青藏高原东北边缘高寒草甸植物种群生态位特征对牦牛放牧的响应
2016-02-23苗福泓薛冉郭正刚沈禹颖
苗福泓,薛冉,郭正刚,沈禹颖*
(1.兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室, 甘肃 兰州 730020;
2.青岛农业大学经济草本植物应用研究所,山东 青岛 266109)
青藏高原东北边缘高寒草甸植物种群生态位特征对牦牛放牧的响应
苗福泓1,2,薛冉1,郭正刚1,沈禹颖1*
(1.兰州大学草地农业科技学院, 草地农业生态系统国家重点实验室, 甘肃 兰州 730020;
2.青岛农业大学经济草本植物应用研究所,山东 青岛 266109)
摘要:在甘肃天祝县境内的青藏高原高寒草甸牦牛牧区设置不同强度放牧小区,采用野外样方调查法研究了放牧对青藏高原高寒草甸植物组成、种重要值和种群生态位特征的影响。结果表明,青藏高原高寒草甸随放牧强度增大植物种数逐渐减少,其中紫花针茅、西藏香青、高山韭等种类在重度放牧条件下消失。在中度条件下,优势种线叶嵩草的重要值最高;另一优势种珠芽蓼的重要值则是先增加后降低再升高,以轻度放牧条件下最高。在3个放牧强度条件下,珠芽蓼和线叶嵩草的生态位宽度均位居所有种群前两位,并随着放牧强度增加二者的生态位宽度呈增大趋势,在重度放牧条件下二者生态位宽度相等。3个放牧强度下,优势种珠芽蓼和线叶嵩草与各种群间的生态位重叠系数不尽相同,封育或者轻度放牧条件下,珠芽蓼是种群间竞争较强的一方,但当放牧强度达到中度及以上时,珠芽蓼则退出种群间竞争强度激烈的局面;线叶嵩草是重度放牧下,种群间竞争强度最大的一方。说明放牧强度对青藏高寒草甸植物群落植物物种组成、重要值、生态位宽度和生态位重叠值均产生一定的影响,中度放牧强度是珠芽蓼和线叶嵩草的适宜放牧强度。运用“生态位理论”分析青藏高原高寒草甸植物群落生态特征,其结果与“中度干扰理论”相吻合,中度放牧是青藏高原高寒草甸优势种适宜的放牧强度。
关键词:放牧强度;重要值;生态位宽度;生态位重叠值;高寒草甸群落
放牧是青藏高原高寒草甸生态系统中最常见干扰方式,对维持生态系统稳定和功能起着极为重要的作用[1]。放牧采食不仅直接影响高寒草甸植被的产生力,而且牲畜的排泄和践踏也会影响土壤的理化性质,其影响作用表现出积极和消极双重性[2],适宜的放牧强度不仅可以合理地利用草地资源[3],提高生态系统的多样性及生物量,优化群落中的物种组成[4],也可改善土壤的通透性,提高土壤含水量及养分含量[5]。当放牧强度超过草地的承载力时,不仅导致植被覆盖度降低,土壤径流增加,致使土壤养分流失,且改变植物群落种类组成和物种在群落中的地位,降低物种多样性,破坏生态系统的稳定性,进而导致草地发生退化[6-7]。“生态位理论”是解析植物群落物种组分变化的主要机制之一[8],通过定量刻画不同植物种群的生态位特征,可明确不同物种在群落内的地位变化[9-12],从而揭示放牧改变草甸生态系统的植物组分变化机制。研究高寒草甸植物生态位对放牧强度的响应,不仅可以解释放牧与生态系统的相互作用机制,也可为高寒草甸适宜的放牧强度提供理论依据。
目前关于青藏高原高寒草甸对放牧响应的研究主要集中于放牧对群落特征以及多样性的变化。这些研究主要反应草地群落的特征、多样性[13]、营养品质[14-15]以及物种组成[16]等对放牧效应的结果,而忽略了高寒草甸群落植物物种更替及生态位变化对放牧的响应过程。因此,本研究通过对不同放牧强度下高寒草甸植物群落物种特征的调查,阐明植物种群生态位变化规律,以揭示不同放牧强度下高寒草甸植物种群生态位特征的响应,对高寒草甸的合理放牧及保护具有重要的意义。
