3个芍药品种光合特性比较
2015-12-25刘小勇
孔 芬,刘小勇,陶 俊
(1.甘肃省农业科学院林果花卉研究所,甘肃 兰州 730070;2.扬州大学园艺与植物保护学院,江苏扬州 225009)
芍药(Paeonia lactiflora)是我国传统名花,栽培历史悠久,在我国华北、华东、华中以及甘肃兰州等地区均有种植[1-2],其作为观赏使用可追溯到4 000多年以前[3]。有关观赏芍药的研究多集中在品种、栽培管理以及切花生产等方面[4-8]。近年来,学者对芍药的光合特性、叶绿素荧光特性、碳水化合物代谢等方面展开了相关研究[9-13],但对观赏芍药不同品种间光合作用生理生态特性未做深入阐述。我们以常见栽培的3个芍药品种为材料,通过测定大田条件下不同芍药品种叶片光合速率日变化和响应曲线,分析光合速率与生态因子的相关性,探寻影响芍药生长发育的主要环境因子,为芍药高效、精确栽培及优良品种的引进筛选提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为3 a生芍药品种大富贵、黄金轮、杨妃出浴。
1.2 试验地概况
试验地选在扬州大学芍药园基地,位于东经119°42′,北纬32°38′,属于亚热带湿润气候区,四季分明,光照充足,气候温和,雨量充沛,自然条件优越。试验园年平均气温为14.8℃,年均日照时数为2 140 h,年均降水量1 020 mm,无霜期220 d。
1.3 试验方法
采用LI-6400型便携式光合作用系统(LI-COR,Lincoln,USA),6月上旬选择晴朗无风天气进行,光合日变化测定时间为6:00~18:00时,每整点测量1次,为消除时间上的误差,每次重复测定时,各品种间采取随机测定,每次各参数取4~6个有效数值。仪器同时记录各品种气孔导度(Gs)、蒸腾速率(Tr)、胞间 CO2浓度(Ci)等光合生理因子变化及光合有效辐射(PAR)、气温(Ta)、叶温(Tl)、相对湿度(RH)等相关光合生态因子的变化。水分利用效率(WUE)采用Fischer and Turner的方法 WUE=Pn/Tr进行计算[14]。
光响应曲线利用LI-6400自动光曲线程序来测定,选择晴天9:00~11:00时期间,光源为LI-6400配置的红蓝光LED光源,控制LI-6400参比室的CO2浓度为400μmol/mol,控制叶片温度为(25±1)℃。分别在光合有效辐射(PAR)依次为2 000,1 800,1 600,1 400,1 200,1 000,800,600,400,200,100,50,20,0μmo1/(m2·s)下测定3个芍药品种叶片的净光合速率(Pn)、Tr、Gs、Ci等光合参数。