温度、饵料种类对中华哲水蚤(Calanus sinicus)油脂积累与生长发育的影响*
2016-01-15周孔霖
周孔霖 孙 松①
(1. 中国科学院海洋研究所海洋生态与环境科学重点实验室 青岛 266071; 2. 中国科学院大学 北京 100049; 3. 中国科学院海洋研究所山东胶州湾海洋生态系统国家野外科学观测研究站 青岛 266071; 4. 青岛海洋科学与技术国家实验室海洋生态与环境科学功能实验室 青岛 266071)
中华哲水蚤(Calanus sinicus)是我国近海浮游动物优势种, 是黄海大型桡足类功能群的代表, 其种群的动态变化将影响海洋生态系统中物质与能量的流动(Sun et al, 2010)。夏季, 黄海陆架区中华哲水蚤种群以 C5期幼体为主体迁移至黄海冷水团内度夏(Pu et al, 2004)。度夏期间, C5期幼体处于静息状态, 个体呼吸率低、蜕皮发育受到抑制、垂直移动减弱, 仅靠储存的油脂维持基础代谢(Li et al, 2004; Pu et al,2004; Zhou et al, 2016)。春末, 黄海冷水团区的中华哲水蚤在油囊内积累大量油脂, 其积累程度关系到中华哲水蚤能否顺利度夏(孙松等, 2011)。此外, “油脂积累假说”(lipid accumulation window hypothesis)指出当油脂积累超过某个阈值时哲水蚤会启动休眠(Rey-Rassat et al, 2002), 故中华哲水蚤前期油脂的积累可能是诱导度夏启动的内源性因子(孙松等, 2011)。目前, 哲水蚤大体积油囊形成的原因尚不明确。哲水蚤在休眠前存在能量分配问题, 即选择将摄食所获取的能量用于油脂积累亦或是蜕皮发育(Ji, 2011)。油脂的积累受到多种环境因素的影响(如食物、温度、摄食压力等), 也存在内源因素的调节(Johnson et al,2008)。
春末, 随着温跃层(11—22°C)逐步形成, 冷水团(<10°C)在黄海中部陆架区初见雏形并盘踞在温跃层以下(Hu et al, 2004)。与此同时, 黄海冷水团区中华哲水蚤 C5期幼体积累大量油脂并储存在油囊中, 以备度夏所需; 而近岸种群油脂积累少, 个体活跃(孙松等, 2011)。中华哲水蚤C5期幼体和雌体存在昼夜垂直移动习性, 即夜间上升到温跃层进行摄食活动,日间返回底层躲避捕食者(Huanget al, 1993)。在冷水团区, 中华哲水蚤通过昼夜垂直移动能感受到一定的温差(1—12°C), 同时底层冷水有利于个体通过降低代谢消耗来增加能量的积累。相反, 黄海浅水区水体垂直混合较好, 表底层温差小。由此推测, 温差和冷水团的低温环境有利于C5期幼体油脂的积累。
饵料也是影响哲水蚤油脂积累的重要因素之一。在高饵料浓度培养下, 飞马哲水蚤(C. finmarchicus)能积累更多的油脂(Rey-Rassatet al, 2002)。然而在黄海, 中华哲水蚤C5期幼体的油囊体积与叶绿素a浓度之间无显著关联(孙松等, 2011)。叶绿素a浓度只能粗略反映浮游植物的丰度, 无法说明浮游植物的种类和粒径大小。由此推测, 饵料浓度不是中华哲水蚤大体积油囊形成的关键因素。此外, 中华哲水蚤是杂食性种类, 能选择性摄食纤毛虫等微型浮游动物(Huoet al, 2008)。黄海纤毛虫的生物量在春夏季达到高值(赵楠, 2008), 但动物性饵料对油脂积累的影响尚不明确。
鉴于油脂积累影响因素的复杂性, 本文从温度(恒温、变温培养)和饵料种类(硅藻、自然饵料)两个角度设计了双因子培养实验, 旨在探讨温度和饵料种类对中华哲水蚤生长发育以及C5期幼体油脂积累的影响, 初步探讨中华哲水蚤度夏前大体积油囊形成的原因和影响因素。
1 材料与方法
1.1 实验设计