1材料与方法
1.1研究区自然概况
研究区域位于甘肃省天祝藏族自治县抓喜秀龙乡南泥沟八支沟(102°47′13″ E, 37°10′37″ N),年均温0.7℃,年降水量416 mm,≥0℃年积温1327.7~1738.2℃,海拔2900~3000 m,属高原气候类型,土壤主要为山地栗钙土和山地黑钙土。高寒草甸是其主要草地类型[17],珠芽蓼(Polygonumviviparum)和线叶嵩草(Kobresiacapillifolia)是主要优势物种,伴生种有紫花针茅(Stipapurpurea),矮火绒草(Leontopodiumnanum)和矮小风毛菊(Saussureapumila),主要毒草为狼毒(Stellerachamaejasme)和线叶龙胆(Gentianafarreri)。
1.2放牧样地设置与取样
放牧试验从2009年开始,放牧时间为每年的6月至9月,选取平均体重为76 kg的16月龄36头雌性牦牛。设4个放牧梯度,分别为零放牧、轻牧(0.75 牦牛/hm2)、中牧(1.00 牦牛/hm2)、重牧(1.25 牦牛/hm2),每个小区4 hm2,3次重复,在每个放牧小区内设置1 m×1 m的笼扣作为零放牧,9个放牧小区。
2012年8月中旬采样一次。在每个放牧小区内随机设置1 m×1 m的样方9个,保证样方离小区边缘的距离大于20 m,记录每个样方内植物种类、高度和盖度。齐地面分别剪下每个样方内的每个物种的地上部分,带回室内80℃烘干至恒重。
1.3指标计算
物种重要值(SDR)=(相对盖度+相对干重+相对高度+相对密度)/4。
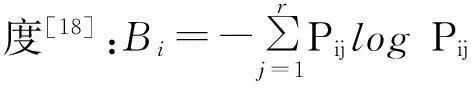
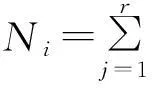

式中,Oik为物种i与物种k的生态位重叠值,nij和nkj为物种i和物种k在资源j上的优势度,即物种i与物种k在养分j的重要值,r为资源等级数,其值域为[0,1]。
1.4数据分析
采用Excel 2013对数据进行预处理,运用SPSS 16.0进行ANOVA方差分析。
2结果与分析
2.1不同放牧强度对高寒草甸植物群落组成及其重要值的影响
高寒草甸群落物种对放牧强度具有明显的响应,样方内植物物种数介于17~23之间,与轻度和中度放牧相比重度放牧种数减少的多3种,但轻度和中度相比减少的种数一样。与零放牧相比3个放牧强度均明显减少,而缺失的种不尽相同,封育条件下物种数最多(表1)。放牧强度大小不会改变高寒草甸植物群落优势种珠芽蓼和线叶嵩草在群落中的优势地位,但一些伴生种和稀有种发生了一定程度改变,总体表现为在重度放牧条件下禾本科的紫花针茅消失,毒杂草增多。放牧强度由封育增至0.75牦牛/hm2时,主要伴生种紫花针茅、瑞香狼毒和蓝花棘豆均无明显变化,线叶龙胆消失;放牧强度增至1.00 牦牛/hm2时,主要伴生种出现了冷蒿,但放牧强度达到1.25 牦牛/hm2时,主要伴生种紫花针茅消失。
