中国伞形科柴胡属果实结构的系统学价值
2016-01-28常修奇刘玫程薪宇王宇婷魏雪莹王雪微
常修奇,刘玫,程薪宇,王宇婷,魏雪莹,王雪微
(哈尔滨师范大学生命科学与技术学院,黑龙江省普通高等学校植物生物学重点实验室,黑龙江 哈尔滨 150025)
中国伞形科柴胡属果实结构的系统学价值
常修奇,刘玫*,程薪宇,王宇婷,魏雪莹,王雪微
(哈尔滨师范大学生命科学与技术学院,黑龙江省普通高等学校植物生物学重点实验室,黑龙江 哈尔滨 150025)
摘要:本文首次深入研究了中国柴胡属29种,5变种及2变形果实的三维结构。结构显示果实结构变化较大,分果表面平滑, 横切面近圆形,脊略微突出。中果皮有两种分泌结构,伴生油管及非伴生油管(油管)。前者仅存在于少数物种,且直径小,后者包括不分支及分支油管。不分支油管存在于多数物种的沟(1~5)和结合面(2~7),油管通常与分果近等长,但亦存在短油管。少数物种的油管呈分支状,在中果皮不规则分布。种皮由1或2层薄壁细胞构成。胚乳在结合面平直或微凹。中果皮无结晶。心皮柄多为2个,在结合面彼此相对。柴胡属果实的多数特征存在于芹亚科其他类群,但分支状油管及2层细胞的种皮仅见于牵环花亚科,变豆菜亚科及芹亚科的基础类群。柴胡属果实的解剖结构支持分子系统学将柴胡属位于芹亚科其他类群的基础位置。
关键词:柴胡属;果实解剖;油管;伞形科;系统学
The phylogenetic significance of fruit structures inBupleurumof the family Apiaceae
CHANG Xiu-Qi, LIU Mei*, CHENG Xin-Yu, WANG Yu-Ting, WEI Xue-Ying, WANG Xue-Wei
CollegeofLifeScienceandTechnology,HarbinNormalUniversity,KeyLaboratoryofPlantBiology,CollegeofHeilongjiangProvince,Harbin150025,China
Abstract:The three-dimensional structure of fruits of 29 species, 5 varieties and 2 forms in the genus Bupleurum, from China were studied in depth and are reported for the first time in this paper. The fruit structures were found to vary greatly. The mericarp surfaces are smooth, approximately circular in transverse section, and ribs are slightly prominent. Two types of secretory structures, rib ducts and vittae, occur in the mesocarps. The former which are small in diameter are found in a few taxa and the latter include unbranched and branched vittae. The vittae occur in the furrow (1-5) and commissure (2-7) in most taxa, and they are similar to mericarps in length, usually unbranched, and sometimes short. The vittae are branched in a few taxa and dispersed in the mesocarps. The testa is composed of one or two layers of parenchyma cells. The endosperm is flat or slightly concave on the commissural side. Crystals are not present in the mesocarp. There are usually two carpophores, which are arranged on opposite sides of the commissural plane. Most features of the genus are present