稻米垩白QTL定位分析
2016-01-16张大双吴健强黄培英王际凤朱速松
彭 强, 张大双, 吴健强, 刘 颖, 黄培英, 王际凤, 朱速松
(贵州省水稻研究所, 贵阳550006)
垩白是指稻米胚乳中白色不透明的部分,是由于胚乳中淀粉的不正确排列而造成的复杂表型,通常以垩白粒率、垩白大小和垩白度来表示。垩白粒率偏高、垩白度偏大是长期困扰我国优质稻米生产的最主要障碍,其中南方稻区早籼稻尤为突出[1]。因此,培育低垩白水稻品种仍是水稻品质育种(尤其是籼稻)的重要方向之一。
研究表明,稻米垩白形状是受多基因控制的、复杂的数量性状。学者们利用多种群体(BIL、CSSL、RIL、DH、IL等)进行垩白性状QTL分析,共发现有62个垩白性状QTL分布在32个位点上[2-3]。与此同时,一些与垩白表型有关的基因也被成功克隆,如位于第5染色体的 OsPPDKB 基因[4]和 Chalk 5基因[5]、第8染色体上的SSIIIa基因[6]和对稻米垩白粒率具有一因多效作用的GW 2基因[7]。虽然垩白性状的遗传关系复杂,且稻米垩白的形成易受环境影响,但存在一些成簇分布的垩白性状主效QTL能在不同群体和环境下稳定表达[8-9]。为 此,本研 究以广亲和性爪 哇稻CPSLO 17(低垩白)和配合力强籼稻 V 20 B(高垩白)作为亲本构建新的籼爪交遗传背景RIL,以稻米垩白度为表型数据,结合高密度SLAF标签连锁图谱,进行稻米垩白性状QTL定位及其遗传效应分析,旨在为米垩白性状QTL的精细定位和控制垩白基因克隆提供新素材,也为分子标记辅助选择(MAS)培育低垩白水稻品种提供理论基础。

表1 稻米垩白QTL定位及其遗传效应
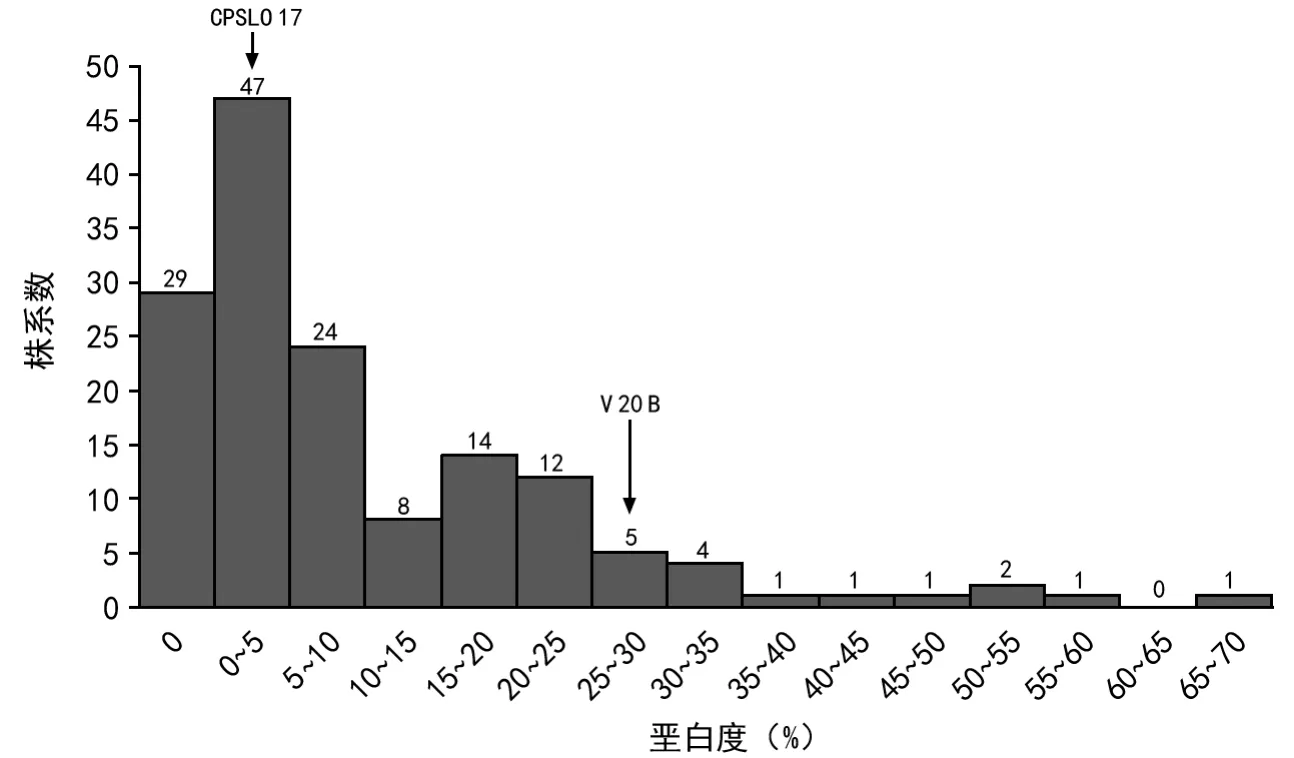
图1 RIL群体的稻米垩白度分布
1 材料与方法
1.1 供试材料
V 20 B/CPSLO 17重组自交群体由F1代通过单粒传方法得到的。亲本和RIL家系种植在贵州省水稻研究所(贵阳)试验田,每个材料种植20株,成熟时混收,晒干保存,共收2份亲本和150份RIL材料,用于稻米垩白性状QTL分析。
1.2 稻米垩白度测定
对于每份供试样品,随机挑选50粒精米置于玻璃板上,目测挑选出有阴影的米粒,并记录米粒数量,计算出该样品的垩白率(%);随机挑选10粒有垩白的米粒,逐粒目测垩白面积占整个米粒面积的百分率并取平均值,即为该样品的垩白大小;该样品的垩白度(%)为垩白率(%)与垩白大小的乘积。
1.3 QTL分析
V 2 0 B/CPSLO 1 7重 组 自 交 家 系 的SLAF标签的分子数据(未发表)是由北京百迈客生物科技有限公司利用SLAF-seq(Specific-Locus Amplified Fragment Sequencing)技术[10]和 High Map 软件[11]开发获得的。总共有8 602个高质量SLAF标签,比较均匀分布在12条染色体上。该图谱覆盖水稻全基因组2 508.65 c M,标记间平均距离为0.292 c M。采用软件 Map QTL 5 的 Internal Mapping 方法进行垩白性状QTL分析,扫描步长设定为默认值1.0 c M ,LOD值设定为默认值3.9,并计算每个 QTL对苗期耐冷性的贡献率和加性效应,QTL的命名原则遵循McCouch等[12]提出的方法。加性效应为正值表示增效等位基因来源于高垩白亲本V 20 B,为负值表示来源于低垩白亲本CPSLO 17。