珠芽蓼的重要值随放牧强度呈先增加后下降再升高,即在轻度放牧时最大,在中度放牧时降低,在重度放牧时又上升;而线叶嵩草的重要值以中度放牧强度最高,重度放牧下次之,与前两种放牧强度相比轻度放牧较低,但比零放牧时有所增加。主要伴生种矮火绒草重要值变化过程趋同于线叶嵩草,但伴生种紫花针茅重要值在放牧强度为1.00 牦牛/hm2下大于其他2个放牧梯度,且在1.25 牦牛/hm2时消失,而狼毒重要值在1.25 牦牛/hm2时最大。蓝花棘豆的重要值随放牧强度增加而先降低后升高,在放牧强度为1.25牦牛/hm2时最高,在1.00 牦牛/hm2时最低。垂穗披碱草的重要值随放牧强度增加而逐渐增加,在1.25 牦牛/hm2时最大。这说明放牧强度变化严重影响高寒草甸群落内不同种群的优势度,且变化过程趋异。
2.2放牧强度对高寒草甸群落内植物种群生态位宽度的影响
放牧强度对植物种群生态位宽度有明显影响,致使高寒草甸群落内各种植物种群利用资源顺序重新配置。不同放牧强度下,优势种珠芽蓼和线叶嵩草生态位宽度在所有的植物种群中占据前两位,说明其利用资源的能力最强(表2)。主要伴生种紫花针茅、蓝花棘豆和矮小风毛菊的生态位宽度随着放牧强度的增加而降低;阴山扁蓿豆生态位宽度随放牧强度增加表现为降低→升高→降低,说明该种在放牧强度下利用资源能力较高。在重度放牧时,垂穗披碱草和矮火绒草生态位宽度分别为0.764和0.326,高于其他放牧强度下的生态位宽度,说明在重度放牧下这2个种有较强的资源利用能力,但矮火绒草比零放牧条件下的生态位宽度明显降低。放牧强度为1.00 牦牛/hm2时,高寒草甸群落内雪地黄芪和矮金莲花均消失,而它们均在其他放牧强度下出现,以重度放牧下的生态位宽度值最大,说明其种群生存能力加强。其中雪地黄耆在零放牧和轻度放牧时,生态位宽度值均小于重度放牧。
2.3放牧强度对高寒草甸群落内植物种群生态位重叠值的影响
高寒草甸群落内种群间生态位重叠程度总体较高,4个放牧强度下生态位重叠系数均大于0.800(表3),说明种群间存在很大可能的竞争性。随放牧强度的增加,种群间生态位重叠系数表现为先降低后增加,在0.75牦牛/hm2时最大,而在1.00牦牛/hm2时最小,到1.25牦牛/hm2时又有所增加。
封育条件下,种群间生态位重叠值系数介于0.058~0.998之间,其中珠芽蓼和高山唐松草,矮金莲花和皱叶委陵菜的生态位重叠度最高(表4),其系数为0.998;兰石草和矮金莲花,矮小风毛菊和紫花针茅,团花马苋蒿和蓝花棘豆次之,分别为0.994,0.989,0.987;西藏香青和单花拉拉藤重叠度最低,系数为0.299。与优势种珠芽蓼生态位重叠度最高的是高山唐松草,系数为0.998,与优势种线叶嵩草生态位重叠度最高的是线叶龙胆,系数为0.962。说明此时优势种的主要竞争种群为高山唐松草和线叶龙胆,群落利用不当,正在向毒杂草演替。
放牧强度增加为0.75牦牛/hm2时,生态位重叠度最高的是珠芽蓼和蓝花棘豆,紫花针茅和皱叶委陵菜其系数为0.994(表5),而线叶嵩草和紫花针茅,西藏香青和皱叶委陵菜,高山嵩草和西藏香青次之,系数分别为0.988,0.979,0.971,阴山扁蓿豆和单花拉拉藤之间的生态位重叠度最低,系数为0.348。
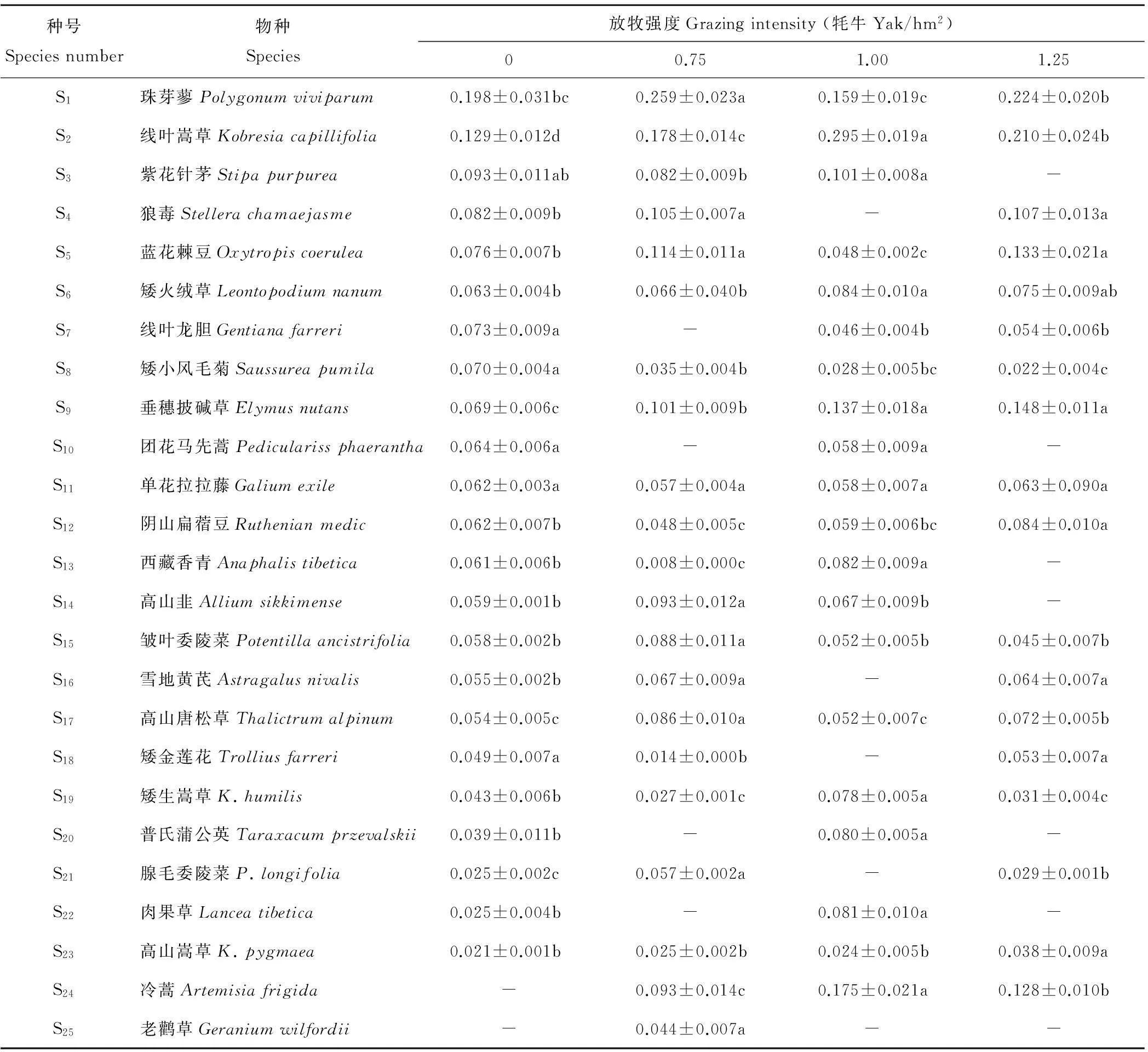
表1 不同放牧强度下高寒草甸植物群落的物种组成及其重要值
“-”表示此物种在该样方中未出现,同行不同小写字母表示在0.05水平上差异显著,下同。“-”represents that species does not appear in the plot, different lowercase letters in the same line represent significant difference atP<0.05, the same below.