春末, 黄海冷水团区表、底层的温差(1—12°C)和冷水团的低温环境(<10°C)可能有利于 C5期幼体油脂的积累。中华哲水蚤最适生长温度的上限为20°C(蒲新明, 2003)。为研究温度在C5期幼体油脂积累中的影响作用, 本实验设置两个恒温组(10°C和19°C)以及两个变温组(10—19°CⅠ和 10—19°CⅡ)以探讨昼夜温差、温度变化强度对油脂积累的影响。10—19°CⅠ组温度变化周期为12h(10°C)∶12h(19°C),升降温在2小时内完成。10—19°CⅡ组温度变化周期为 12h(10°C)∶12h(升温/降温)∶12h(19°C), 升降温在12小时内完成。春季, 黄海冷水团区局部叶绿素a浓度超过4mg/m3(Wanget al, 2009)。根据经验系数1∶50换算, 该叶绿素a浓度相当于0.2μgC/mL的碳浓度(Atkinson, 1996)。为保证饵料充足, 本实验饵料浓度设置为0.2μgC/mL。每个温度组下设置硅藻饵料和自然饵料两个处理。哲水蚤油脂积累主要发生在C3—C5期(Leeet al, 2006), 故选择中华哲水蚤C3期幼体作为本实验的培养对象。每个温度饵料组有 30只中华哲水蚤C3期幼体。
1.2 硅藻饵料
实验选取2种常见硅藻作为饵料, 即中肋骨条藻(Skeletonema costatum)和海链藻(Thalassionsirasp.)。实验藻种由中国科学院海洋研究所海洋生态毒理实验室提供, 均采自青岛胶州湾海域。硅藻在加硅酸盐的 f/2 培养液、盐度(30±0.5)、温度(18±0.5)°C、光周期 12L∶12D的条件下培养。每日用库尔特计数器(Coulter counter, Multisizer 3)检测新鲜藻液的密度,并绘制藻类的生长曲线。取对数生长期的藻液, 根据藻体积换算碳含量, 将2种硅藻按碳浓度进行等比例混合后作为中华哲水蚤的硅藻饵料。硅藻的特征参数见表1。

表1 实验藻种特征Tab. 1 Characteristics of the phytoplankton species
1.3 自然饵料的获取
自然饵料取自中华哲水蚤的采样点, 每 3—4日采集1次。用孔径为20μm的筛绢网具拖取水下5米到表层的网样, 存放于3—4个2L样品瓶中, 置于保温桶中暂存。回实验室后, 样品暂存于 10°C培养箱中。每日取混合均匀的7.5mL网样3份并用1%Lugol’s试剂固定, 于倒置显微镜(Olympus IX51)下分类并测量。根据硅藻、甲藻、微型浮游动物的体积估算碳含量, 从而估计水样中饵料生物的碳浓度。自然饵料中,硅藻的碳含量占总体的 95.2%—98.2%, 甲藻为0.3%—1.5%, 微型浮游动物为 1.2%—3.4%。自然饵料中的优势种为角毛藻(Chaetocerossp.)和骨条藻(Skeletonemasp.)。
1.4 中华哲水蚤的采集
实验所用的中华哲水蚤是2014年4月于青岛港码头(36°04′N, 120°19′E)用浅水Ⅱ型浮游生物网(网口直径0.32m, 筛绢孔径160μm)进行垂直拖网获得, 现场水温 11.5°C。样品暂存于保温桶(20L)中, 并于 1小时内转移至实验室。通过肉眼挑选大小相似的、健康活跃的中华哲水蚤幼体, 并在解剖镜下确认其发育阶段。将 C3期幼体暂养于预冷的过滤海水(11°C)中, 以备后期的培养实验所需。另随机挑选C3、C4、C5期幼体和雌体各 20只, 测量后保存于 5%福尔马林海水溶液中。
实验开始前, 将C3期幼体随机转移到8个2L烧杯中, 每个烧杯约30只。根据实验温度设置, 将烧杯置于 4个培养箱(江南 SPX-270)中, 初始温度设置为11°C。适应1天后, 根据实验设置每小时将培养箱调整1°C直至预定温度。实验开始后, 每两天换水1次,并根据实验设置更换饵料。实验所用的过滤海水均经过 0.45μm混合膜过滤并充分曝气, 随后存放于各培养箱中预冷, 以保证换水时不产生温差。所有个体发育成熟后结束实验, 10°C组培养 18天, 其余实验组15天。
1.5 测量与计算
每隔两天在解剖镜(Nikon SMZ645)下对中华哲水蚤进行测量并确认发育期, 随后放回烧杯中继续培养, 操作过程约1min。测量指标包括: 前体部体长(PL)、体宽(PW)、油囊长(OL)、油囊宽(OW)。油囊体积大小可用于表示油脂积累的程度。根据Svetlichny等(2006)的方法计算中华哲水蚤前体部的体积(prosome volume, PV)和油囊体积(oil sac volume,OSV), 具体计算公式为