设定PAR为1 000μmol CO2/(m2·s)作为测定光强,利用LI-6400标配液化CO2钢瓶提供不同的CO2体积分数,分别在CO2浓度为 400、300、200、150、100、50、20、400、400、600、800、1 000、1 200μmol/mol的条件下测定叶片的Pn值。测定前使用1 200μmo1/(m2·s)冷光源诱导10 min,在每个CO2浓度下平衡120 s后测定Pn,当测量变异率小于0.05时由红外气体分析仪自动记录。测定时每品种选3~5个典型植株,每植株选取南部枝条自顶端向下数第4枚复叶的中间小叶为测定部位,重复3次,结果取平均值。
采用SPSS17.0及Excel软件进行数据处理分析和作图。
2 结果与分析
2.1 3个芍药品种光合日变化的规律
从图1可以看出,3个芍药品种的净光合速率变化的趋势一致,均呈双峰曲线,3个芍药品种叶片 Pn值在6:00~8:00时增加最为迅速,8:00时出现第1个峰值,分别是16.43、18.25、17.82μmo1/(m2·s),随后呈下降趋势,在13:00时大富贵和杨妃出浴出现低谷,黄金轮在14:00时谷值为7.78 μmo1/(m2·s),也就是出现了光合“午休”现象,经过午休之后,Pn值又开始上升,到16:00时大富贵和黄金轮达到第2个峰值,分别为9.49、8.88 μmo1/(m2·s),杨妃出浴则在15:00时达到第2个峰值为8.36μmo1/(m2·s),随后,净光合速率迅速下降,到18:00时达到全天最低值,分别是3.91、3.06、3.59μmo1/(m2·s)。
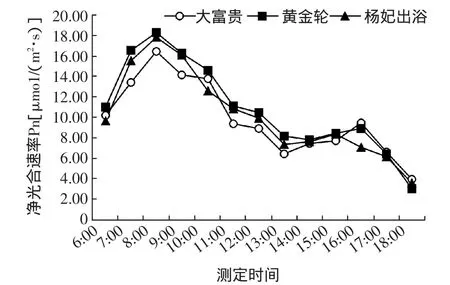
图1 3个芍药品种的净光合速率日变化
图2可以看出3个品种Gs日变化呈不明显的双峰曲线,植物通过改变气孔的开度控制与外界的CO2和水汽交换,从而调节光合速率和蒸腾速率[15],Gs第1个峰值出现在8:00时,黄金轮为0.31 mo1/(m2·s),明显高于大富贵和杨妃出浴,到16:00时均达到第2个峰值,均为0.07 mo1/(m2·s)。3个品种Ci日变化呈现下降的趋势,杨妃出浴变化幅度比较大,在9:00~14:00时期间3个品种保持在105~216μmo1/(m2·s);光照可引起气孔开放,提高大气和植物体温度,增加叶内外蒸汽压差加速蒸腾。