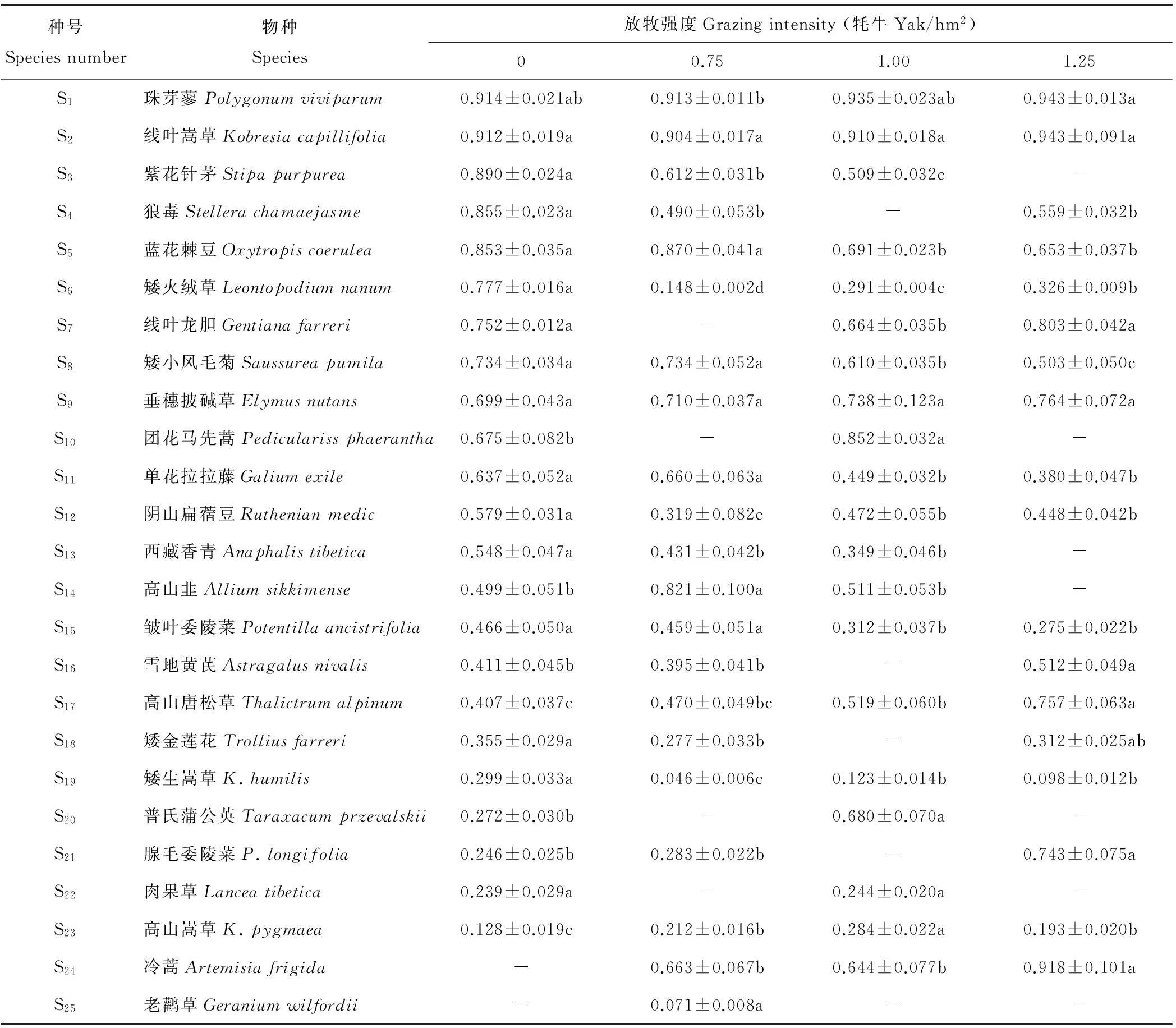
表2 不同放牧强度下植物种群的生态位宽度

表3 不同放牧强度下高寒草甸群落内种群间生态位重叠系数的平均值
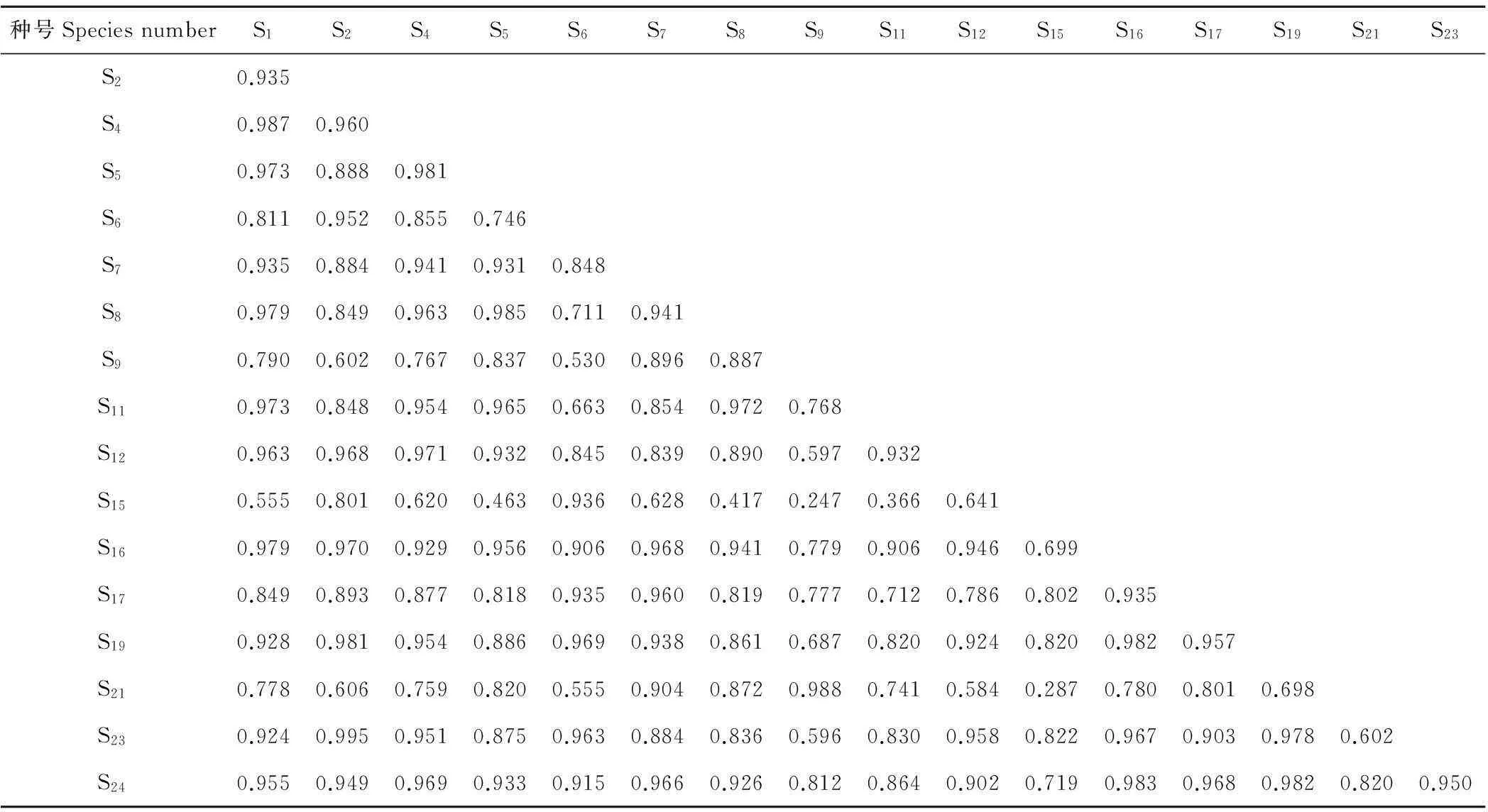
放牧强度增加为1.25牦牛/hm2时,生态位重叠度最高的是线叶嵩草和高山嵩草,其系数为0.995(表7),而腺毛委陵菜和垂穗披碱草、珠芽蓼和瑞香狼毒、矮小风毛菊和蓝花棘豆次之,系数分别为0.988,0.987,0.985,皱叶委陵菜和垂穗披碱草之间的生态位重叠度最低,系数为0.247。


表7 放牧强度为1.25 牦牛/hm2时不同种群间生态位的重叠值系数
种号SpeciesnumberS1S2S4S5S6S7S8S9S11S12S15S16S17S19S21S23S20.935S40.9870.960S50.9730.8880.981S60.8110.9520.8550.746S70.9350.8840.9410.9310.848S80.9790.8490.9630.9850.7110.941S90.7900.6020.7670.8370.5300.8960.887S110.9730.8480.9540.9650.6630.8540.9720.768S120.9630.9680.9710.9320.8450.8390.8900.5970.932S150.5550.8010.6200.4630.9360.6280.4170.2470.3660.641S160.9790.9700.9290.9560.9060.9680.9410.7790.9060.9460.699S170.8490.8930.8770.8180.9350.9600.8190.7770.7120.7860.8020.935S190.9280.9810.9540.8860.9690.9380.8610.6870.8200.9240.8200.9820.957S210.7780.6060.7590.8200.5550.9040.8720.9880.7410.5840.2870.7800.8010.698S230.9240.9950.9510.8750.9630.8840.8360.5960.8300.9580.8220.9670.9030.9780.602S240.9550.9490.9690.9330.9150.9660.9260.8120.8640.9020.7190.9830.9680.9820.8200.950
3讨论与结论
群落中的每个物种,具有独特的生态生理特性,使其与环境中各生态因子间形成特定的联系,并随生态因子梯度变化形成特有的分布格局。针对高寒草甸放牧因子而言,其放牧过程就是物种适应放牧引起的生境变化过程,从而导致物种的生态位发生分化,并表现为放牧演替系列上群落类型的更替现象。