油囊的体积可用于表示油脂积累的程度。不同发育期的中华哲水蚤个体存在差异, 并会影响油囊的大小(Miller et al, 2000)。为消除个体大小差异产生的影响, 本研究采用油囊体积百分数(oil sac volume proportion, OSV%)来表征油脂积累情况, 即油囊体积占其前体部体积的百分比: OSV%=(OSV/PV)×100%(孙松等, 2011)。根据“50%”原则对各发育期的起始时间点进行估计, 即前一个发育期的 50%个体发育成该发育期的时间点(Uye, 1988)。此外, 根据 Niehoff等(2003)对哲水蚤性腺不同发育期的定义, 对中华哲水蚤的雌体性腺发育程度进行鉴定(四个性腺发育阶段: GS1、GS2、GS3和GS4, 其中GS4期为成熟期),并计算繁殖指标(reproductive index, RI)来表示整体性腺的发育水平, 即GS4期占雌性的百分比(Wang et al, 2009)。
1.6 统计方法
本文统计分析均使用SPSS(16.0)完成。对中华哲水蚤的生长发育、油脂积累以及性腺发育进行温度、饵料种类双因素方差分析。对野外各发育期中华哲水蚤体长与OSV%进行单因素方差分析。若方差分析结果显著, 选用S-N-K方法(Student-Newman-Keuls test,亦称 q检验)进行多重比较来确定不同实验处理间的差异。相关性分析采用Pearson相关系数。
2 结果
2.1 野外中华哲水蚤体长与油脂积累情况
4月胶州湾中华哲水蚤各发育期特征如图1所示,C5期幼体油脂含量最高(P<0.01), OSV%达到(1.03±1.35)%, 而其它发育阶段的OSV%接近0。雌体性腺发育成熟, 均为GS4期(RI=100%)。本实验未采到雄体。

图1 4月胶州湾中华哲水蚤各发育期的形态特征(a, b, c代表显著性差异, P<0.05)Fig. 1 Morphological characters in different development stages of Calanus sinicus in Jiaozhou Bay in April
2.2 各发育阶段的天数
在 15—18天培养中, 各温度饵料组的中华哲水蚤从C3期幼体逐步蜕皮发育为成体。各发育阶段的天数如图2所示。将50%个体蜕皮发育为成体前定义为幼体发育期, 各实验组中华哲水蚤在 8.2—13.6天后进入成体阶段, 10°C组幼体发育期较长(13.6天), 其它温度组发育较快(8.2—11.4天)。双因素方差分析结果显示(表 2), 温度对幼体发育期长短影响显著(P<0.05),饵料种类对发育时间的影响未达显著水平(P>0.05)。
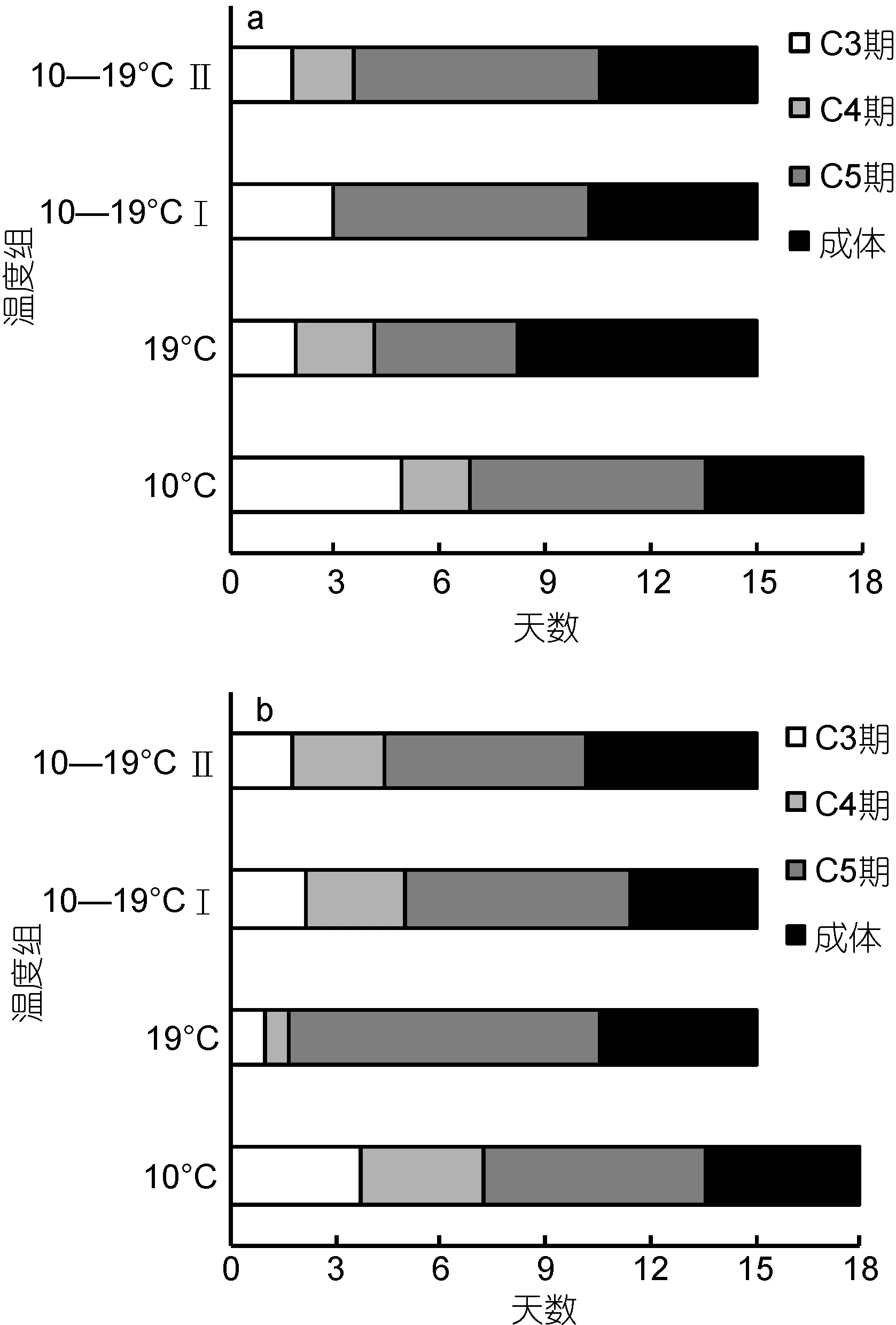
图2 各温度饵料组中华哲水蚤不同发育期的天数Fig.2 The number of days of development stage for Calanus sinicus in every temperature-food combination

表2 温度、饵料种类对中华哲水蚤生长发育与油脂积累影响的二因素方差分析结果汇总Tab. 2 Two-way ANOVA results for the impacts of temperature and food treatment on development and lipid accumulation of Calanus sinicus
2.3 油脂积累与C5期形态特征