图2可看出,3个品种的Tr均在中午之前达到一天中的最大值,其中大富贵在 10:00时,为 4.37 mmol/mol,黄金轮在 11:00时,为 4.76 mmol/mol,随后,迅速下降,16:00时小幅升高后迅速下降。3个品种的WUE总体上呈“倒抛物线”形状,从6:00时开始,大富贵和黄金轮2个品种的WUE从较高的数值随着蒸腾速率的升高而持续下降,杨妃出浴则在7:00时略有升高,中午前后3个品种的WUE维持在很低的水平,随后又随着蒸腾速率的下降而迅速升高。
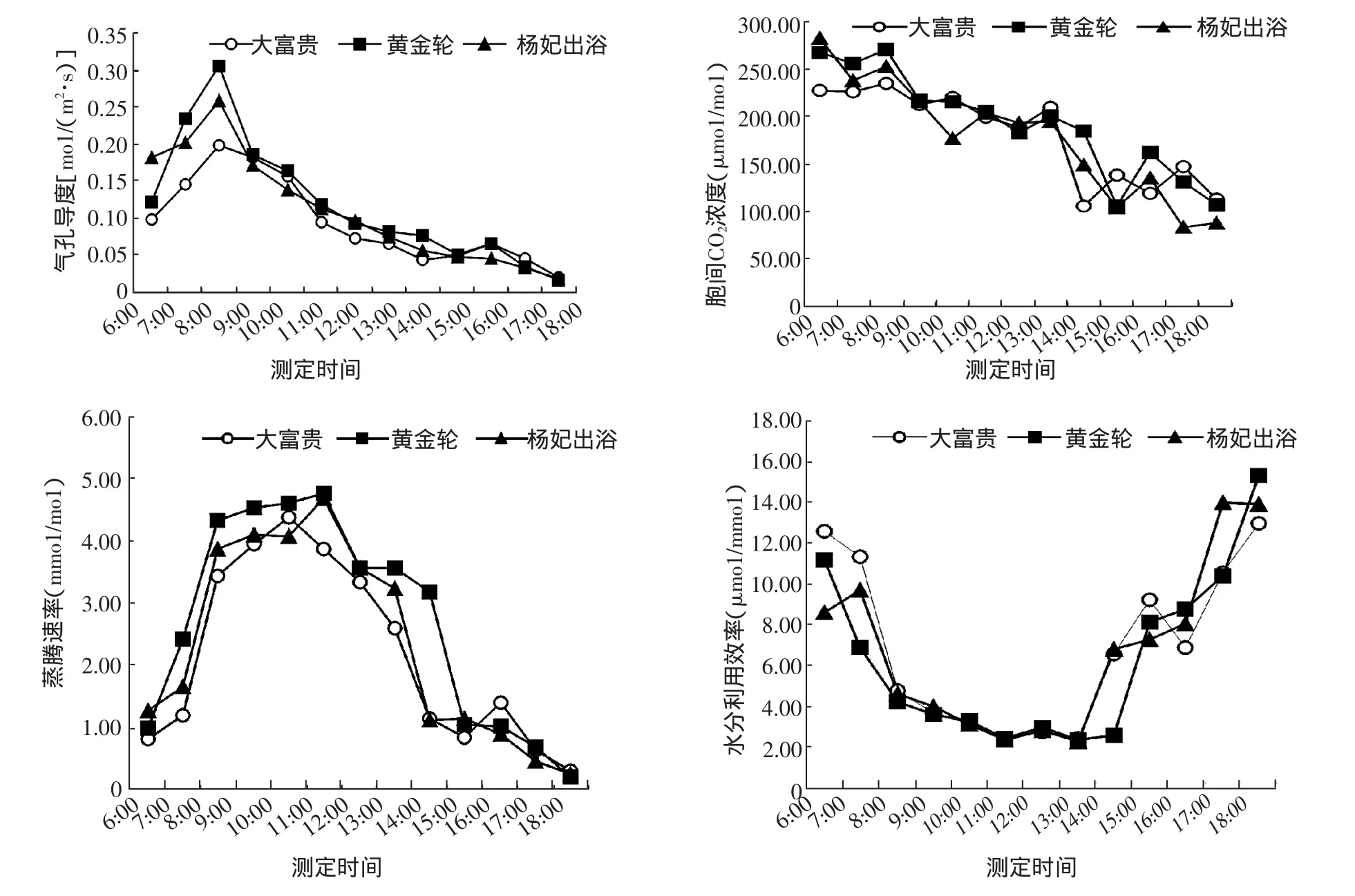
图2 3个芍药品种光合生理因子日变化
2.2 光合速率与环境因子的相关性分析
从表1可以看出,Pn与PAR和Tr呈显著正相正相关(r=0.752**,r=0.642**),与 Ta、T1呈负相关;当PAR增强时,大气温度(Ta)和叶片温度(Tl)随着升高,导致叶肉细胞间隙的水汽压与叶表面大气压差增大,蒸腾速率(Tr)随之提高;Tr与RH呈显著负相关(r=-0.733**),与Ta、Tl呈正相关,这是由于Ta、Tl与RH呈显著负相关(r=-0.958**,r=-0.626**),过高的RH将导致Ta和Tl的降低,进而降低了蒸腾速率。PAR和Tr呈极显著相关(r=0.899**),Tl的变化受Ta的直接影响(r=0.991**),同时Ta与RH呈极显著正相关(r=-0.958**),表明芍药的净光合速率可随光照和大气相对湿度的增大、温度的下降而提高。影响芍药光合的主要因子为Tr和PAR,RH通过别的因子间接影响光合速率。
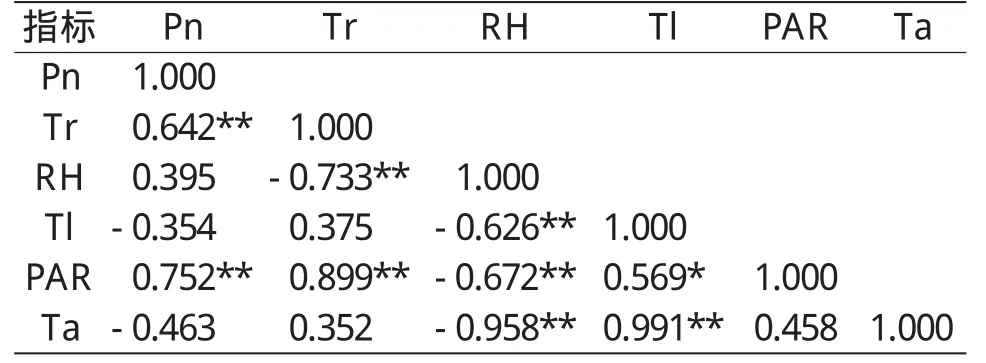
表1 光合速率与环境因子的相关性分析①
2.3 3个芍药品种光合-光响应特征
如图3所示,3个芍药品种的PAR在0~500 μmo1/(m2·s)时,Pn随着PAR的增加迅速增大,之后升幅较为平缓。PAR在500μmo1/(m2·s)之后,大富贵与杨妃出浴的Pn值较为接近,但明显低于黄金轮,差值约2μmo1/(m2·s);表明黄金轮较另外2个品种的光合能力强,其光能利用率最高。
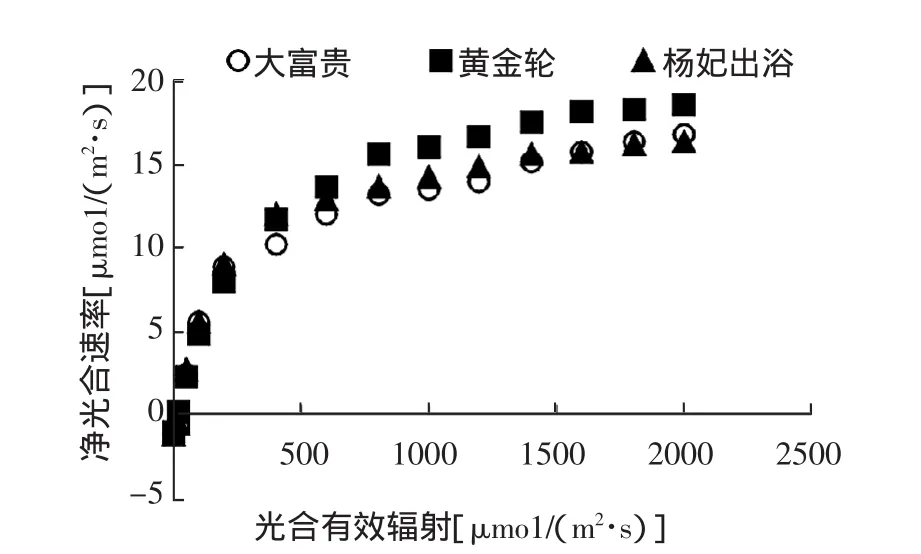
图3 3个芍药品种的光响应曲线
从图4可以看出,随着PAR的增大,Gs、Tr迅速升高,在PAR为1 000~2 000μmo1/(m2·s)左右时,黄金轮呈迅速上升趋势,之后的上升趋势则较为缓慢,3个品种的Gs自从大到小依次为黄金轮、大富贵、杨妃出浴。