大别山五针松的研究进展及保护对策
2016-02-11吴甘霖项小燕段仁燕王志高
吴甘霖,项小燕,段仁燕,王志高
(安庆师范大学生命科学学院,安徽安庆246133)
大别山五针松的研究进展及保护对策
吴甘霖,项小燕,段仁燕,王志高
(安庆师范大学生命科学学院,安徽安庆246133)
本文介绍了大别山五针松命名的由来、现有分布范围及研究现状,并提出了相应的保护策略。
大别山五针松;分布范围;研究现状;保护策略
1 大别山五针松在我国的分布情况
大别山五针松(Pinus dabeshanensis W.C. Cheng&Y.W.Law),隶属于松科(Pinaceae)松属(Pinus),模式产地位于安徽省岳西县鹞落坪国家级自然保护区门坎岭[1]。1956年该种首次被发现,经林木分类学家郑万钧教授鉴定,称之为安徽五针松(或安徽松),并于1961年在《中国树木学》第一分册上对其特征进行了描述[2]。1975年,郑教授再度考订该种,将其与五针松组其他物种反复综合比较后,与刘玉壶先生将其正式命名为Pinus dabeshanensis W.C.Cheng&Y.W.Law,即大别山五针松[3]。
据资料记载,大别山五针松零星分布于安徽省的岳西县和金寨县、湖北省的英山县和罗田县、河南省的商城县和新乡县交界的大别山区[4]。笔者近几年多次实地考察后发现,湖北的罗田和河南的商城、新乡现并未发现大别山五针松的踪迹,说明大别山五针松的分布区日益狭窄。主要种群散生于岳西县大王沟海拔900~1 300m的范围内,共约200余株,多分布在阴坡近山脊处,且多为中老龄级个体;其次是金寨县,成年个体约10~20株,林下散生少量幼苗;其他地方仅1~2株孤存于脆弱的生境中,如门坎岭仅有1株成年个体孤立于悬崖边;英山县也仅有2株成年植株,且1株虫害严重,具体分布范围如图1所示。
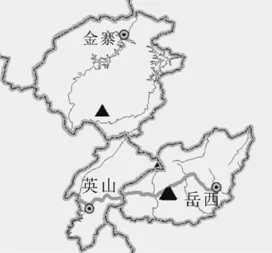
图1 大别山五针松的分布图(▲表示分布地,其大小表示种群大小)
由于人为破坏和自身特性,大别山五针松种群日益减少,被列为国家二级珍稀树种之一[5]。
2 大别山五针松的生物学特性
大别山五针松(图2)为常绿乔木,一年生枝条微褐色,针叶横断面具2个边生的树脂道。树干通直圆满,材质细致均匀,茎有早材和晚材之分,且早材的比例较大[6]。花芽常在3~4月形成,5月中下旬花粉成熟并散发,花粉具有气囊,此时,雌球花张开接受传粉,球果翌年9月中上旬成熟[7],下垂,种鳞木质,具有明显的反卷现象[8]。种鳞基部着生种子2枚,种子三角状卵圆形,褐色,具短木质化种翅,种皮较厚,种子大小和重量均高于五针松组其他种[9]。

图2 大别山五针松
近几年有部分研究者对大别山五针松开展了一些生物学方面的研究。项小燕等[10]对大别山五针松母树林的花粉飞散规律进行了研究,分析了花粉飞散的时空特点及与气象因子的关系。王雷宏等[11]对金寨县天马保护区大别山五针松种群的种间联结及相关性进行了研究,认为蜡瓣花(Corylopsis sinensis)、白檀(Symplocos paniculata)和荚蒾(Viburnum dilatatum)等与大别山五针松的生态位不同,应予以控制以促进大别山五针松种群的发展。项小燕等[12]对大别山五针松的种内和种间竞争开展了研究,结果表明,大别山五针松的竞争主要来自种间。种内与主要伴生种间的竞争关系从强到弱依次为短柄枹、大别山五针松种内、黄山松、满山红、茅栗、金缕梅、紫茎、四照花、灯台树、鹅耳枥。竞争强度随对象木胸径的增大而减小,当对象木的胸径小于25 cm时,所受到的竞争强度较大;当胸径在25 cm以上时,竞争强度变化很小,二者符合幂函数关系。项小燕等[13]对大别山五针松的种群结构及动态也开展了研究,结果发现大别山五针松种群数量少,结构存在波动性。幼苗阶段个体较丰富,幼树阶段个体较少,种群趋于衰退。