中华哲水蚤的 OSV%随时间先增加后减小(图 3),并在6—12日达到了最高值。与此同时, C5期幼体所占的比例(C5%)也相应达到高值, OSV%与C5%成正相关(R=0.405,P<0.01)。对OSV%达到最高值时的各实验组 C5期幼体体长、油囊体积比进行温度、饵料浓度二因素方差分析(图 4; 表 2)。结果显示, 饵料种类对C5期幼体体长影响显著(P<0.01), 且硅藻培养下的C5期幼体体长较长。室内培养的 C5期幼体体长小于野外个体(P<0.01)。硅藻培养下, 各温度组C5期幼体体长相似; 而自然饵料培养下, 19°C组C5期幼体体长显著小于其它温度组(P<0.01)。另一方面, 温度和饵料种类对C5期幼体油脂积累均影响显著(P<0.01), 低温和硅藻饵料有利于油脂的积累。10°C硅藻培养下的 C5期幼体OSV%最大, 为(3.32±1.88)%, 最大值为7.58%。在硅藻培养下, C5期幼体OSV%在10°C组最高, 变温组次之, 19°C最低(P<0.01)。在自然饵料培养下, OSV%在 19°C组显著低于其它实验组(P<0.05)。各变温组间OSV%无显著差异, 均值范围为0.64%—1.49%。
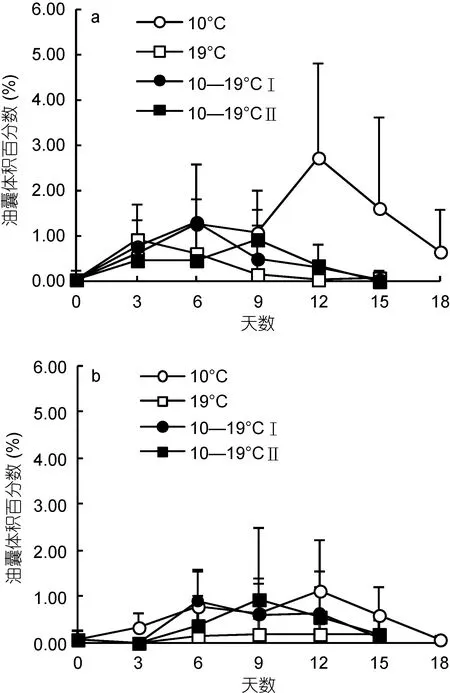
图3 各温度饵料组中华哲水蚤油囊体积百分数的变化Fig. 3 Variations in OSV% of Calanus sinicus in every temperature-food combination
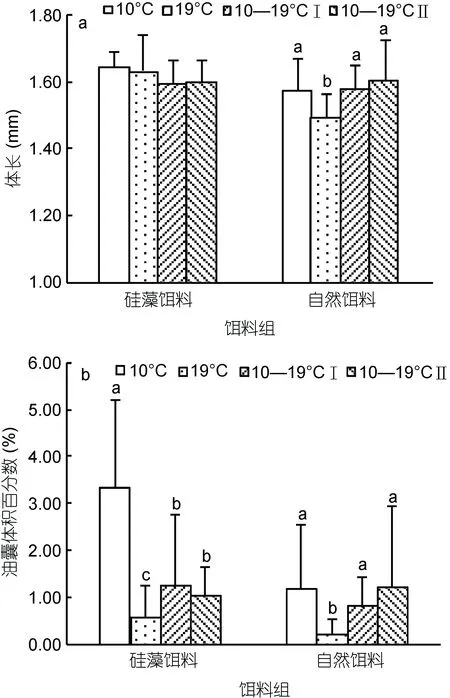
图4 各温度饵料组的油囊体积百分数达到最大时C5期幼体的形态特征(a, b, c代表显著性差异, P<0.05)Fig. 4 Morphological characters in the C5s stage in every temperature-food combination when maximum OSV% was reached
2.4 雌体形态特征与性腺发育
在两种饵料条件下的变温组中各有一只发育成雄体, 其余个体均发育为雌体。对雌体的形态特征进行温度、饵料种类双因素方差分析(表 2)。结果表明饵料种类对雌体的体长与 OSV%影响均不显著(P>0.05), 而温度对两者的影响均显著(P<0.01)。在硅藻培养下, 10°C 组的雌体体长为(1.99±0.06)mm, 显著高于其它温度组(P<0.01), 而在自然饵料培养下,19°C 组的雌体体长为(1.78±0.09)mm, 显著低于其它温度组(P<0.01)(图5)。室内培养的雌体的体长小于野外种群, 后者体长为(2.28±0.13)mm(P<0.01)。OSV%的对比结果与体长类似(图 5)。在硅藻培养下, 10°C组的 OSV%显著高于其它温度组(P<0.01); 而在自然饵料培养下, 19°C组的 OSV%显著低于其它实验组(P<0.01)。除10°C、硅藻培养下的雌体OSV%显著高于野外种群外(P<0.01), 其余个体与野外种群无显著差异。
进入成体阶段后, 中华哲水蚤的 OSV%逐渐减小(图3), 而性腺逐步发育至成熟(图6)。与硅藻饵料相比, 自然饵料培养下中华哲水蚤性腺发育速度较快、性腺成熟度更高(P<0.05)。实验结束时, 自然饵料培养下的 19°C 和变温组的雌性成熟度最高(RI:58%—65%)。而温度对性腺发育无显著影响(P>0.05)。
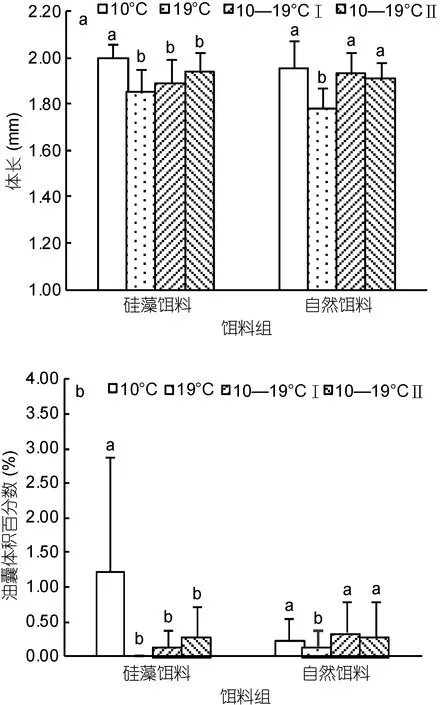
图5 各温度饵料组中华哲水蚤雌体的形态特征(a, b, c代表显著性差异, P<0.05)Fig. 5 Morphological characters of newly-molted females in every temperature-food combination
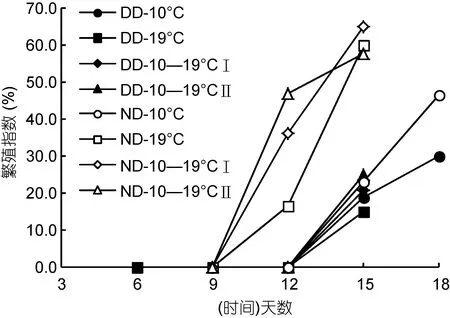
图6 各温度饵料组雌体繁殖指数变化Fig. 6 Variation of the reproduction index of Calanus sinicus females in each combination of temperature and food treatment
3 讨论
3.1 温度、饵料种类对中华哲水蚤油脂积累的影响