in other groups of the Apioideae, but branching vittae and testa with two layers of cells are only found in the Azorelloideae, Saniculoideae, and some groups of the Apioideae. The anatomical structures of Bupleurum fruits support its continued classification in the Apioideae, as shown by molecular studies.
Key words:Bupleurum; fruit anatomy; vitta; Apiaceae; systematics
柴胡属(Bupleurum)是伞形科(Apiaceae)芹亚科(Apioideae)的大属之一,全世界约180种,主要分布于欧洲,亚洲,非洲及北美[1-3]。中国有42种,其中22种为特有种[4]。植物主要为多年生(稀为一年生)草本,少为木本,单叶[3-5]。芹亚科植物通常为复叶,具单叶的物种主要存在于天胡荽亚科(Drude’s Hydrocotyloideae)和变豆菜亚科(Saniculoideae)[1],前者已被分子系统学证明不是个自然类群, 其中大部分属被分别移入两个新的亚科,参棕亚科(Makinlayoideae)及牵环花亚科(Azorelloideae)[6]。花粉研究[7-9]表明柴胡属的花粉同参棕亚科,牵环花亚科及变豆菜亚科类似,近菱形,近圆形及椭圆形,为原始类型。染色体数亦同这几个亚科相似,通常为n=4, 6, 7 或 8, 很少为芹亚科多数物种具有的染色体数n=11[10-12]。
分子系统学研究[13-27]显示芹亚科不是个自然的类群,传统位于这一亚科的非洲特有属Polemanniopsis及Steganotaenia应靠近变豆菜亚科,其他一些属Anginon,Heteromorpha,Dracosciadium,Polemannia,Glia这5个属组成一个新的族Heteromorpheae[24],Annesorhiza及Chamarea应位于芹亚科的基础位置[24]。尽管分子系统研究证明柴胡属是单系类群,比这些非洲特有属进化,但仍位于芹亚科的基础位置[6,28-31]。
柴胡属的果实具有游离的心皮柄,5个丝状的脊,每个沟里通常具1~3个油管,通常无伴生油管[1,3]。Snogerup 和Snogerup[32]指出柴胡属果实解剖结构的变化大于伞形科任何一个类群。Liu[33]及王长宝等[34]揭示了柴胡属果实油管数量变化较大。然而迄今为止缺少这一属果实结构的详细报道。伞形科果实结构常被视为其亚科、族及属的重要分类特征[1,35-36],近年来随着果实研究的深入,进一步揭示了果实微形态(如油管、维管束、结合面及心皮柄等)对伞形科系统分类的重要性,为分子系统学的研究提供了重要的形态学支持[33-40]。本文的目的是深入研究中国柴胡属果实的三维结构,比较不同物种果实结构的异同,并将所研究的结构与已研究的伞形科的其他类群相比较,为分子系统学确立的柴胡属的系统位置提供形态学依据,并为进一步深入研究世界柴胡属及伞形科系统学奠定了形态学基础。
1材料与方法
1.1 材料
选取中国柴胡属29种、5变种及2变型作为研究对象,物种名称及凭证标本信息见表1。果实材料分别取自于中国科学院华南植物园标本馆(IBSC-South China Botanical Garden Herbarium),中国科学院沈阳应用生态研究所标本馆(IFP-Herbarium, Institute of Appling Ecology, CAS),中国科学院昆明植物研究所标本馆(KUN-Herbarium, Kunming Institute of Botany, CAS),中国科学院植物研究所标本馆(PE-Herbarium, Institute of Botany, CAS),四川大学标本馆(SZ-Herbarium, Sichuan University),中国科学院新疆生态与地理研究所标本馆(XJBI-Herbarium, Xinjiang Institute of Ecology and Geography,CAS)。
1.2 方法
观察果实外形,油管及心皮柄的结构:用具数码相机(Olympus DP 70)的解剖镜(Olympus SP-350)观察柴胡属果实外形及心皮柄的结构并照相。将干的果实放入热水(约90℃)中浸泡,待材料完全吸水膨胀并沉入水底(约2 h)时,取出果实,撕取果皮,将其放在载玻片上并加1~2滴50%甘油水溶液,用具数码相机(Olympus DP 26)的显微镜(Olympus BX 51)观察油管结构并照相。