本研究结果表明,青藏高原高寒草甸受不同放牧强度干扰引起植物种群数量、优势度及生态位发生不同程度的变化。重度放牧牦牛致使草甸植物的光合作用面积减少,进而影响植物营养物质的积累,种群数量下降。物种的生态位宽度受不同放牧强度影响而发生变化。在不同放牧强度下优势种珠芽蓼与线叶嵩草的生态位宽度均比伴生种宽,这与Moral[20]关于亚高山草甸植被自适应研究的结果基本一致。由于优势种在创建植物群落内部独特的生境条件及决定群落种类组成方面起主要作用,珠芽蓼和线叶嵩草的生活力及生态适应能力较强,繁殖速度较快,因而在群落内部适应小生境的能力及对资源利用能力都表现出较强的优势。在重度放牧情况下,由于牦牛采食量大于生长量,线叶嵩草和珠芽蓼的生长受到抑制,它们的竞争能力有所降低,使一些毒杂草(瑞香狼毒和线叶龙胆)的数量及优势度增加,表现为可食杂草和毒杂草的盖度增加,在生境梯度上表现出较宽的生态位,这与韩苑鸿等[21]在内蒙古典型草原上的研究结果基本一致。优势种线叶嵩草虽具有耐牧、耐旱等生态生物学特性,并在构建群落环境中起重要作用,生态位宽度也最大,但它的优势度随放牧强度增加而减小,这与松嫩平原羊草(Leymuschinensis)草地上,随放牧强度增加,植物优势种种群数量很小或消失的结论[22]基本一致。随放牧强度的增加,线叶嵩草等种群数量下降,而对高强度放牧能够做出较高适应性反应的杂类草(高山唐松草等)的优势度有增加的趋势,并在重度放牧成为次优势种或主要伴生种。这些种生态位宽度较大,表明它们具有较强的利用和适应环境的能力。豆科牧草阴山扁蓿豆随着放牧强度的增加,其优势度逐渐增加,可能是其在一定程度上有助于提高土壤氮和碳的固持力。作为草地退化先锋植物的垂穗披碱草、腺毛委陵菜等具有极强的适应放牧强度变化的能力[22],它们的生态位宽度随着放牧强度的增加而增加。
在群落内部同属物种之间由于生物-生态学特征更为相似,在一定程度上它们对环境资源的需求分化,导致生态位重叠程度降低,以便使它们共同生存于同一小生境中,具有长期生存适应的生态学意义。因此可以认为,群落内部种对生态位重叠程度的降低,是种间在不同放牧强度梯度下对资源利用分化的结果。在4个放牧强度梯度中,矮小风毛菊等一些伴生种表现出较大的生态位宽度,与优质牧草(紫花针茅)间有较大的生态位重叠,表明这些阔叶草本对资源利用和放牧强度梯度变化有较强的敏感性,这可能是由于长期放牧或者植物对家畜采食的响应,也说明放牧演替是一个较长时间尺度的过程[23]。
综上所述,随着放牧强度的增加,青藏高原高寒草甸群落物种组成、物种重要值、种群生态位宽度和生态位重叠值均发生一定程度的变化,样方内植物种数逐渐减少,且出现了较多的毒杂草的优势度增加。刘艳书等[24]同样在高寒草甸的研究中,发现杂类草的比例随着退化的加剧而增加,这与我们的研究结果吻合。优势种线叶嵩草的重要值先增加后降低,在中度下最高,优势种珠芽蓼的重要值先增加后降低最后增加,在轻度下最高。说明其在群落内的地位和作用发生了显著变化。在不同放牧强度下,优势种珠芽蓼和线叶嵩草的生态位宽度位居前两位,且随着放牧强度增加而增加。不同放牧强度下,与优势种珠芽蓼和线叶嵩草生态位重叠系数最大的物种不一样,当处于封育或者轻度时,珠芽蓼是种群间竞争强度最大的一方,但当放牧强度达到中度及以上时珠芽蓼则退出了种群间竞争强度最激烈的局面;而另一优势种线叶嵩草处于重度放牧时,成为种群间竞争强度最大的一方,因此,可以认为中度放牧(1.00牦牛/hm2)是当地高寒草甸两种优势种适宜的放牧强度。