在幼体发育期(C3—C5期), 中华哲水蚤油脂不断积累, 油囊体积在C5期幼体达到最大。除19°C组OSV%低于同期野外种群, 其余个体与之相似或更大,实验所用饵料能够满足中华哲水蚤生长发育的需求。分析结果表明, 饵料种类、温度均对 C5期幼体油脂积累影响显著。恒温培养条件下硅藻饵料有利于油脂的积累, OSV%为自然饵料组的2.8倍。与其它休眠的哲水蚤一样, 中华哲水蚤油囊中主要成分为蜡脂(王延清等, 2014)。哲水蚤的蜡脂中主要的脂肪醇20:1(n-9)和 22:1(n-11)是由桡足类利用食物中的脂类和非脂类物质合成而来(Graeve et al, 2005)。相对于甲藻, 飞马哲水蚤摄食硅藻能积累更多的油脂(Pepin et al, 2011)。本研究中自然饵料的主要成分为硅藻, 优势种是角毛藻和骨条藻, 而硅藻饵料为中肋骨条藻和海链藻, 可见不同硅藻种类对 C5期幼体油脂积累贡献不同。自然饵料组成复杂, 微型浮游动物含量低,动物性饵料对中华哲水蚤油脂积累的影响需要对单种饵料进行研究。在变温条件下, 两种饵料对OSV%影响无显著差异, C5期幼体在变温条件下的油脂积累过程更为复杂。
低温有助于 C5期幼体油脂的积累, 个体的OSV%随着温度降低而增加。以往研究表明, 中华哲水蚤的呼吸率在适温下会随温度的降低而减少(张武昌等, 2000)。根据个体体长、培养温度进行估算(Ikeda et al, 2001; Li et al, 2004), C5期幼体在19°C时呼吸所需碳量为 3.04μgC/ind.·d, 约为 10°C(1.7μgC/ind.·d)的1.8倍, 可见低温有利于个体降低呼吸消耗以积累更多的油脂。此外, 低温下中华哲水蚤幼体发育期延长,为油脂积累提供了更多的时间。本研究中, 各变温组的C5期幼体的OSV%相似, 昼夜温度变化速率不会影响个体的油脂积累。变温组的C5期幼体的OSV%为10°C组的31%—102%, 为19°C组的 1.8—6.1倍,可见低温有利于油脂的积累。
本研究所用的饵料浓度较高, 其叶绿素a浓度相当于黄海春季较高水平。室内培养的 C5期幼体OSV%虽与黄海冷水团区四月份的种群相似, 却远低于五月份的种群(OSV%平均为30%)(孙松等, 2011)。室内变温条件虽有利于 C5期幼体油脂的积累, 却难以替代春末黄海冷水团复杂的水文环境。春末, 随黄海冷水团初见雏形, 温跃层底部出现叶绿素最大层(韦钦胜等, 2013), 加上此时黄海冷水团区浮游植物优势种为硅藻(宋书群, 2010), 饵料环境有利于油脂的积累。中华哲水蚤具有昼夜垂直迁移习性, 夜间摄食率高于白天(李超伦等, 2003)。白天, 中华哲水蚤在低温、低饵料环境的冷水团中活动, 呼吸率降低, 摄食率极低(Li et al, 2004); 夜间能迁移到温跃层附近,在饵料丰富的温暖水层摄食。研究表明, 与持续摄食的桡足类相比, 经 12—24小时饥饿培养的桡足类能在几小时内提高摄食率数倍, 大大增加了净能量的获取(Enright, 1977)。由此推断, 春末冷水团的水温和饵料条件有利于中华哲水蚤C5期幼体油脂的积累。然而, 度夏前黄海冷水团内 C5期幼体大体积油囊的形成还无法只用垂直移动和温差来解释。除环境因素外, 油脂积累过程也受到内部因素调节, 如相关基因表达、激素体液调节等(Tarrant et al, 2008)。哲水蚤休眠前的油脂积累可能是对多种环境因素变化的响应(如温度、饵料等), 其生理过程以及与多种环境因素的相互作用尚不明确 (Johnson et al, 2008), 需进一步研究以探讨中华哲水蚤的度夏机制。
3.2 温度、饵料种类对雌体形态与性腺发育的影响
雌体的体长主要受到温度的影响, 低温下体长较长, 这与 Uye(1988)研究结果一致。两种饵料培养下的雌体体长相似, 可见在个体生长方面硅藻饵料与自然饵料的作用差异不大。然而室内培养的雌体体长均小于同期野外种群, 包括 10°C下自然饵料组的雌体。一般情况下, 水温是影响中华哲水蚤体长的主要因素(陈清潮, 1964)。然而采样期间胶州湾水温为11.5°C, 高于室内的低温组(10°C)。这可能与室内培养条件存在瓶子效应(bottle effects)有关。自然海区水体大, 饵料种类丰富并存在斑块分布和集群效应, 实验室内的饵料种类和浓度均无法与之相比(Hakanson,1984)。