2 结果与分析
2.1 亲本和重组自交群体的稻米垩白表型数据
每份材料收种保存3月后进行稻米垩白度测定,亲本V 20 B稻米品质差,垩白多,其垩白度为27.5%(图1右箭头);亲本CPSLO 17稻米品质优良,其垩白度为4.29%(图1左箭头)。对150份 V 20 B/CPSLO 17重组自交群体(RILs)进行稻米垩白度测定,稻米垩白度大小分布如图1所示。从图1可知,RILs的150个家系的稻米垩白度在0~70%之间连续分布(图1),群体平均垩白度为10.02%。垩白性状表现出受多基因控制的数量性状遗传特征。
2.2 水稻稻米垩白性状QTL分析
采用软件 MapQTL 5的Internal Mapping方法对稻米垩白性状进行QTL分析,在第5染色体上共检测到连续排列的4个稻米垩白性状QTL位点,分别命名为qC-5 a、qC-5 b、qC-5 c和qC-5 d,对应的连锁群位置及其遗传效应如表1所示。4个垩白性状QTL位点的LOD值分别为4.02、4.09、3.94和4.1,对表型变异的解释率分别为11.6%、11.8%、11.2%和11.8%,且这4个QTL抗性等位基因都来自高垩白亲本V 20 B(表1)。

图2 稻米垩白性状QTL在chr 5染色体上的分布
3 结果与讨论
垩白的形成是一个复杂的生理过程,涉及多种代谢途径、物质运输过程和不同类型的酶;此外,稻米垩白是复杂的数量遗传性状,且受与各种环境因素相互作用的多基因系统调控。目前有关稻米垩白的研究进展主要集中在垩白的理化特性描述和突变体基因的鉴定方面,Chalk 5基因[5]是已知的唯一一个通过 QTL精细定位克隆到的控制稻米垩白的QTL基因,因此揭示垩白形成分子机理及其调控网络仍有大量工作需要完成。本研究利用V 20 B/CPSLO 17的RIL群体从分子水平对稻米垩白度QTL进行定位分析,在第5染色体上检测到4个连续排列的垩白性状相关QTL(qC-5 a、qC-5 b、qC-5 c 和qC-5 d)。在 RGAP网站(http://rice.plantbiology.msu.edu/)进行 blast比对分析发现,Chalk 5基因与本研究的4个QTL位点不同。该4垩白QTL在遗传图谱上距离近,各自的LOD值和贡献率也相差不大,说明上述4个垩白QTL基因很可能是以基因簇形式存在并参与垩白调控。本研究利用的2个亲本的表型差异明显,且RIL群体垩白度是连续性分布的,也表现出受多基因控制的数量性状遗传特征。这些QTL的发现和定位为克隆分离稻米垩白性状相关基因提供材料和奠定基础,也为利用分子标记辅助选择培育低垩白水稻品种提供理论依据。
[1]罗玉坤,朱智伟,陈能,等.中国主要稻米的粒型及其品质特性[J].中国水稻科学,2004,18(2):49-53.
[2]郭涛.水稻垩白材料CSSL 50的形态学解析和 QTLqPGWC-8的精细定位[D].南京农业大学博士论文,2011:14-16.