我们的结论与“中度干扰理论”相吻合,运用“生态位理论”分析高寒草甸植物群落生态特征,再度支持了中度放牧是利于高寒草甸优势种的适宜放牧强度。
References:
[1]Ren J Z, Lin H L. Assumed plan on grassland ecological reconstruction in the source region of Yangtse River, Yellow River and Lantsang River. Acta Prataculturae Sinica, 2005, 14(2): 1-8.
凉州区作为全国水利改革试点县(区),2014年组建成立凉州区水利工程质量监督与安全管理站,负责全区水利工程建设质量监督与安全管理工作,委派监督员,对辖区内在建水利工程进行监督管理。近年来,按照质量监督分级管理权限,先后承担了全区高效节水灌溉、农村饮水安全、水环境综合治理、水库移民后期扶持、农业水价综合改革、抗旱应急、山洪灾害和山洪沟治理等工程的质量监督任务,质量监督管理工作取得了一定成绩。
[2]Jiang X L, Zhang W G, Yang Z Y,etal. The influence of disturbance on community structure and plant diversity of alpine meadow. Acta Botanica Boreali-Occidentalia Sinica, 2003, 23(9): 1479-1485.
[3]Hou F J, Ren J Z. Evaluation on trampling of grazed Gansu wapiti (CervuselaphuskansuensisPocock) and its effects on soil property in winter grazing land. Acta Ecologica Sinica, 2003, 23(3): 486-495.
[4]Miao F H, Guo Y J, Miao P F,etal. Influence of enclosure to community characteristics of alpine meadow in the northeastern edge region of Qinghai-Tibet Plateau. Acta Prataculturae Sinica, 2012, 21(3): 11-16.