GMA(glycolmethacrylate:乙二醇甲基丙烯酸酯)切片:每个物种取2~3个果实,根据Feder和O’Brien[41]的方法,果实经吸水膨胀后,放入FAA(formalin-acetic acid-alcohol: 福尔马林-冰醋酸-酒精)固定,时间不少于24 h。材料经50%乙醇(4~6 h),100%乙醇(4~6 h),异丙醇(6 h)和正丁醇(6 h)脱水, 每步各重复1次。接着材料进入GMA渗透,共3次,第1和2次各1 d,第3次不少于5 d。之后将材料及GMA装入无色胶囊,置于60℃温箱聚合(24 h)。用Leica Ultracut R 切片机切片,厚度约2~3 μm。同样根据Feder和O’Brien[41]的方法,将切片用甲苯胺蓝染色,中性树胶封片。用具数码相机(Olympus DP 26)的显微镜(Olympus BX 51)观察果实结构并照相。
2结果与分析
本文详细地描述了中国柴胡属果实的形态结构,其主要特征见表2。图1A~L展示了分果的果皮,伴生油管及非伴生油管,维管束,种皮及心皮柄等特征,图2A~JJ展示了不同物种果实结构的变化。
分果形状:研究的柴胡分果(mericarp)的横切面均为近圆形(图1I, J,2A~JJ),具5个脊,1个中脊(median rib),2个侧脊(lateral rib)及2个边脊(marginal rib)(图1I, 2A)。多数物种的脊略微突起,如线叶柴胡,金黄柴胡及锥叶柴胡(图2A~C),少数物种,即北柴胡,秦岭柴胡,有柄柴胡,红柴胡及小柴胡的脊具较明显突起,大于果皮厚度的2倍(图2E, U, BB, DD, GG)。

表1 用于研究果实结构的中国柴胡属物种的凭证标本信息及其采集地

表2 中国柴胡属果实的重要分类特征Table 2 Summary of taxonomically important characters of the fruits in various taxa of Bupleurum in China
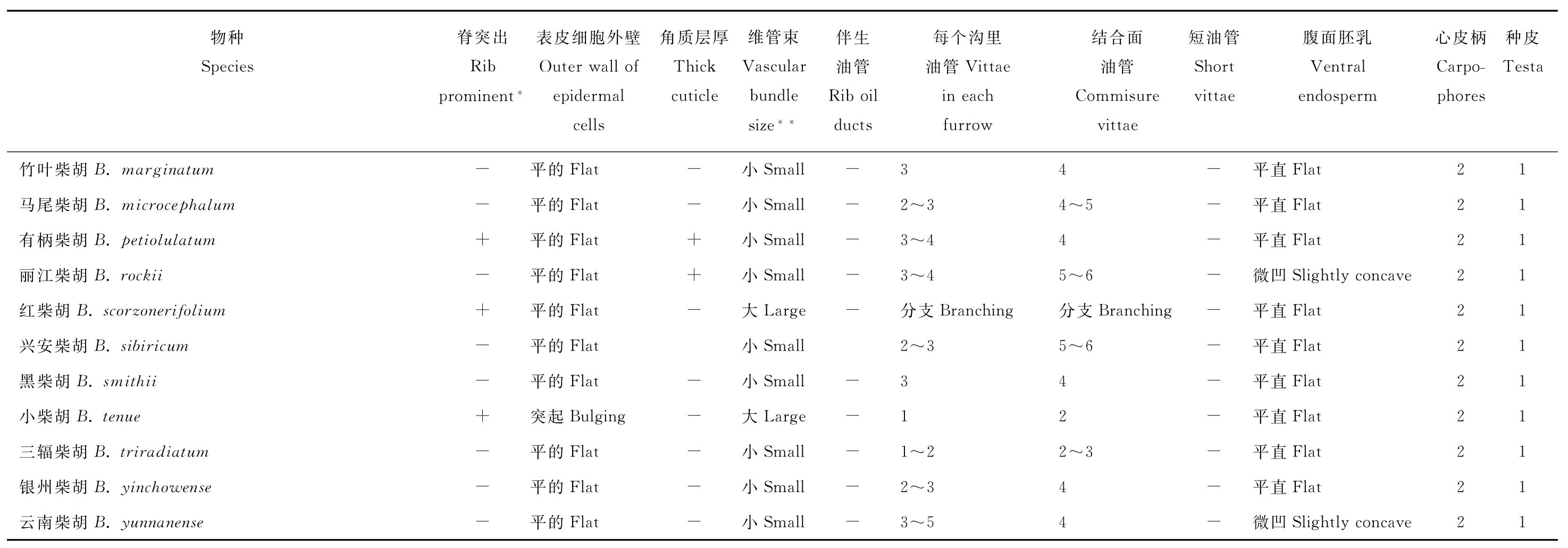
续表2 Continued
分果表面,表皮,中果皮及内果皮:分果表面平滑,具角质层(cuticle),多数物种的角质层较薄,横切面不明显(图1G),但少数物种,如金黄柴胡,紫花鸭趾柴胡及细柄柴胡的角质层较厚,约等于表皮细胞的厚度(图1F)。表皮(epidermis)为1层扁平状细胞,多数物种其细胞的外壁是平的(图1F, H),细柄柴胡、纤细柴胡及小柴胡的外壁向外突起,呈弧形(图1I)。中果皮(mesocarp)通常由5~8层近圆形薄壁细胞构成(如图1F, H)。内果皮(endocarp)为1层长方形薄壁细胞,其细胞的长轴与果实的表面平行(图1F, H)。
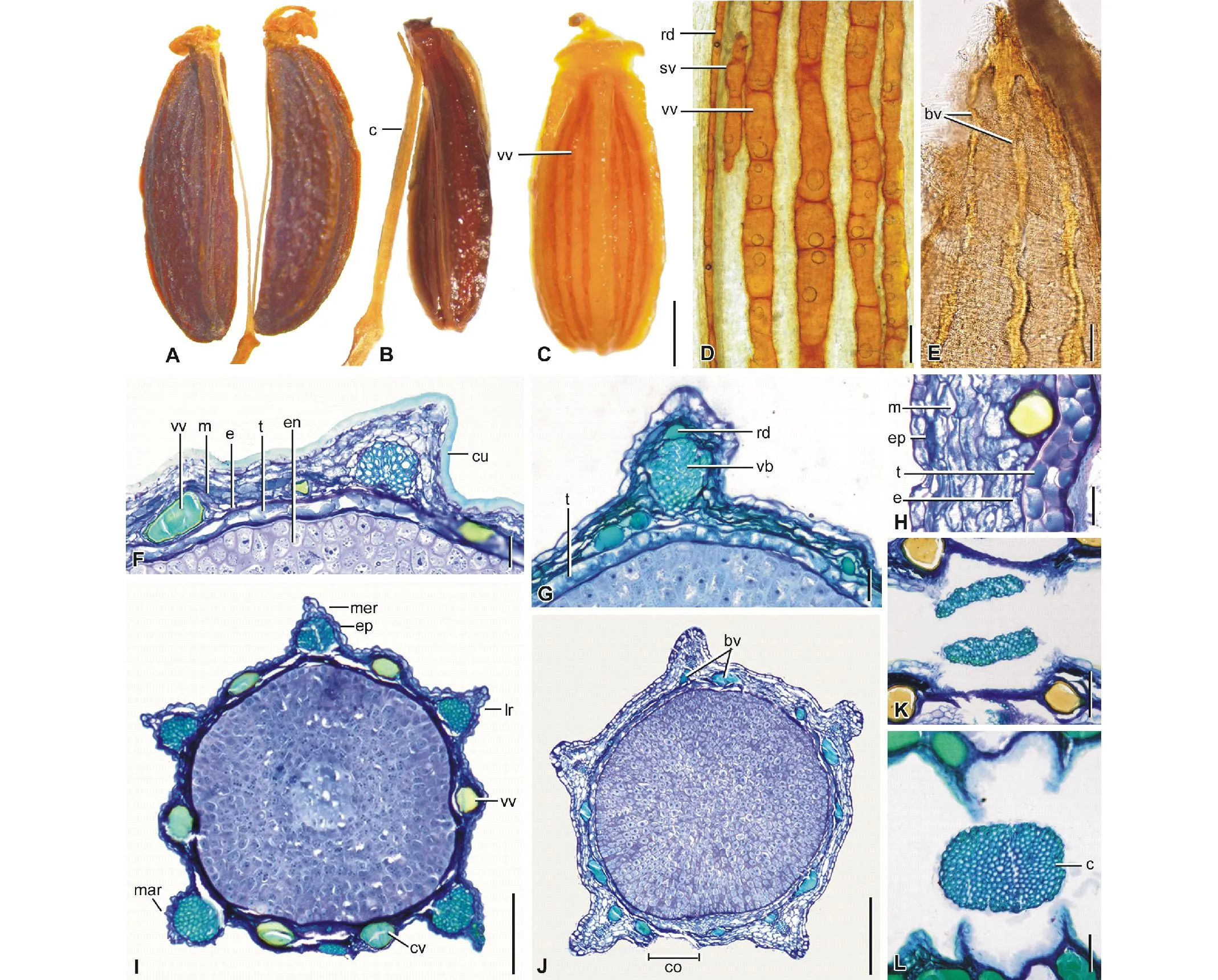
图1 柴胡属果实或果实横切面Fig.1 Fruits or their transverse sections of Bupleurum 示分果表面,表皮,中果皮,内果皮,分泌油管,种皮,结合面及心皮柄。(A,F)金黄柴胡。(B)细柄柴胡。(C)锥叶柴胡。(D)大苞柴胡。(E)红柴胡。(G)北柴胡。(H)紫花大叶柴胡。(I)小柴胡。(J)北京柴胡。(K)有柄柴胡。(L)细柄柴胡。缩写:bv=分支油管;c=心皮柄;co=结合面;cu=角质层;cv=结合面油管;e=内果皮;en=胚乳;ep=表皮;lr=侧脊;m=中果皮;mar=边脊;mer=中脊;rd=伴生(或脊中)油管;sv=短油管;vb=维管束;vv=沟中油管; t=种皮。下同。标尺:A~C=1 mm; D, E=0.1 mm; F~H, K, L=0.05 mm; I, J=0.3 mm。