3个品种Ci的变化趋势均是随PAR的增大而迅速减小,而后逐渐趋于平衡,当PAR大于1 200μmo1/(m2·s)时,Ci又略有上升,随着PAR的增大,黄金轮的Ci值始终较大富贵与杨妃出浴大,而大富贵与杨妃出浴之间差异则不明显;水分利用效率决定于植物光合与蒸腾速率作用的比例。在PAR为0~400μmo1/(m2·s)时,3个芍药品种的WUE呈直线迅速上升趋势,大富贵与黄金轮在PAR为800μmo1/(m2·s)左右时达到最大值,分别是3.87、3.59 μmol/mmol,而杨妃出浴则是在PAR为600μmo1/(m2·s)左右时达到最大值,为 4.18μmol/mmol,3个品种的WUE值从大到小依次为杨妃出浴、大富贵、黄金轮。

图5 3个芍药品种的光合-CO2响应曲线

图4 3个芍药品种Gs、Ci、Tr、WUE对光强的响应
2.4 3个芍药品种光合作用的CO2响应曲线特征
从图5可以看出,3个品种的Pn值变化趋势一致,随着CO2浓度的增加,Pn值呈逐渐增大的趋势,当CO2浓度在0~400μmol/mol变幅内,Pn值增幅比较大,两者呈直线相关,相关系数都在0.98以上;当CO2浓度大于400μmol/mol之后,Pn值增加幅度逐渐减少,CO2浓度在800~1 200 μmol/mol时,Pn值逐渐趋于稳定,此时的净光合速率达到最大,即最大净光合速率(Pmax),这时的光合速率反映了光合能力的强弱,此时环境中的CO2浓度则为该植物的CO2饱和点,3个芍药品种的CO2饱和点介于800~1 200μmol/mol。杨妃出浴较之其它2个品种,Pn值一直较低。3个品种均在CO2浓度1 000μmol/mol时达到最大值,黄金轮和大富贵的Pn最大值分别为26.1、26.2μmo1/(m2·s),杨妃出浴较低,为23.2μmo1/(m2·s)。
3 小结与讨论
1)光合作用日变化是植物生产过程中物质积累与生理代谢的基本过程,也是分析环境因素影响植物生长和代谢的重要手段,不同种类、不同品种之间的光合特性都存在明显差异,如国兰、樱桃、牡丹品种等[16-18]。本试验3个芍药品种净光合速率日变化均呈双峰型曲线,第1个峰值都出现在8:00时,第2个峰值出现的时间不相同,这一结果与前人的研究结果存在一定差异。刘海琴和张玉对芍药的研究表明,光合速率日变化均呈单峰曲线, 峰值分别出现在 9:00时和11:00时[11,19],这可能是由植物本身的生理特性、生长环境及测定的时间不同而导致的。对同属植物牡丹的研究表明,夏季晴天大田栽培牡丹净光合速率日变化多呈明显的双峰曲线,最高峰华北和东北地区出现在9:00~11:00时,长江中下游地区在8:00~9:00时,次高峰东北地区则出现在16:00时左右[20-21]。