雌体的油囊体积主要受温度影响, 随温度上升而减小。中华哲水蚤进入成体阶段后, 油囊体积逐渐缩小, 与此同时性腺快速发育。越冬结束后, 飞马哲水蚤能将储存的蜡脂转变为磷脂用于卵磷脂的合成与性腺的初步发育(Rey-Rassat et al, 2002; Lee et al,2006)。中华哲水蚤雌体油囊的减小应与性腺发育有关。然而, 储存的油脂仅能支持中华哲水蚤性腺的初步发育, 性腺成熟必须通过摄食获取能量(Wang et al,2009)。
硅藻和自然饵料均能使中华哲水蚤的性腺发育成熟。投喂自然饵料能使雌体性腺发育速度加快、性成熟度更高。自然饵料的种类丰富, 其中纤毛虫生物量为 2.42—6.84μgC/L, 在黄海春夏季纤毛虫生物量范围(0.02—5.5μgC/L)附近(赵楠, 2008)。虽然纤毛虫在饵料中的比例低(1.21%—3.42%), 中华哲水蚤却能够优先摄食纤毛虫等微型浮游动物(汤宏俊等, 2015)。与单一的硅藻饵料相比, 微型浮游动物能为桡足类提供更多必需的营养物质, 如二十碳五烯酸(eicosapentaenoic acid, EPA)、二十二碳六烯酸(docosahexaenoic acid, DHA)、氨基酸和固醇类等, 有利于桡足类性腺的发育(Stoecker et al, 1990)。
4 结论
中华哲水蚤C5期幼体的油脂积累受温度和饵料种类的影响。低温有利于个体降低代谢消耗以增加油脂的积累。变温组的低温条件也有类似作用, 其 C5期幼体比高温组能积累更多的油脂。硅藻有利于 C5期幼体的油脂积累。与硅藻饵料相比, 自然饵料有利于中华哲水蚤的性腺发育。
致谢 本课题组罗璇在藻类培养上给予指导, 王世伟和刘梦坛在中华哲水蚤分类、培养上给予指导,冯颂和汤宏俊在样品采集上给予帮助, 谨致谢忱。
王延清, 李超伦, 刘梦坛等, 2014. 黄海春季和秋季中华哲水蚤脂类含量及组成分析. 海洋学报, 36(2): 99—107
韦钦胜, 傅明珠, 李 艳等, 2013. 南黄海冷水团海域溶解氧和叶绿素最大现象值及营养盐累积的季节演变. 海洋学报, 35(4): 142—154
孙 松, 王世伟, 李超伦, 2011. 黄海中华哲水蚤C5发育期油脂积累初步研究. 海洋与湖沼, 42(2): 165—169
陈清潮, 1964. 中华哲水蚤的繁殖、性比率和个体大小的研究.海洋与湖沼, 6(3): 272—288
宋书群, 2010. 黄、东海浮游植物功能群研究. 青岛: 中国科学院研究生院(海洋研究所)博士学位论文, 21
张武昌, 王 荣, 王 克, 2000. 温度对中华哲水蚤代谢率的影响. 海洋科学, 24(2): 42—44
李超伦, 王荣, 孙 松, 2003. 南 黄海 鳀 产卵场中华哲水蚤的数量分布及其摄食研究. 水产学报, 27(S1): 55—63
汤宏俊, 孙 松, 2015. 长江口几种优势桡足类对微型浮游动物的摄食研究. 海洋与湖沼, 46(1): 148—156
赵 楠, 2008. 黄海纤毛虫的丰度和生物量. 青岛: 中国科学院研究生院(海洋研究所)博士学位论文, 5—6
蒲新明, 2003. 中华哲水蚤夏季在南黄海的生活策略. 青岛:中国科学院研究生院(海洋研究所)硕士学位论文, 42
Atkinson A, 1996. Subantarctic copepods in an oceanic, low chlorophyll environment: ciliate predation, food selectivity and impact on prey populations Mar Ecol Prog Ser, 130: 85—96
Enright J T, 1977. Diurnal vertical migration: Adaptive significance and timing. Part 1. Selective advantage: A metabolic model1. Limnol Oceanogr, 22(5): 856—872
