrass types. Acta Prataculturae Sinica, 2013, 21(3): 556-563.
[14]Zhang F, Wan X Q, Zhai J. Effects of nitrogen supplement on chlorophyll synthesis and chloroplast ultrastructure of poplar plants under cadmium stress. Journal of Nuclear Agricultural Sciences, 2014, 28(3): 485-491.
[15]Zhang X H, Lang D Y, Zhang E H,etal. Diurnal changes in photosynthesis and antioxidants ofAngelicasinensisas influenced by cropping systems. Photosynthetica, 2013, 51(2): 252-258.
[16]Chinese Ministry of Health. GB 5009.12-2010. Determination of Pb in Foods[S]. Beijing: China Standard Press, 2010.
[17]Chinese Ministry of Health. GB/T 5009.15-2003. Determination of Cd in Foods[S]. Beijing: China Standard Press, 2003.
[18]Jian F M, Naoki Y J, Namki M,etal. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proceedings of the National Academy of the Sciences of the United States of America, 2008, 105: 9931-9935.
[19]Saraswat S, Rai J P. Phytoextraction potential of six plant species grown in multimetal contaminated soil. Chemistry and Ecology, 2009, 25(1): 1-11.
[20]Liu M M, Li J N, Shen Y X. Effects of Pb2+pollution on the seed germination and seedling growth of Italian ryegrass. Pratacultural Science, 2007, 24(1): 52-56.
[21]Wang J W, Bian C M, Chen Z. Effect of plumbum and cadmium stress on germination, growth and physiological of rice seedling. Jiangsu Agricultural Sciences, 2009, 4: 77-79.
[22]Wang L Y, Zheng S Y. Effect of cadmium, lead and their combined pollution on seed germination of wheat. Journal of Triticeae Crops, 2009, 29(1): 146-148.
[23]Ge C J, Chen Q B, Yu H M,etal. Effect of Cd on germination and inhibition of root elongation of tropical forage plants. Chinese Journal of Tropical Crops, 2008, 29(5): 567-571.
[24]Wu Y, Mu L Q. Effect of soil Pb, Cd stress on the growth, physiological and accumulation characteristics of four ornamental trees. Journal of Soil and Water Conservation, 2013, 27(5): 234-240.
[25]Xia H X, Zhu Q H, He C. Response of chlorophyll fluorescence characteristics ofLoliumperenneto Pb stress. Guizhou Agriculture Science, 2012, 40(12): 33-35.
[26]Zhou C B, Hu T X, Xu X G,etal. Effect of lead stress on chlorophyll content and photosynthetic characters in leaf of melilotus suavena. Journal of Sichuan Agricultural University, 2005, 23(4): 432-435.
[27]Drziewicz M, Baszynski T. Growth parameters and photosynthetic pigments in leaf segments ofZeamaysexposed to cadmium, as related to protect on mechanisms. Journal of Plant Physiology, 2005, 162: 1013-1021.
[28]Matos M, Rammalho J. Photosynthetic activity and cessular integrity of the Andean legumePachyrhizusahipaparody under heat and water stress. Photosynthetica, 2002, 40: 493-501.
[29]Liao M, Huang C Y. Effects of organic acids on the toxicity of cadmium during ryegrass growth. Chinese Journal of Applied Ecology, 2002, 13(1): 109-112.
[30]Yang M Y, Liang Y Y, Zen D B,etal. Effect of lead stress on accumulation capacity and physiological metabolism of ryegrass. Journal of Northwest Agriculture and Forestry University, 2014, 42(12): 97-102.
[31]Zhang R, Tian Z G, Cao C L,etal. Characteristics of cadmium tolerance, accumulation and cellular distribution inLoliumperenneseedlings. Journal of Agro-Environment Science, 2010, 29(11): 2080-2086.
[32]Zhang X, Lin A J, Zhao F J,etal. Arsenic accumulation by the aquatic fern Azolla: Comparison of arsenate uptake, speciation and efflux byA.carolinianaandA.filiculoides. Environmental Pollution, 2008, 156: 1149-1155.
[33]Li S K, Zhang C L, Li K Q,etal. Repairation ofLoliumperenneon yellow soil heavy metal pollution. Guizhou Agricultural Sciences, 2014, 42(11): 147-151.