此外3个品种的Ci、Gs、Tr、以及WUE方面均存在差别,在上午,Ci、Gs、Tr 3个指标的数值均为黄金轮最高,大富贵和杨妃出浴数值比较接近。而WUE却相反,随着蒸腾速率的升高而下降,3个品种间黄金轮最低,说明黄金轮较另外两个品种的光合能力更强,在强光下具有较高的光合速率,杨妃出浴则对弱光的利用能力要高于黄金轮和大富贵。
植物叶片Pn下降的自身因素主要是气孔的部分关闭以及叶肉细胞光合活性的下降。Ci的大小是评判气孔限制和非气孔限制的依据之一。判断叶片Pn下降究竟是气孔因素还是非气孔因素的方法是:若Gs降低的同时,Ci降低,则为气孔限制;若Ci升高,则为非气孔限制[15]。本试验3个品种,杨妃出浴的Pn、Gs和Ci同时下降,可认为Pn的下降为气孔限制,而大富贵和黄金轮在上午为气孔限制而下午Gs降低的同时Ci升高,构成非气孔限制,这表明芍药叶片光合(午休)不仅受来自气孔因素的限制也受非气孔因素的限制。有关气孔限制与非气孔限制的具体原因有待进一步研究。
2)植物的光合作用是一个对生态因子敏感的复杂生理过程,生态因子不仅直接影响光合作用,而且还通过影响植物的生理因子进而影响光合作用,各种因子间有着错综复杂的关系[22]。针对不同的植物,学者们得到结论也不一致。林金科认为茶园生态系统中光温因子是第1主成分,空气相对湿度和环境CO2浓度是第2主成分,阴天,叶温、环境CO2浓度是第1成分,光、空气相对湿度是第2主成分[23];对苜蓿的研究中表明,Pn与PAR与气孔导度呈极显著正相关,而与田间CO2浓度呈极显著负相关[24]。本研究表明,Pn与PAR和Tr呈显著正相关,与Ta、T1呈负相关,这与前人研究结果相一致[12,25]。影响光合的主要因子在不同的环境和条件下也是变化的,关于生理生态因子对不同芍药品种光合过程影响的机制有待于进一步研究。
3)植物的光响应曲线反映出植物光合生理特性的差异及其对不同生长光强的适应潜力[26]。本试验3个芍药品种的光响应曲线中黄金轮的Pn值一直高于大富贵和杨妃出浴,CO2的响应曲线中杨妃出浴Pn值最低,这说明黄金轮在强光、低浓度CO2条件下,如夏季的上午及午后,具有较高的净光合速率,而杨妃出浴则对光的反应能力最快,即对光更敏感,具有较好的利用弱光的能力。从光合速率对光及CO2响应曲线来看,CO2对Pn值的效应要高于PAR,这说明强光下(接近或超过光饱和点)芍药Pn值的增加主要是受CO2供应的限制,即CO2加富可以显著提高芍药叶片的净光合速率。
[1] 李嘉珏.中国牡丹与芍药[M].北京:中国林业出版社:1999:13-14.