3 大别山五针松的遗传结构特点
自上世纪50年代大别山五针松被发现以来,研究者们相继开展了一些研究工作。早期的研究主要集中在材质特性、力学特征和核型等方面[6,14-15]。研究发现大别山五针松的材质及大多物理力学指标均高于红松(Pinus koraiensis),染色体数目为2n=24。这些研究表明大别山五针松具有重要的应用价值。
韩建伟等[9,16]对大别山五针松的花粉和种子萌发等方面进行了初步研究,发现大别山五针松花粉的离体萌发率仅为33.15%,人工破壳处理和添加激素等方式均可促进种子萌发。因而认为大别山五针松的濒危可能与较低的花粉萌发率、种子萌发受较厚种皮的限制及种子存在生理休眠等因素有关。
Liu等[17]通过线粒体单倍型(mtDNA)研究发现,大别山五针松与同组的海南五针松线粒体单倍型并不相同。Xiang等[18]通过转录组测序的方法从大别山五针松针叶序列中开发出SSR分子标记,并用所开发的SSR标记对大别山五针松的遗传多样性进行了研究。同松类其他物种相比,大别山五针松的遗传多样性属于中等水平,种群间遗传分化较小,这与种群间存在较大的基因流有关[19]。但由于小种群易导致近交衰退,而近交衰退会导致种群适合度降低和对环境的适应能力下降[20-21],因而需采取相应措施保护和扩大现有种群,以挽救这一濒危物种。
4 大别山五针松的保护策略与研究展望
(1)由于大别山五针松花芽形成及花粉飞散均受到林分密度的影响,且生长过程中受到竞争的影响,因而,在保护中应择伐次林层和灌木层的一些阔叶树种,如满山红、蜡瓣花等,加强通风,促使花芽形成和花粉飞散,提高自然种群种子萌发率,为幼苗生长提供林窗,提高幼苗成活率。
(2)择伐主林层中占优势的短柄枹、茅栗等,降低对大别山五针松的竞争。
(3)由于目前大别山五针松自然种群数量太少,短期内难以复壮,迁地保护是扩大大别山五针松种群的重要措施之一。建议迁地保护时应从遗传多样性高的种群适当的采集种子,并采收包含稀有等位基因个体的种子,进行人工育苗,保护基因库,防止遗传资源的丢失。
(4)栽培选址时选择光照和水分条件较好的地点造林,注意种植密度以提高种植园的结实量及种子的品质。尽管前人在以上方面对大别山五针松开展了相应的研究,但仍有众多问题尚未解决,如自然环境下,种子雨和种子库及种子萌发特点尚不清楚,这将直接影响大别山五针松自然种群的更新和种群大小的稳定;精细规模的基因流研究,尤其是花粉流的贡献大小尚待研究。此外,繁育系统能决定居群的遗传结构和种内进化模式,大别山五针松的繁育系统研究也有待进一步开展。
[1]吴轶峰.岳西大别山五针松的种群分布、生存现状与保护对策[J].安徽农学通报,2010,16(13):193-194.
[2]郑万钧.中国树木学(第一分册)[M].1版.南京:江苏人民出版社,1961:191.
[3]郑万钧,傅立国,诚静容.中国裸子植物[J].植物分类学报,1975,13(4):85.
[4]汪为民.濒危植物——大别山五针松[J].安徽林业,1997(4):5. [5]王爱华,王传贵,柯曙华,等.大别山五针松木材主要解剖特征及其变异研究[J].安徽农业大学学报,2004,31(2):173-177.