[5]Li S Q, Wang X Z, Guo Z G,etal. Effects of short-term grazing on C and N content in soil and soil microbe in alpine meadow in the north-eastern edge of the Qinghai-Tibet Plateau. Chinese Journal of Grassland, 2013, 35(1): 55-60, 66.
[6]Guo Z G, Long R J, Niu F J,etal. Effect of highway construction on plant diversity of grassland communities in the permafrost regions of the Qinghai-Tibet plateau. The Rangeland Journal, 2007, 29(2): 161-167.
[7]Wu N, Luo P. Rethinking on the ecological theories related to the ecological construction and management of the high-frigid grasslands in the upper Yangtse River. Chinese Journal of Applied and Environmental Biology, 2004, 10(4): 537-542.
[8]Grinnell J. The niche-relationships of California Thrasher. The Auk, 1917, 34(4): 427-433.
[9]Nee S. The neutral theory of biodiversity: do the numbers and up. Functional Ecology, 2005, 19(1): 173-176.
[10]Jia T T, Mao L, Guo Z G. Effect of available burrow densities of plateau pika (Ochotonacurzoniae) on plant niche of alpine meadow communities in the Qinghai-Tibet Plateau. Acta Ecologica Sinica, 2014, 34(4): 869-877.
[11]Yuan C J, Yu L F, An M T,etal. Study on the niche of main tree species ofOrmosiasaxatiliscommunity in natural forest. Ecological Science, 2013, 32(4): 487-493.
[12]Wang X, Hu Y K, Rehemudula A,etal. Effects of soil salt on the niche of main plant species in alpine meadow. Chinese Journal of Applied Ecology, 2008, 19(7): 1448-1454.
[13]Yang L M, Han M, Li J D. Plant diversity change in grassland communities along a grazing disturbance gradient in the northeast China transect. Acta Phytoecologica Sinica, 2001, 25(1): 110-114.
[14]Milchunas D G, Varnamkhasti A S, Lauenroth W K,etal. Forage quality in relation to long-term grazing history, current-year defoliation, and water resource. Oecologia, 1995, 101(3): 366-374.
[15]Duan M J, Gao Q Z, Wan Y F,etal. Effect of grazing on community characteristics and species diversity ofStipapurpureaalpine grassland in Northern Tibet. Acta Ecologica Sinica, 2010, 30(14): 3892-3900.
[16]Dong Q M, Zhao X Q, Ma Y S,etal. Effects of grazing on the major plant populations niche in alpineKobreciaparvameadow cold-seasonal pastureland. Chinese Journal of Grassland, 2006, 28(6): 10-17.
[17]Song L M, Ma X H, Wei Y L,etal. Causes and countermeasures of degradation and desertification of natural meadow in Haibei District of Qinghai Province. Pratacultural Science, 2009, 26(7): 186-190.
[18]Levins R. Evolution in Changing Environments: Some Theoretical Explorations[M]. Princeton: New Jersey Princeton University Press, 1968.
[19]Pianka E R. The structure of lizard communities. Annual Review of Ecology and Systematics, 1973, 4: 53-74.
[20]Moral R D. Vegetation ordination of subalpine meadows using adaptive strategies. Canadian Journal of Botany, 1983, 61(12): 3117-3127.
[21]Han Y H, Wang S P, Chen Z Z. Responses of the major plant populations of Inner Mongolia typical steppe to grazing rate based on niche considerations. Acta Agrestia Sinica, 1999, 7(3): 204-210.
[22]Cole D N. Experimental trampling of vegetation II. Predictors of resistance and resilience. Journal of Applied Ecology, 1995, 32(1): 215-224.
[23]Wang R Z. The niche breadths and niche overlaps of main plant populations inLeymuschinensisgrassland for grazing. Acta Phytoecologica Sinica, 1997, 21(4): 304-311.
[24]Liu Y S, Fan J W, Li Y Z,etal. Plant community productivity and diversity on alpine meadow steppe in Tree River Headwater Region, Qinghai Province under different denudation levels. Acta Prataculturae Sinica, 2014, 23(3): 1-7.
参考文献:
[1]任继周, 林慧龙. 江河源区草地生态建设构想. 草业学报, 2005, 14(2): 1-8.
[2]江小蕾, 张卫国, 杨振宇, 等. 不同干扰类型对高寒草甸群落结构和植物多样性的影响. 西北植物学报, 2003, 23(9): 1479-1485.
[3]侯扶江, 任继周. 甘肃马鹿冬季放牧践踏作用及其对土壤理化性质影响的评价. 生态学报, 2003, 23(3): 486-495.
[4]苗福泓, 郭雅婧, 缪鹏飞, 等. 青藏高原东北边缘地区高寒草甸群落特征对封育的响应. 草业学报, 2012, 21(3): 11-16.
[5]李世卿, 王先之, 郭正刚, 等. 短期放牧对青藏高原东北边缘高寒草甸土壤及微生物碳氮含量的影响. 中国草地学报, 2013, 35(1): 55-60, 66.
[7]吴宁, 罗鹏. 长江上游高寒草地生态建设和管理中生态理论的若干质疑. 应用与环境生物学报, 2004, 10(4): 537-542.