Showing mericarp surface, epidermis, mesocarp, endocarp, secretary oil ducts, testa, commissural side, and carpophores. (A, F) B. aureum. (B) B. gracilipes. (C) B. bicaule. (D) B. euphorbioides. (E) B. scorzonerifolium. (G) B. chinense. (H) B. longiradiatum var. porphyranthum. (I) B. tenue. (J) B. chinense f. pekinense. (K) B. petiolulatum. (L) B. gracilipes. Abbreviations: bv=branching vitta; c=carpophore; co=commissure; cu=cuticle;cv=commissural vitta; e=endocarp; en=endosperm; ep=epidermis; lr=lateral rib; m=mesocarp; mar=marginal rib; mer=median rib; rd=rib duct; sv=short vitta; vb=vascular bundle; vv=vallecular vitta; t=testa. The same below. Scale bar=1 mm in A-C; 0.1 mm in D, E; 0.05 mm in F-H, K, L; 0.3 mm in I, J.
维管束:每个分果具5个维管束(vascular bundle),分别存在于中脊,侧脊及边脊(图1I,2A)。多数物种维管束的直径小于果皮厚度,如锥叶柴胡,川滇柴胡及簇生柴胡(图2C, D, I)。少数物种,包括线叶柴胡、北柴胡、红柴胡及小柴胡的维管束大,直径等于或大于果皮的厚度(图2A, E, DD, GG)。

图2 柴胡属果实横切面Fig.2 Transverse sections of fruits of Bupleurum 示分果形状,维管束,分泌油管,胚乳,结合面及心皮柄的变化。(A)线叶柴胡。(B)金黄柴胡。(C)锥叶柴胡。(D)川滇柴胡。(E)北柴胡。(F)百花山柴胡。(G)北京柴胡。(H)紫花鸭趾柴胡。(I)簇生柴胡。(J)葡枝柴胡。(K)密花柴胡。(L)太白柴胡。(M)大苞柴胡。(N)新疆柴胡。(O)细柄柴胡。(P)纤细柴胡。(Q)长白柴胡。(R)阿尔泰柴胡。(S)抱茎柴胡。(T)空心柴胡。(U)秦岭柴胡。(V)大叶柴胡。(W)短伞大叶柴胡。(X)紫花大叶柴胡。(Y)马尔康柴胡。(Z)竹叶柴胡。(AA)马尾柴胡。(BB)有柄柴胡。(CC)丽江柴胡。(DD)红柴胡。(EE)兴安柴胡。(FF)黑柴胡。(GG)小柴胡。(HH)三辐柴胡。(II)银州柴胡。(JJ)云南柴胡。标尺=0.3 mm。Showing variation in mericarp shape, vascular bundles, secretary oil ducts, endosperm shape, commissural width and carpophores. (A) B. angustissimum. (B) B. aureum. (C) B. bicaule. (D) B. candollei. (E) B. chinense. (F) B. chinense f. octoradiatum. (G) B. chinense f. pekinense. (H) B. commelynoideum. (I) B. condensatum. (J) B. dalhousianum. (K) B. densiflorum. (L) B. dielsianum. (M) B. euphorbioides. (N) B. exaltatum. (O) B. gracilipes. (P) B. gracillimum. (Q) B. komarovianum. (R) B. krylovianum. (S) B. longicaule var. amplexicaule. (T) B. longicaule var. franchetii. (U) B. longicaule var. giraldii. (V) B. longiradiatum. (W) B. longiradiatum var. breviradiatum. (X) B. longiradiatum var. porphyranthum. (Y) B. malconense. (Z) B. marginatum. (AA) B. microcephalum. (BB) B. petiolulatum. (CC) B. rockii. (DD) B. scorzonerifolium. (EE) B. sibiricum. (FF) B. smithii. (GG) B. tenue. (HH) B. triradiatum. (II) B. yinchowense. (JJ) B. yunnanense. Scale bar=0.3 mm.
