黄曲霉漆酶基因HIGS载体的构建及对花生的转化
2015-12-23孙全喜王云云王秀贞唐月异吴琪张青云曹广英王传堂
孙全喜 王云云 王秀贞 唐月异 吴琪 张青云 曹广英 王传堂


摘要:黄曲霉毒素污染严重影响着花生食品安全。通过常规育种的方式培育抗黄曲霉花生新品种进展缓慢,效果也难如人意。HTGS(Host-Induoed Gene Silencing,寄主诱导的转基因沉默)技术是一种新兴的RNA干扰技术,为培育抗病植物提供了可能。但该技术在花生上的应用尚未见报道。为创造抗黄曲霉花生新品系,本研究利用该技术构建了两个黄曲霉漆酶基因(Lac1和Lac2)的HIGS载体,并试图将其转化到易感黄曲霉的花生品种粤油20中,获得了转Lac1基因的PCR阳性种子。根据HIGS的原理,黄曲霉侵染后,转基因花生中产生的dsRNA将抑制黄曲霉漆酶基因表达,从而使转基因花生对黄曲霉产生抗性。基于该原理,下一步将对转基因种子是否抗黄曲霉侵染进行鉴定。本研究为利用HTGS技术探索黄曲霉漆酶基因与黄曲霉致病性的关系奠定了基础,为培育抗黄曲霉花生品种提供了一条新思路。
关键词:花生;黄曲霉;漆酶基因;寄主诱导的转基因沉默(HIGS);遗传转化
中图分类号:S565.203.53
文献标识号:A
文章编号:1001-4942(2015)10-0008-05
花生是世界重要的油料作物和经济作物,特别在广大发展中国家,是重要的食用蛋白源和食用植物油源。我国是世界花生生产和消费大国,也是花生贸易大国。然而,花生易受黄曲霉菌侵染而产生黄曲霉毒素,严重影响着花生的食品安全。
黄曲霉侵染花生时,会诱导花生产生抗病反应,释放植保素(Phytoalexins)。花生不同品种中植保素的积累被认为与抗病性有关。花生中的植保素主要是芪类化合物(Stilbenoids),此类化合物大部分具有抗真菌活性。而黄曲霉漆酶(Laccase)能够降解芪类化合物,漆酶活性高低与黄曲霉致病性强弱有直接关系。
漆酶是一种含铜多酚氧化酶,广泛存在于植物、高等真菌和一些细菌分泌物中。在高等植物中,漆酶主要与木质素降解有关。另外一些研究认为漆酶还与病原真菌的致病性相关。单宁酸(Tannic acid)是一种植保素,Kim等认为,在板栗疫病菌(Cryphonectria parasitica)侵染过程中,漆酶的存在可以使致病菌对单宁酸的抗性提高,从而增强对该病原的易感性。此外,还有漆酶参与人类病原真菌致病性的研究报道。
RNA沉默(RNAi)是植物抵御病毒入侵的一种自然防御机制。植物RNAi信号不但可以在相邻细胞间传递,而且可以通过维管束进行远距离传播,并且能够跨越物种界限,由寄主传递给寄生的植物、线虫、细菌、真菌或取食植物的昆虫。通过在植物(寄主)中异源表达病原物或昆虫(寄生物)目标基因的dsRNA,可诱发RNAi特异沉默病原或昆虫中目标基因的表达。这种寄主诱导的转基因沉默(Host-In-duced Gene Silencing,HIGS)为作物抗病育种提供了新思路。
为研究HIGS技术在花生抗黄曲霉中的应用效果,培育出抗黄曲霉花生新品种,本研究以黄曲霉漆酶基因作为HIGS靶标,构建其RNAi载体,对感黄曲霉花生品种粤油20进行转化,获得了推断转化体。