[6]江泽慧.大别山五针松在不同立地条件下木材构造及材性研究[J].安徽农学院学报,1985(1):51-57.
[7]祝霞灿.大别山五针松生长发育情况调查[J].安徽林业科技,2005,4:16-17.
[8]彭镇华.五针松组起源中国[J].安徽农业大学学报,1999,26(1):1-8.
[9]韩建伟,张智勇,王恩茂,等.大别山五针松种子特性及促进种子萌发的研究[J].中国农学通报,2014,30(1):5-10.
[10]项小燕,张小平,段仁燕,等.濒危植物大别山五针松母树林花粉传播规律[J].广西植物,2014,34(3):333-337.
[11]王雷宏,黄庆丰,蒲发光,等.天马自然保护区大别山五针松与群落中优势种的种间关系[J].长江流域资源与环境,2014,23(7):960-965.
[12]项小燕,吴甘霖,段仁燕,等.大别山五针松种内和种间竞争[J].生态学报,2015,35(2):389-395.
[13]项小燕,吴甘霖,段仁燕,等.大别山五针松种群结构及动态研究[J].长江流域资源与环境,2016,25(1):55-61.
[14]王传贵,柯曙华.大别山五针松木材结构与材性[J].南京林业大学学报(自然科学版),1997,21(3):51-54.
[15]邢自琢,康忠铭.大别山五针松核型研究[J].安徽农学院学报, 1989(1):48-52.
[16]韩建伟,王恩茂,张智勇,等.珍稀濒危物种大别山五针松花粉离体萌发的研究[J].中国农学通报,2014,30(13):15-19.
[17]LIU L,HAO Z Z,LIU Y Y,et al.Phylogeography of Pinus armandii and its relatives:heterogeneous contributions of geography and climate changes to the genetic differentiation and diversification of Chinesewhite pines[J/OL].PloSONE,2014,9(1):e85920[2016-02-01].http://dx.doi.org/10.1371/journal. pone.0085920.
[18]XIANG X Y,ZHANG Z X,WANG Z G,et al.Transcriptome sequencing and development of EST-SSR markers in Pinus dabeshanensis,an endangered conifer endemic to China[J]. MolecularBreeding,2015,35:158.
[19]XIANG X Y,ZHANG Z X,DUAN R Y,et al.Genetic diversity and structure of Pinus dabeshanensis revealed by expressed sequence tag-simple sequence repeat(EST-SSR)markers[J]. BiochemicalSystematic and Ecology,2015,61:70-77.
[20]DELMASC E L,CHEPTOU PO,ESCARAVAGE N,et al.High lifetime inbreeding depression counteracts the reproductive assurance benefitof selfing in a mass-flowering shrub[J].BMC Evolutionary Biology,2014,14:243.
[21]THEODOROULK,COUVETD.Theefficiency ofclose inbreeding to reducegenetic adaptation to captivity[J].Heredity,2015,114:38-47.
Research Progressand Conservation Measuresof Pinus dabeshanensis
WU Gan-lin,XIANG Xiao-yan,DUAN Ren-yan,WANG Zhi-gao
(School of Life Science,Anqing Normal University,Anqing,Anhui246133,China)
This paper summarizes the named origin,current distribution and research progress of Pinus dabeshanensis, and puts forward the conservationmeasures.
Pinus dabeshanensis;distribution;research progress;conservation measures
Q941
A
1007-4260(2016)04-0097-03
时间:2017-1-3 17:19
http://www.cnki.net/kcms/detail/34.1150.N.20170103.1719.025.html
2016-03-03
国家林业公益性行业科研专项(201304314)和安徽省教育厅自然科学重点项目(KJ2016A437)。
吴甘霖,男,安徽金寨人,硕士,安庆师范大学生命科学学院教授,研究方向为植物资源保护与利用。
E-mail:wugl@sina.cn
10.13757/j.cnki.cn34-1150/n.2016.04.025