伴生油管:伴生油管(rib duct)为与维管束相伴的油管,存在于分果的脊内维管束的外侧,所研究的物种通常无伴生油管,如金黄柴胡,葡枝柴胡及密花柴胡(图2B, J, K)。少数物种,锥叶柴胡,北柴胡, 大苞柴胡及长白柴胡有直径小的伴生油管(图1D),他们可存在分果的5个脊(图2B, M),或仅1或2个脊(图2E, Q)。
非伴生油管(简称油管):非伴生油管(vitta)存在于中果皮,不与维管束相伴。研究的柴胡均具油管,油管不分支(图1D)或分支(图1E)。多数物种的油管不分支(unbranching vittae),他们规则地分布在沟里,即两个脊之间(vallecular vitta)和结合面(commissural vitta)(图1C, I, 2C)。油管数目在不同物种有变化,簇生柴胡,密花柴胡,新疆柴胡,阿尔泰柴胡,小柴胡的油管为每个沟里1个,结合面2个,每个分果共有6个油管(图2I, K, N, R, GG)。多数物种分果的油管数量多于6,如锥叶柴胡,川滇柴胡及太白柴胡,沟里油管2~3个,结合面油管2~4个,每个分果具12~14个油管(图2C, D, L)。大苞柴胡及细柄柴胡沟里的油管4~5个,结合面油管4~6个,每个分果具22~24个油管(图2M, O)。油管通常与分果的长度近似(图1C),但短油管存在于紫花鸭趾柴胡,大苞柴胡及大叶柴胡(图1D)。分支油管(branching vitta)不规则地分布在中果皮(图1E),果实横切面油管的位置不定(图1J),这些分支油管存在于线叶柴胡、北柴胡、百花山柴胡、北京柴胡、长白柴胡及红柴胡(图2A, E~G, Q, DD)。
种皮:几乎所有物种的种皮由1层薄壁细胞构成,细胞较大,长方形(图1F, G),但紫花大叶柴胡的种皮可见2层细胞(图1H)。
胚乳:胚乳(endosperm)在结合面通常平直,如葡枝柴胡,密花柴胡及太白柴胡(图2J-L),但少数物种的胚乳微凹,如川滇柴胡,丽江柴胡及云南柴胡(图2D, CC, JJ) 。
结合面,心皮柄及结晶:结合面(commissure)为2分果的结合处,所有物种具有较窄的结合面,其宽度为果宽的10%~20%。心皮柄(carpophore)通常较细(直径小于0.1 mm),在结合面2个彼此相对,各支持1个分果(图1A, K),如马尔康柴胡及竹叶柴胡(图2Y, Z)。但纤细柴胡及细柄柴胡的心皮柄较粗(直径大于 0.2 mm)(图2O, P)。细柄柴胡的心皮柄仅在顶端裂开(图1B),因而果实中部横切面显示为单心皮柄(图1L)。所有物种缺少结晶(crystal)。
3讨论
分果形状:与柴胡类似,芹亚科及变豆菜亚科许多类群(如Alepidea及Hacquetia)分果的横切面近圆形[37]。然而参棕亚科的果实明显两侧压扁,牵环花亚科除了几个属(如Bowlesia及Oschatzia)的果实略微背腹压扁外,其他的果实明显两侧压扁[42]。
表皮细胞外壁:芹亚科多数物种表皮细胞外壁平直,但突起的外壁普遍存在于变豆菜亚科(如Actinolema macrolema,Astrantia maxima,Eryngium armatum,Hacquetia epipactis及Sanicula lamelligera),Heteromorpheae(Anginon,Glia及Heteromorpha)及几个其他芹亚科类群(如Eremodaucus lehmannii,Spermolepis patens)[33]。果实表皮细胞外具厚角质层的几种柴胡均生长在较干旱的地方,可见角质层的厚度与物种生长的环境有关。
内果皮:内果皮在芹亚科多为1层薄壁细胞构成[1],然而这一亚科的基础类群,如非洲的Heteromorpheae(Anginon swellendamense,Glia prolifera)及Lichtensteinia lacera的内果皮轻微木化。这种轻微木化的内果皮还存在变豆菜亚科及相关类群(如Eryngium planum,Sanicula lamelligera,Arctopus echinatus,Polemanniopsis marlothii及Steganotaenia araliacea)[33,37,43]。参棕亚科和牵环花亚科的内果皮由多层木化细胞构成[1,33]。
伴生油管及非伴生油管:分泌油管在Baumann[44],Tikhomitov[45]及Eyde和Tseng[46]的系统学研究起着重要的作用。芹亚科许多类群无明显的伴生油管,然而伴生油管普遍存在于芹亚科基础类群(Heteromorpheae,Annesorhiza及Chamarea),变豆菜亚科,参棕亚科及牵环花亚科,其中变豆菜亚科及相关类群(如Sanicula,Alepidea,Polemanniopsis,Steganotaenia,Marlothiella及Lichtensteinia)的伴生油管明显较大[30,33,37-38,43,47-48]。芹亚科许多类群(如Angelica及Peucedanum)不分支油管在沟里可多于1个,结合面多于2个,然而Heteromorpheae,Annesorhiza及Chamarea的油管数通常沟里只有1个,结合面2个[33,47-49]。分支油管存在于牵环花亚科(Dickinsia,Hermas,Klotzschia),变豆菜亚科及相关类群(Alepidea,Sanicula,Eryngium及Steganotaenia)[37-38,48],靠近Heteromorpheae的马达加斯加特有物种Pseudocarpum laxiflorum,然而在芹亚科若果实中油管分支,总是与不分支油管同时存在(如Ferula)。
结晶:草酸钙单晶或晶簇出现在五加科(Araliaceae)的一些薄壁组织[50],单晶存在于参棕亚科和牵环花亚科紧挨内果皮的中果皮[1,30,37-38,43,51],但不存在于变豆菜亚科及芹亚科。Burtt[52]指出芹亚科如果有结晶,只发生在结合面。然而我们观察到芹亚科所有基础类群(Heteromorpheae,Annesorhiza,Chamarea,Lichtenisteinia及Marlothiella)均具有晶簇。此外晶簇还普遍存在在变豆菜亚科及相关类群(Steganotaenia及Polemanniopsis)及牵环花亚科(如Huanaca andina及Bolax caespitosus)的中果皮,但芹亚科其他类群多无结晶[35,38,47]。
种皮:种皮被认为是有价值的分类特征[53-55]。多数芹亚科的种皮由1层细胞构成,但我们观察到多于1层细胞的种皮存在于变豆菜亚科(如Alepidea natalensis及Eryngium hookeri),非洲的Heteromorpheae(如Anginon pumilum),Annesorhiza(如Annesorhiza wilmsii),Chamarea(如Chamarea capensis),及Itasina(Itasina filifolia)。Magee等[30]的研究显示Itasina与Annesorhiza的亲缘关系较近。此外多于1层细胞的种皮还存在于Bupleirumbaldense(产于不列颠群岛),B.distichophyllum(产于印度)及B.rigidum(产于阿尔及利亚)(未发表)。王艳杰和申家恒[56]指出北柴胡具一层珠被,然而这个属的其他物种的珠被结构尚不知晓。
心皮柄:心皮柄来源于腹束,连同周围的非维管组织一起支持着心皮[57]。芹亚科多数物种具2个心皮柄,类似于细柄柴胡(图1B)的一个顶端分裂的心皮柄在芹亚科见于Eremodaucus及Scandix(未发表),但广泛存在于牵环花亚科(如Bolax,Dickinsia,Hermas及Huanaca)及变豆菜亚科(Alepidea及Arctopus)[40]。