1材料与方法
1.1试剂及材料
本研究所用花生材料为易感黄曲霉品种粤油20。大肠杆菌DH5α感受态、DNA分子量标记、PCR Mix等购自北京全式金生物有限公司;限制性内切酶均购自Fermentas公司;T4 DNA连接酶购自宝生物工程(大连)有限公司;质粒小提试剂盒和胶回收试剂盒购自天根生化科技(北京)有限公司;农杆菌菌株EHA105以及RNAi载体pF-GC5941为本实验室保存。
1.2DNA和氨基酸序列分析、DNA合成
DNA测序和DNA合成均由上海桑尼生物科技有限公司完成。基因开放阅读框(ORF)由ORF Finder(http://www.ncbi.nlm.nih.gov/projects/gorf/)获得。氨基酸序列比对由DNAMAN序列分析软件(LynnonBiosoft)完成。
1.3农杆菌介导的遗传转化
2014年7月在莱西试验站专门的转基因试验区按本项目组专利技术(专利号:ZL2011 10058531.2)对粤油20进行农杆菌注射。10月收获种子,晾干,保存。
1.4转基因花生的PCR检测
在花生种子远离胚芽的一端切下薄薄的一层子叶组织,利用该部分组织提取DNA,提取方法参照国家发明专利“花生健康组织和病组织简便快速DNA提取方法”(专利号:ZL 2009 10255786.0)。经上述处理的花生种子可以进行正常种植。用基因特异引物(见表1)进行PCR扩增,筛选PCR阳性花生种子。PCR程序如下:94℃预变性5min;94℃变性30s,58℃退火30s,72℃延伸30s,35个循环;72℃延伸10min。
2结果与分析
2.1黄曲霉漆酶基因片段的获得
在NCBI上搜索到两个可能的漆酶基因(XM_002373291和XM_002382249),长度分别为1764bp和1797bp,分别标记为Lac1和Lac2。经NCBI Blast分析,发现两者都具有3个保守的铜原子结合位点。将Lac1和Lac2编码的氨基酸与GenBank中登录的蛋白序列进行比对后发现:这两个基因与其它真菌漆酶基因编码的氨基酸序列具有较高的同源性(图1)。
分别选取Lac1(139-448)和Lac2(20-329)各310bp的序列(图2),每个片段的5′端添加限制性内切酶Xba I-Asc I识别序列,3′端添加Swa I-BamH I识别序列,以方便构建RNAi载体。由上海桑尼生物科技有限公司进行全基因合成。片段合成后,亚克隆到克隆载体pUC57中,得到的载体分别命名为pUC57-Lac1和pUC57-Lac2。经M13F引物测序,序列完全正确。endprint
2.2黄曲霉漆酶基因RNAi载体的构建
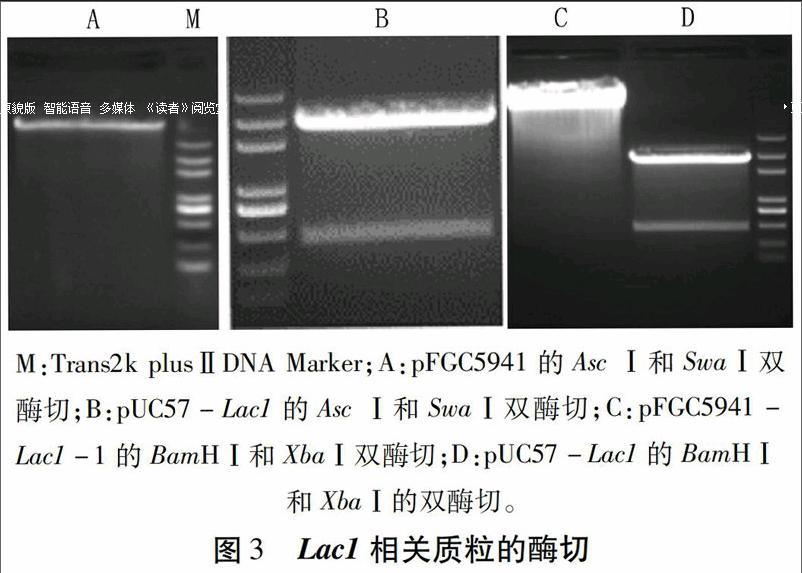