[3]周立军,江玲,刘喜,等.水稻千粒重和坚白粒率的QTL及其互作分析[J].作物学报,2009,35(2):69-75.
[4]Kang HG,Park SH,Matsuoka M,et al.White-core endosperm floury endosperm-4 in rice is generated by knockout mutations in the C 4-type pyruvate orthophosphate dikinase gene(OsPPDKB)[J].Plant J,2005,42(6):901-911.
[5]Li YB,Fan CC,Xing YZ,et al.Chalk 5 encodes a vacuolar H+-translocating pyrophosphatase influencing grain chalkiness in rice[J].Nature Genetics,2014,46(4):398-404.
[6]Fujita N,Yoshida M,Kondo T,et al.Characterization of SSIIIa-deficient mutants of rice:The function of SSIIIa and pleiotropic effects by SSIIIa deficiency in the rice endosperm[J].Plant Physiol,2007,144(4):2 009-2 023.
[7]Song XJ,Huang W,Shi M,et al.A QTL for rice grain width and weight encodes a previously un-known RING-type E 3 ubiquitin ligase[J].Nat Genet,2007,39(5):623-630.
[8]万向元.水稻品种性状QTL表达稳定性分析与精细定位[D].南京农业大学博士论文,2005:78-87.
[9]张大双,黄培英,彭强,等.稻米垩白性状的 QTL分析[J].种子,2013,32(10):25-32.
[10]Sun X,Liu D,Zhang X,et al.SLAF-seq:an efficient method of large-scale De novo SNP discovery and genotyping using high-throughput sequencing[J].PLoS One,2013,8(3):e 58 700.
[11]Liu D,Ma C,Hong W,et al.Construction and Analysis of High-Density Linkage Map Using High-Throughput Sequencing Data[J].PLoS One,2014,9(6):e 98 855.
[12]MaCouch S R,Cho Y G,Yang M,et al.Blinstrub M,Morishima H,Kinoshita T.Report on QTL nomenclature[J].Rice Genet Newslett,1997,14:11-13.