[34]Yang Z, Wang W, Li B W,etal. Study on characteristics ofLoliummultiflorumandFestucaarundinaceaabsorbing and accumulating cadmium, lead and zinc from contaminated soil with these metals. Journal of Soil and Water Conservation, 2008, 22(2): 83-88.
参考文献:
[3]吴秋玲, 王文初, 何闪英. GA3与EDTA 强化黑麦草修复Pb 污染土壤及其解毒机制. 应用生态学报, 2014, 25(10): 2999-3005.
[4]侯伶龙, 黄荣, 周丽蓉, 等. 鱼腥草对土壤中镉的富集及根系微生物的促进作用. 生态环境学报, 2010, 19(4): 817-821.
[6]徐卫红, 王宏信, 王正银, 等. 锌、镉复合污染对重金属蓄集植物黑麦草养分吸收及锌、镉积累的影响. 生态毒理学报, 2006, 11(1): 70-74.
[7]徐佩贤, 费凌, 陈旭兵, 等. 四种冷季型草坪植物对镉的耐受性与积累特性. 草业学报, 2014, 23(6): 176-188.
[9]李慧芳, 袁庆华, 赵桂琴. 镉胁迫对多年生黑麦草种质幼苗生长的影响. 草业科学, 2014, 31(5): 898-904.
[10]王晨, 王海燕, 赵琨, 等. 硅对镉、锌、铅复合污染土壤中黑麦草生理生化性质的影响. 生态环境, 2008, 17(6): 2240-2245.
[11]陈伟, 张苗苗, 宋阳阳, 等. 重金属离子对2种草坪草荧光特性及根系形态的影响. 草业学报, 2014, 23(3): 333-342.
[12]徐本美. 种苗评定与种子活力测定方法手册(国际种子检验协会)[M]. 北京:中国农业大学出版社, 2001: 53-68.
[13]陈伟, 张苗苗, 宋阳阳, 等. 重金属胁迫对4种草坪草种子萌发的影响. 草业学报, 2013, 21(3): 556-563.
[14]张帆, 万雪琴, 翟晶. 镉处理下增施氮对杨树叶绿素合成和叶绿体超微结构的影响. 核农学报, 2014, 28(3): 485-491.
[16]中华人民共和国卫生部. GB 5009.12-2010. 食品中铅的测定[S]. 北京:中国标准出版社, 2010.
[17]中华人民共和国卫生部. GB/T 5009.15-2003. 食品中镉的测定[S].北京:中国标准出版社, 2003.
[20]刘明美, 李建农, 沈益新. Pb2+污染对多花黑麦草种子萌发及幼苗生长的影响. 草业科学, 2007, 24(1): 52-56.
[21]王锦文, 边才苗, 陈珍. 铅、镉胁迫对水稻种子萌发、幼苗生长及生理指标的影响. 江苏农业科学, 2009, 4: 77-79.
[22]王丽燕, 郑世英. 镉、铅及其复合污染对小麦种子萌发的影响. 麦类作物学报, 2009, 29(1): 146-148.
[23]葛成军, 陈秋波, 俞花美, 等. Cd胁迫对2种热带牧草种子发芽与根伸长的抑制效应. 热带作物学报, 2008,29(5): 567-571.
[24]温瑀, 穆立蔷. 土壤铅、镉胁迫对4种绿化植物生长、生理及积累特性的影响. 水土保持学报, 2013, 27(5): 234-240.
[25]夏红霞, 朱启红, 何超. 黑麦草叶绿素荧光特性对Pb离子胁迫的响应. 贵州农业科学, 2012, 40(12): 33-35.
[26]周朝彬, 胡庭兴, 胥晓刚, 等. 铅胁迫对草木樨中叶绿素含量和几种光合特性的影响. 四川农业大学学报, 2005, 23(4): 432-435.
[29]廖敏, 黄昌勇. 黑麦草生长过程中有机酸对镉毒性的影响. 应用生态学报, 2002, 13(1): 109-112.
[30]杨明琰, 梁语燕, 曾德榜, 等. 铅胁迫对黑麦草Pb富集特性及生理代谢的影响. 西北农林科技大学学报(自然科学版), 2014, 42(12): 97-102.
[31]张尧, 田正贵, 曹翠玲, 等. 黑麦草幼苗对镉耐性能力及吸收积累和细胞分布特点研究. 农业环境科学学报, 2010, 29(11): 2080-2086.