叶及导管分子:单叶是参棕亚科,牵环花亚科,变豆菜亚科及柴胡属的主要特征[1,40],但亦存在于Heteromorpheae(Anginon及Heteromorpha)[58]。同伞形科木本类群一样柴胡属的导管分子通常短,具单穿孔板[59-62],此外柴胡属及非洲的Heteromorpha,Anginon,Glia及Polemannia的导管具螺纹增厚[60,62]。Stepanova和Oskolski[5]指出导管壁螺纹增厚应视为芹亚科的原始类型。
柴胡属果实的多数特征同样存在于芹亚科的其他类群(如横切面近圆形,中果皮有伴生油管及规则分布的非伴生油管。中果皮无结晶。心皮柄2个),然而有些特征(如油管呈分支状,种皮由2层细胞构成)仅见于芹亚科的基础类群及伞形科的其他亚科,因而果实结构的研究为分子系统学将柴胡属放在靠近芹亚科的基础类群,即这一亚科其他类群的基础位置提供了形态学依据,进而为进一步研究芹亚科系统学提供了果实形态学基础。
References:
[1]Drude O. Umbelliferae. In: Engler A, Prantl K. Die Natürlichen Pflanzenfamilien[M]. Leipzig: Wilhelm Engelmann, 1898.
[2]Pimenov M G, Leonov M V. The Genera of the Umbelliferae: A Nomenclator[M]. London: Royal Botanic Gardens, Kew, 1993.
[3]Editorial Committee of Flora of Chinese China Academy of Sciences. Flora Repubulicae Popularis Sinicae (Volume fifty-fifth, The third section)[M]. Beijing: Science Press, 1992.
[4]She M L, Pu F T, Pan Z H,etal. Apiaceae. In: Flora of China Editorial Committee. Flora of China (14)[M]. St. Louis, Missouri: Missouri Botanical Garden Press, 2005.
[5]Stepanova A V, Oskolski A A. Wood anatomy ofBupleurumL. (Apioideae, Apiaceae) in relation to habit, phylogenetic relationships, and infrageneric taxonomy. Plant Diversity and Evolution, 2010, 128(3-4): 501-516.
[6]Plunkett G M, Chandler G T, Lowry P P II,etal. Recent advances in understanding Apiales with a revised classification. South African Journal of Botany, 2004, 70(3): 371-381.
[7]Cerceau-Larrival M T. Plantules et pollen d’Ombellifères. Mémoires du Muséum National d'Histoire Naturelle, 1962, 14: 1-166.
[8]Cerceau-Larrival M T. Morphologie pollinique et corrèlations phylogènetiques chez les ombellifères. In: Heywood V H. The Biology and Chemistry of the Umbelliferae[M]. Pittsburgh: Academic Press, 1971.
[9]Shu P, She M L. Pollen Photographs and Flore of Umbelliferae in China[M]. Shanghai: Shanghai Science & Technology Publishers, 2001.
[10]Cauwet-Marc A M. Races chromosomiques, écologie et biologie duBupleurumranunculoidesL. dans la parties orientale des Pyénées. Bulletin de la Société Botanique de France, 1970, 117: 373-384.
[11]Cauwet-Marc A M. Connaissances caryologiques actuelles sur le genreBupleurumL. (Umbelliferae): nombres chromosomiques et numbers de base. Bulletin of the British Museum-Natural History, 1979, 3: 191-211.
[12]Cauwet-Marc A M. Contribution de la caryologie á la connaissance de la systèmatique et de la phylogènie du genreBupleurumL. Candollea, 1979, 34: 49-86.
[13]Wang Q Z, He X J, Zhou S D,etal. Phylogenetic inference of the genusBupleurum(Apiaceae) in Hengduan Mountains based on chromosome counts and nuclear ribosomal DNA ITS sequences. Journal of Systematics and Evolution, 2008, 46(2): 142-154.
[14]Downie S R, Katz-Downie D S. A molecular phylogeny of Apiaceae subfamily Apioideae: evidence from nuclear ribosomal DNA internal transcribed spacer sequences. American Journal of Botany, 1996, 83(2): 234-251.
[15]Kondo K, Terabayashi S, Okada M,etal. Phylogenetic relationship of medicinally importantCnidiumofficinaleand Japanese Apiaceae based on rbcL sequences. Journal of Plant Research, 1996, 109(1): 21-27.
[16]Plunkett G M, Soltis D E, Soltis P S. Higher level relationships of Apiales (Apiaceae and Araliaceae) based on phylogenetic analysis of rbcL sequences. American Journal of Botany, 1996, 83(4): 499-515.