[2] 秦魁杰.芍药[M].北京:中国林业出版社,2004.
[3] 王彦卓,姜卫兵,魏家星,等.芍药的文化意蕴及其园林应用[J]. 广东农业科学,2013(20):58-61.
[4] 郭先锋,臧德奎,袁 涛,等.我国栽培芍药溯源[J]. 山东农业大学学报,2008,39(3):388-392.
[5] 刘玉梅.观赏芍药生态习性及栽培技术研究进展[J].安徽农业科学,2008,36(12):4 965-4 967.
[6] 黄国京,贾清华,李 旻,等.我国观赏芍药栽培基质及施肥技术研究进展[J].黑龙江农业科学,2012,(1):148-150.
[7] 王历慧,郑黎文,于晓南.观赏芍药促成栽培技术与休眠解除的研究进展[J].北方园艺,2011(6):201-204.
[8] 袁燕波,王历慧,于晓南.低温和外源GA3对芍药切花品种花期调控的影响[J].东北林业大学学报,2014(1): 98-103.
[9] 简在友,王文全,孟 丽,等.芍药组内不同类群间光合特性及叶绿素荧光特性比较[J].植物生态学报,2010,34(12):1 463-1 471.
[10] 王秋玲,魏胜利,王文全.野生和栽培芍药植株形态特征与光合生理特性的比较研究[J].中国中药杂志,2012,37(1):32-36.
[11] 刘海琴,王康才,罗庆云,等.叶面喷施铜肥对芍药光合及荧光动力学参数的影响[J].江苏农业学报,2010,26(5): 982-986.
[12] 周盛茂,段净亚,李 明,等.华北高寒区芍药光合特性的研究[J]. 江苏农业科学,2013,41(3):148-150.
[13] 宋焕芝.芍药光合特性及碳水化合物代谢研究[D].北京:北京林业大学,2012.
[14] PEÑUELASJ,FILELLA I,LLUSIÀJ,et al.Compara-tive field study of spring and summer leafgas exchange and photobiology of the Mediterranean trees Quercusilex and Philly realatifolia[J]. Journal of Experimental Botany,1998,49:229-238
[15] 许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[16] 李鹏民,高辉远,邹 琦,等.5种国兰(Cymbium)的光合特性[J]. 园艺学报,2005,32(1):151-154.
[17] 杨江山,常永义,种培芳.3个樱桃品种光合特性比较研究[J]. 园艺学报,2005,32(5):773-777.
[18] 侯小改.4个牡丹品种光合特性的比较研究[J].河南农业大学学报,2007,41(5):527-530.
[19] 张 玉,李 霞,郭绍霞.芍药光合特性研究[J].青岛农业大学学报(自然科学版),2011,28(1):24-27.
[20] 侯小改,马慧丽,段春燕,等.牡丹“佛门袈裟”光合特性的研究[J].沈阳农业大学学报,2006,37(5):772-774.
[21] 岳 桦,王欠欠.六个紫斑牡丹品种光合特性初步研究[J]. 黑龙江农业科学,2011(1):39-41.
[22] 付瑞锋,梁宗锁,王 琬.大叶秦艽的光合特研究[J]. 西北农业学报,2014,23(11):185-190.
[23] 林金科,赖明志.影响茶树叶片净光合速率的生态生理因子的初步分析[J]. 作物学报,2000,26(1),110-115.
[24] 万素梅,贾志宽,杨宝平.苜蓿光合速率日变化及其与环境因子的关系[J]. 草地学报,2009,17(1):27-31.
[25] 杜庆平,孙 燕,李成忠,等.扬州地区2个芍药品种光合生理生态特性比较[J].安徽农业大学学报,2011,38(6): 935-939.
[26] 韦兰英,曾丹娟,张建亮,等.岩溶石漠化区四种牧草植物光合生理适应性特征[J].草业学报2010,19(3): 212-219.