用限制性内切酶Asc I和Swa I将Lac1和Lac2片段分别从pUC57-Lac1(图3B,以Lac1为例,下同)和pUC57-Lac2切下,连接到同样经过Asc I和Swa I酶切的植物RNAi载体pFGC5941(图3A,载体信息见http://www.snapgene.com/resources/plasmid_files/plant_vectors/pFGC5941/)。经特异引物(见表1)筛选,得到含目的片段的质粒pF-GC5941-Lac1-1(图4A)和pFGC5941-Lac2-1。
质粒pUC57-Lac1和pUC57-Lac2经BamH I和Xba I酶切(图3D),得到的片段连接到经BamH I和Xba I酶切的pFGC5941-Lac1-1(图3C)和pFGC5941-Lac2-1中,经PCR筛选,得到带有第二个片段的质粒pFGC5941-Lac1-2(图4B)和pFGC5941-Lac2-2。此时,载体中均包含一个正向目的片段和反向目的片段及中间由ChsA intron间隔的结构。
2.3转基因花生T0代种子的PCR检测
将构建好的质粒pFGC5941-Lac1-2转化到农杆菌菌株EHA105中,利用本课题组发明的农杆菌转化法对易感黄曲霉品种粤油20进行遗传转化,收获了转Lac1基因的花生种子29粒;提取DNA后,利用Lac1特异引物(见表1)进行PCR筛选。结果筛选到转Lac1的PCR阳性花生种子11粒,转化率达38%。图5仅显示部分结果,其中2~5号和7号样品有目的条带,N为阴性对照,P为阳性对照。目前,这些PCR阳性花生种子已种于转基因隔离区,所结种子将用于黄曲霉抗性鉴定。
3讨论与结论
国内外长期以来通过常规手段培育抗黄曲霉花生品种,周期长、进展缓慢且效果不甚理想。基因工程技术有望解决花生黄曲霉毒素污染问题。寄主诱导的转基因沉默(HIGS)技术是一种研究病原物(真菌等)基因功能以及控制病原物造成其寄主病害的全新技术。该技术将构建的含病原物目的基因片段发夹结构的载体转化到寄主中,从而导致病原物特定基因序列双链RNA(dsRNA)表达;当病原物侵染表达了dsRNA的寄主时,dsRNA由寄主传递给病原物,对病原菌内源特定基因的表达进行RNA干扰,从而引起病原菌特定基因表达的下调。该技术为花生抗黄曲霉育种提供了新思路,但目前尚未见有利用HIGS技术增强花生对黄曲霉抗性的研究报道。
黄曲霉漆酶活性高低被认为与黄曲霉致病性强弱有直接关系。为了抑制黄曲霉漆酶基因Lac1和Lac2的表达,本研究以黄曲霉两个漆酶基因为HIGS的靶标,构建其RNAi载体,并对感黄曲霉花生品种粤油20进行农杆菌转化,初步获得了转Lac1的PCR阳性花生种子。从图5中可以看出,PCR阳性样品目的条带较弱,可能与提取的DNA质量较差有关。花生种子中油分含量较高,DNA较难提取,其提取质量与操作者的不熟练程度密切相关。
为进一步研究转黄曲霉漆酶基因的花生是否抗黄曲霉,本课题组已将PCR阳性转Lac1花生推断转化体种子种植于转基因试验区,待收获种子后将进行黄曲霉侵染鉴定,检测其是否获得了黄曲霉抗性。同时,将继续用含Lac2的RNAi载体对粤油20进行遗传转化,以期获得抑制Lac2表达的转基因花生。
本研究为利用HIGS技术探讨黄曲霉漆酶基因与黄曲霉致病性的关系奠定了基础,为培育抗黄曲霉花生新品种提供了一条新的思路。endprint