[17]Plunkett G M, Soltis D E, Soltis P S. Evolutionary patterns in Apiaceae: inferences based on matK sequence data. Systematic Botany, 1996, 21(4): 477-495.
[18]Plunkett G M, Soltis D E, Soltis P S. Clarification of the relationships between Apiaceae and Araliaceae based on matK and rbcL sequence data. American Journal of Botany, 1997, 84(4): 565-580.
[19]Downie S R, Katz-Downie D S, Cho K J. Phylogenetic analysis of Apiaceae subfamily Apioideae using nucleotide sequences from the chloroplast rpoC1 intron. Molecular Phylogenetics and Evolution, 1996, 6(1): 1-18.
[20]Downie S R, Ramanath S, Katz-Downie D S,etal. Molecular systematics of Apiaceae subfamily Apioideae: phylogenetic analyses of nuclear ribosomal DNA internal transcribed spacer and plastid rpoC1 intron sequences. American Journal of Botany, 1998, 85(4): 563-591.
[21]Downie S R, Katz-Downie D S, Spalik K. A phylogeny of Apiaceae tribe Scandiceae: evidence from nuclear ribosomal DNA internal transcribed spacer sequences. American Journal of Botany, 2000, 87(1): 78-95.
[22]Downie S R, Katz-Downie D S, Watson M F. A phylogeny of the flowering plant family Apiaceae based on chloroplast DNA rpl16 and rpoC1 intron sequences: towards a suprageneric classification of subfamily Apioideae. American Journal of Botany, 2000, 87(2): 273-292.
[23]Downie S R, Watson M F, Spalik K,etal. Molecular systematics of Old World Apioideae (Apiaceae): relationships among some members of tribe Peucedaneae sensu lato, the placement of several island-endemic species, and resolution within the apioid superclade. Canadian Journal of Botany, 2000, 78(4): 506-528.
[24]Downie S R, Plunkett G M, Watson M F,etal. Tribes and clades within Apiaceae subfamily Apioideae: the contribution of molecular data. Edinburgh Journal of Botany, 2001, 58(2): 301-330.
[25]Valiejo-Roman K M, Pimenov M G, Terentieva E I,etal. Molecular systematics of the Umbelliferae: using nuclear ribosomal DNA internal transcribed spacer sequences to resolve issues of evolutionary relationships. Botanicheskii Zhurnal, 1998, 83(7): 1-22.
[26]Katz-Downie D S, Valiejo-Roman C M, Terentieva E I,etal. Towards a molecular phylogeny of Apiaceae subfamily Apioideae: additional information from nuclear ribosomal DNA ITS sequences. Plant Systematics and Evolution, 1999, 216(3): 167-195.
[27]Plunkett G M, Downie S R. Major lineages within Apiaceae subfamily Apioideae: a comparison of chloroplast restriction site and DNA sequence data. American Journal of Botany, 1999, 86(7): 1015-1026.
[28]Downie S R, Katz-Downie D S. Phylogenetic analysis of chloroplast rps16 intron sequences reveals relationships within the woody southern African Apiaceae subfamily Apioideae. Canadian Journal of Botany, 1999, 77(8): 1120-1135.
[29]Plunkett G M. Relationship of the order Apiales to subclass Asteridae: a re-evaluation of morphological characters based on insights from molecular data. Edinburgh Journal of Botany, 2001, 58(2): 183-200.
[31]Neves S S, Watson M F. Phylogenetic relationships inBupleurum(Apiaceae) based on nuclear ribosomal DNA ITS sequence data. Annals of Botany, 2004, 93(4): 379-398.
[32]Snogerup S, Snogerup B.BupleurumL. (Umbelliferae) in Europe-1. The annuals, B. sect.Bupleurumand sect.Aristata. Willdenowia, 2001, 31(2): 205-308.
[33]Liu M. A taxonomic Evaluation of Fruit Structure in the Family Apiaceae[D]. Johannesburg: Rand Afrikaans University, 2004.
[34]Wang C B, Ma X G, He J X. Fruit features of someBupleurumspecies (Apiaceae) and their systematic implication. Plant Science Journal, 2011, 29(4): 399-408.
[35]Heywood V H, Dakshini K M M. Fruit structure in the Umbelliferae-Caucalideae. The biology and chemistry of the Umbelliferae. Botanical Journal of the Linnean Society, 1971, 64(Suppl.1): 215-232.
[36]Wang Y T, Liu M, Cheng X Y. Trichomes and fruit structures of Angelica and related genera of the family Apiaceae and their taxonomic significance. Acta Prataculturae Sinica, 2015, 24(4): 66-74.
[37]Liu M, Van Wyk B E, Tilney P M. The taxonomic value of fruit structure in the subfamily Saniculoideae and related African genera (Apiaceae). Taxon, 2003, 52(2): 261-270.
[38]Liu M, Van Wyk B E, Tilney P M,etal. Evidence from fruit structure supports in general the circumscription of Apiaceae subfamily Azorelloideae. Plant Systematics and Evolution, 2009, 280(1-2): 1-13.
[39]Liu M, Plunkett G M, Lowry II P P. Fruit anatomy provides structural synapomorphies to help define Myodocarpaceae (Apiales). Systematic Botany, 2010, 35(3): 675-681.
[40]Liu M, Plunkett G M, Van Wyk B E,etal. The phylogenetic significance of the carpophore in Apiaceae. Annals of Botany, 2012, 110(8): 1531-1543.
[41]Feder N, O’Brien T P. Plant microtechnique: some principles and new methods. American Journal of Botany, 1968, 55(1): 123-142.