Graeve M, Albers C, Kattner G, 2005. Assimilation and biosynthesis of lipids in Arctic Calanus species based on feeding experiments with a13C labelled diatom. J Exp Mar Biol Ecol, 317(1): 109—125
Hakanson J L, 1984. The long and short term feeding condition in field-caught Calanus pacificus, as determined from the lipid content. Limnol Oceanogr, 29(4): 794—804
Hu D X, Wang Q Y, 2004. Interannual variability of the southern Yellow Sea Cold Water Mass. Chin J Oceanol Limnol, 22(3):231—236
Huang C, Uye S, Onbé T, 1993. Ontogenetic diel vertical migration of the planktonic copepod Calanus sinicus in the Inland Sea of Japan. III. Early summer and overall seasonal pattern. Mar Biol, 117(2): 289—299
Huo Y Z, Wang S W, Sun S et al, 2008. Feeding and egg production of the planktonic copepod Calanus sinicus in spring and autumn in the Yellow Sea, China. J Plankton Res,30(6): 723—734
Ikeda T, Kanno Y, Ozaki K et al, 2001. Metabolic rates of epipelagic marine copepods as a function of body mass and temperature. Mar Biol, 139(3): 587—596
Ji R B, 2011. Calanus finmarchicus diapause initiation: new view from traditional life history-based model. Mar Ecol Prog Ser,440: 105—114
Johnson C L, Leising A W, Runge J A et al, 2008. Characteristics of Calanus finmarchicus dormancy patterns in the Northwest Atlantic. ICES J Mar Sci, 65(3): 339—350
Lee R F, Hagen W, Kattner G, 2006. Lipid storage in marine zooplankton. Mar Ecol Prog Ser, 307: 273—306
Li C, Sun S, Wang R et al, 2004. Feeding and respiration rates of a planktonic copepod (Calanus sinicus) oversummering in Yellow Sea Cold Bottom Waters. Mar Biol, 145(1): 149—157
Miller C B, Crain J A, Morgan C A, 2000. Oil storage variability in Calanus finmarchicus. ICES J Mar Sci, 57(6): 1786—1799