[33]李松克, 张春林, 李克勤, 等. 多年生黑麦草对黄壤重金属污染的修复. 贵州农业科学, 2014, 42(11): 147-151.
[34]杨卓, 王伟, 李博文, 等. 高羊茅和黑麦草对污染土壤Cd, Pb, Zn 的富集特征. 水土保持学报, 2008, 22(2): 83-88.
*Response and enrichment ability of perennial ryegrass under lead and cadmium stresses
FENG Peng1,2*, SUN Li1, SHEN Xiao-Hui1, JIANG Cheng3, LI Ru-Lai1, LI Zeng-Jie1, ZHENG Hai-Yan1, ZHANG Hua1, GUO Wei1, HAN Xu-Dong1, HONG Ya-Nan1
1.JiamusiBranchofHeilongjiangAcademyofAgriculturalSciences,Jiamusi154007,China; 2.PostdoctoralWorkstationofHeilongjiangAcademyofAgriculturalSciences,Herbin150086,China; 3.CollegeofLifeScience,JiamusiUniversity,Jiamusi154003,China
Abstract:The object of this study was to evaluate the effect of lead and cadmium stresses on growth and development of perennial ryegrass and to better understand the potential of perennial ryegrass for phytore mediation of Pb and Cd contaminated soils, and provide technical information for soil remediation planning. In a pot experiment, Pb was spiked at 0,300, 500, 1000, 1500 mg/kg, and Cd was spiked at,0,0.3, 3, 10, 50, 100 mg/kg. Further, Pb and Cd treatments were combined as follows: Pb0Cd0(CK), Pb300Cd0.3, Pb500Cd0.3, Pb1000Cd0.3, Pb1500Cd0.3, Pb300Cd10, Pb500Cd10, Pb1000Cd10, Pb1500Cd10. Measurements made included germination vigor, an establishment rate index, chlorophyll content, photosynthetic rate, transpiration, water use efficiencies of seedlings, as well as photosynthetically active radiation, and light use efficiency. The Pb and Cd contents of root, stem and leaf were determined in the seedlings. Compared to CK, there was no significant difference in germination at lower concentrations (300, 500 mg/kg) of Pb stress, but there was an adverse effect at higher concentrations (1000, and 1500 mg/kg). Lower concentrations of Cd stress promoted seed germination, but where Cd concentration was more than 10 mg/kg, there was inhibition of seed germination. With increase in Pb concentration, chlorophyll content also increased, but Cd stress the opposite tendency was noted. Photosynthetic rate decreased at lower concentrations and then increased at higher concentration under Pb or Cd single stress. Photosynthetic rate peaked when Pb concentration was 1000 mg/kg or Cd concentration was 10 mg/kg. The enrichment coefficient decreased with increase in concentration of the metals Pb or Cd. The Pb and Cd combinations showed stronger inhibitory effects on seed germination than single metal exposure stress. The enrichment ability of perennial ryegrass to Pb and/or Cd was root>stem>leaf. The enrichment coefficient of Cd was greater than that of Pb. Perennial ryegrass showed a better remediative effect on lightly polluted soils.
Key words:plumbum; cadmium; stress; perennial ryegrass; accumulation ability
*通信作者Corresponding author.
作者简介:冯鹏(1980-),男,内蒙古化德人,副研究员,博士。E-mail:fenggrass@163.com
基金项目:中国博士后科学基金(2013M541423)和黑龙江省政府博士后项目(LBH-Z12239)资助。
*收稿日期:2015-02-05;改回日期:2015-04-24
DOI:10.11686/cyxb2015069
http://cyxb.lzu.edu.cn
冯鹏,孙力,申晓慧,姜成,李如来,李增杰,郑海燕,张华,郭伟,韩旭东,洪亚南. 多年生黑麦草对Pb、Cd胁迫的响应及富集能力研究. 草业学报, 2016, 25(1): 153-162.
FENG Peng, SUN Li, SHEN Xiao-Hui, JIANG Cheng, LI Ru-Lai, LI Zeng-Jie, ZHENG Hai-Yan, ZHANG Hua, GUO Wei, HAN Xu-Dong, HONG Ya-Nan. Response and enrichment ability of perennial ryegrass under lead and cadmium stresses. Acta Prataculturae Sinica, 2016, 25(1): 153-162.