[42]Liu M, Plunkett G M, Van Wyk B E,etal. The taxonomic value of winged fruits in the order Apiales. American Journal of Botany, 2006, 93(9): 1357-1368.
[43]Liu M, Van Wyk B E, Tilney P M. Ontogeny of the fruits of two anomalous African woody generaPolemanniopsisandSteganotaenia(Apiaceae) and their phylogenetic relationship. Edinburgh Journal of Botany, 2003, 60(3): 249-257.
[44]Baumann M. Myodocarpus und die phylogenie der umbelliferenfrucht-frucht. Berichte der Schweizerischen Botanischen Gesellschaft, 1946, 56: 13-112.
[45]Tikhomitov V N. One the systematic position of the genusHydrocotyleL. andCentellaL. Ememd Urban. Botanicheskii Zhurnal, 1961, 46: 584-586.
[46]Eyde R H, Tseng C C. What is the primitive floral structure of Araliaceae. Journal of Arnold Arboretum, 1971, 52(2): 205-239.
[47]Liu M, Van Wyk B E, Tilney P M. A revision of the genusMarlothiella(Apiaceae). South African Journal of Botany, 2007, 73(2): 209-213.
[48]Liu M, Van Wyk B E, Tilney P M. Irregular vittae and druse crystals in Steganotaenia fruit support a taxonomic affinity with the subfamily Saniculoideae (Apiaceae). South African Journal of Botany, 2007, 73(2): 252-255.
[49]Liu M, Van Wyk B E, Tilney P M. A revision of the genusChoritaenia(Apiaceae). South African Journal of Botany, 2007, 73(2): 184-189.
[50]Cronquist A. An Integrated System of Classification of Flowering Plants[M]. New York: Columbia University Press, 1981.
[51]Liu M, Van Wyk B E, Tilney P M. The taxonomic value of fruit structure in the Chinese endemic genusDickinsia(Apiaceae). Nordic Journal of Botany, 2002, 22(5): 603-607.
[52]Burtt B L. Umbelliferae of southern Africa: an introduction and annotated checklist. Edinburgh Journal of Botany, 1991, 48(2): 135-275.
[53]Vaughan J G, Whitehouse J M. Seed structure and the taxonomy of the Cruciferae. Botanical Journal of the Linnean Society, 1971, 64(4): 383-409.
[54]Bouman F. Integument initiation and testa development in some Cruciferae. Botanical Journal of the Linnean Society, 1975, 70(3): 213-229.
[55]Zeng C L, Wang J B, Liu A L,etal. Seed coat microsculpturing changes during seed development in diploid and amphidiploidBrassicaspecies. Annals of Botany, 2004, 93(5): 555-566.
[56]Wang Y J, Shen J H. Megasporogenesis, microsporogenesis and development of female and male gametophyte ofBupleurumchinense. Chinese Bulletin of Botany, 2007, 24(3): 425-432.
[57]Jackson G. A study of the carpophore of the Umbelliferae. American Journal of Botany, 1933, 20(2): 121-144.
[58]Van Wyk B E. Apiaceae. In: Leistner O A. Seed Plants of Southern Africa: Families and Genera[M]. Pretoria: National Botanical Institute, 2000.
[59]Metcalfe C R, Chalk L. Anatomy of the Dicotyledons, Vol. 2[M]. Oxford: Clarendon Press, 1950.
[60]Rodriguez R L. Systematic anatomical studies onMyrrhidendronand other woody Umbellales. University of California Publications in Botany, 1957, 29(2): 145-318.
[61]Oskolski A A. Systematic and phylogenetic wood anatomy of Apiales. Edinburgh Journal of Botany, 2001, 58(2): 201-206.
[62]Oskolski A A, Van Wyk B E. Systematic and phylogenetic value of wood anatomy in Heteromorpheae (Apiaceae, Apioideae). Botanical Journal of the Linnean Society, 2008, 158(4): 569-583.
参考文献:
[3]中国科学院中国植物志编辑委员会. 中国植物志(第55卷,第1分册)[M]. 北京: 科学出版社, 1992.
[9]舒璞, 佘孟兰. 中国伞形科植物花粉图志[M]. 上海: 上海科技出版社, 2001.
[34]王长宝, 马祥光, 何兴金. 伞形科柴胡属部分物种的果实特征及系统学意义. 植物科学时报, 2011, 29(4): 399-408.
[36]王宇婷, 刘玫, 程薪宇. 中国伞形科当归属及相关类群毛状体和果实结构的系统学价值. 草业学报, 2015, 24(4): 66-74.
[56]王艳杰, 申家恒. 柴胡大、小孢子发生及雌、雄配子体发育. 植物学通报, 2007, 24(3): 425-432.
http://cyxb.lzu.edu.cn
常修奇,刘玫,程薪宇,王宇婷,魏雪莹,王雪微. 中国伞形科柴胡属果实结构的系统学价值.草业学报, 2015, 24(10): 108-119.
CHANG Xiu-Qi, LIU Mei, CHENG Xin-Yu, WANG Yu-Ting, WEI Xue-Ying, WANG Xue-Wei. The phylogenetic significance of fruit structures inBupleurumof the family Apiaceae. Acta Prataculturae Sinica, 2015, 24(10): 108-119.
通讯作者*Corresponding author.E-mail: m.r.liu@126.com
作者简介:常修奇(1991-),男,黑龙江双鸭山人,在读硕士。E-mail: goalhia420@163.com
基金项目:国家自然科学基金(31270235和31070169)资助。
收稿日期:2015-01-29;改回日期:2015-04-30
DOI:10.11686/cyxb2015067