Niehoff B, Runge J A, 2003. A revised methodology for prediction of egg production Calanus finmarchicus from preserved samples. J Plankton Res, 25(12): 1581—1587
Pepin P, Parrish C C, Head E J H, 2011. Late autumn condition of Calanus finmarchicus in the northwestern Atlantic: evidence of size-dependent differential feeding. Mar Ecol Prog Ser,423: 155—166
Pu X M, Sun S, Yang B et al, 2004. Life history strategies of Calanus sinicus in the southern Yellow Sea in summer. J Plankton Res, 26(9): 1059—1068
Rey-Rassat C, Irigoien X, Harris R et al, 2002. Energetic cost of gonad development in Calanus finmarchicus and C.helgolandicus. Mar Ecol Prog Ser, 238: 301—306
Stoecker D K, Capuzzo J M, 1990. Predation on Protozoa: its importance to zooplankton. J Plankton Res, 12(5): 891—908
Sun S, Huo Y Z, Yang B, 2010. Zooplankton functional groups on the continental shelf of the yellow sea. Deep Sea Res,Part II, 57(11—12): 1006—1016
Svetlichny L S, Kideys A E, Hubareva E S et al, 2006.Development and lipid storage in Calanus euxinus from the Black and Marmara seas: Variabilities due to habitat conditions. J Mar Syst, 59 (1—2): 52—62
Tarrant A M, Baumgartner M F, Verslycke T et al, 2008.Differential gene expression in diapausing and active Calanus finmarchicus (Copepoda). Mar Ecol Prog Ser, 355:193—207
Uye S, 1988. Temperature-dependent development and growth of Calanus sinicus (Copepoda: Calanoida) in the laboratory.Hydrobiologia, 167—168(1): 285—293
Wang S W, Li C L, Sun S et al, 2009. Spring and autumn reproduction of Calanus sinicus in the Yellow Sea. Mar Ecol Prog Ser, 379: 123—133
Zhou K L, Sun S, Wang M X et al, 2016. Differences in the physiological processes of Calanus sinicus inside and outside the Yellow Sea Cold Water Mass. J Plankton Res,38(3): 551—653