[10]贾婷婷, 毛亮, 郭正刚. 高原鼠兔有效洞穴密度对青藏高原高寒草甸群落植物生态位的影响. 生态学报, 2014, 34(4): 869-877.
[11]袁丛军,喻理飞,安明态,等. 岩生红豆天然林主要树种生态位研究. 生态科学, 2013, 32(4): 487-493.
[12]王鑫, 胡玉昆, 热合木都拉·阿迪拉, 等. 土壤盐分对高寒草甸主要植物生态位的影响. 应用生态学报, 2008, 19(7): 1448-1454.
[13]杨利民, 韩梅, 李建东. 中国东北样带草地群落放牧干扰植物多样性的变化. 植物生态学报, 2001, 25(1):110-114.
[15]段敏杰, 高清竹, 万运帆, 等. 放牧对藏北紫花针茅高寒草原植物群落特征的影响. 生态学报, 2010, 30(14): 3892-3900.
[16]董全民, 赵新全, 马玉寿, 等. 放牧对高寒小嵩草草甸冷季草场主要植物种群生态位的影响. 中国草地学报, 2006, 28(6): 10-17.
[17]宋理明, 马晓虹, 魏永林, 等. 海北州天然草地退化沙化成因与对策. 草业科学, 2009, 26(7): 186-190.
[21]韩苑鸿, 汪诗平, 陈佐忠. 以放牧率梯度研究内蒙古典型草原主要植物种群的生态位. 草地学报, 1999, 7(3): 204-210.
[23]王仁忠. 放牧影响下羊草草地主要植物种群生态位宽度与生态位重叠的研究. 植物生态学报, 1997, 21(4): 304-311.
[24]刘艳书, 樊江文, 李愈哲, 等. 三江源地区不同剥蚀退化高寒草甸群落生物量与多样性特征.草业学报, 2014, 23(3): 1-7.
*Influence of yak grazing on plant niche characteristics in alpine meadow communities at the northeastern edge of the Qinghai-Tibetan Plateau
MIAO Fu-Hong1,2, XUE Ran1, GUO Zheng-Gang1, SHEN Yu-Ying1*
1.StateKeyLaboratoryofGrasslandAgro-ecosystems,CollegeofPastoralAgricultureScienceandTechnology,LanzhouUniversity,Lanzhou730020,China;2.InstituteofEconomicHerbPlants,QingdaoAgriculturalUniversity,Qingdao266109,China
Abstract:The grazing intensity changes the plant composition of alpine meadow. A field survey was carried out to determine the effect of grazing intensities on plant niche characteristics of alpine meadow communities in the Qinghai-Tibet Plateau. This study showed that the number of plant species appearing in plots reduced as the grazing intensity increased, andAnaphalistibetica,StipapurpureaandAlliumsikkimensedisappeared when grazing intensity exceeded 1.25 yak/ha. The numbers of unpalatable plants increased when the grazing intensity in plots was over 1.00 yak/ha. With increasing grazing intensity, the importance value ofKobresiacapillifolia, a dominant species, initially increased and then decreased, peaking at 1.00 yak/ha grazing intensity, and the importance value ofPolygonumviviparum, another dominant species peaked at 0.75 yak/ha grazing intensity. The niche breadth ofK.capillifoliaandP.viviparumwas greater than that of other species in alpine meadow communities differing in grazing intensity and increased as the grazing intensity increased. The plants with the biggest niche overlap withK.capillifoliaandP.viviparumwere different.P.viviparumwas one of two populations which had a bigger coefficient of niche overlap at 0 and 0.75 yak/ha grazing intensity, however,P.viviparumdid not appear in the populations which had the biggest coefficient of niche overlap when the grazing intensity was over 0.75 yak/ha.K.capillifoliawas one of two populations which had the biggest coefficient of niche overlap at 1.25 yak/ha grazing intensity. These results suggest that the grazing intensity changed the plant composition, species importance values, niche breadth and niche overlap of alpine meadow communities, and that a grazing intensity of 1.00 yak/ha was advantageous to the dominant species,P.viviparumandK.capillifolia, in alpine meadow at the northeastern edge of the Qinghai-Tibetan Plateau.
Key words:grazing intensity; important value; niche breadth; niche overlap; alpine meadow community
*通信作者Corresponding author. E-mail:yy.shen@lzu.edu.cn
作者简介:苗福泓(1988-),男,山东青岛人,博士。E-mail:miaofh10@lzu.edu.cn
基金项目:青藏高原社区草畜高效转化关键技术专项(201203006),长江学者和创新团队发展计划资助-草地农业系统耦合与管理(IRT13019)和农业部公益性行业专项(200903060)资助。
*收稿日期:2015-01-20;改回日期:2015-06-29
DOI:10.11686/cyxb2015033
http://cyxb.lzu.edu.cn
苗福泓, 薛冉, 郭正刚, 沈禹颖. 青藏高原东北边缘高寒草甸植物种群生态位特征对牦牛放牧的响应. 草业学报, 2016, 25(1): 88-97.
MIAO Fu-Hong, XUE Ran, GUO Zheng-Gang, SHEN Yu-Ying. Influence of yak grazing on plant niche characteristics in alpine meadow communities at the northeastern edge of the Qinghai-Tibetan Plateau. Acta Prataculturae Sinica, 2016, 25(1): 88-97.
