板栗‘遵玉’大小孢子发生及雌雄配子体发育研究
2015-12-21苏淑钗
陈 昉,苏淑钗
(北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083)
板栗‘遵玉’大小孢子发生及雌雄配子体发育研究
陈 昉,苏淑钗
(北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083)
为了系统了解板栗雌雄花发育过程,以板栗优良品种‘遵玉’为试材,对大小孢子及雌雄配子体发生发育进行研究。结果表明:花药壁由外而内分别为表皮、药室内壁、中层及绒毡层(为腺质型),属于基本型。小孢子胞质分离为同时型,四分体为典型的四面体型,少数呈左右对称排列。雄配子体只经过1次有丝分裂,花粉成熟为2-细胞型。雌花胚囊发育为蓼型胚囊,只有1个胚囊母细胞发育成胚囊,四分体呈线性排列,整个发育过程与雌雄花外部特征密切相关。确定观察减数分裂最佳取样时期为:雄花花序长度为10~17 cm,总苞直径为0.9~1.6 cm。‘遵玉’小孢子及雄配子体发生发育不存在异常表现,表明‘遵玉’雄配子体发育正常,花粉可育,可以作为授粉树。
板栗;雄配子体;雌配子体
板栗Castanea mollissimaBlume在我国栽培历史悠久,营养丰富,经济价值较高[1-2],是我国重要的经济林树种,同时也是我国优良的坚果类果树[3-6]。
板栗‘遵玉’是“十五”期间审定的适于矮密栽培的优良板栗品种。此前有学者对板栗大小孢子发育进行了初步研究[7-11],但并没有关于‘遵玉’品种的相关报道。该试验中以‘遵玉’为材料,运用解剖方式对其大小孢子及雌雄配子体发生发育进程进行显微拍照记录,并且对外部形态特征进行对应性探讨,以期系统地了解板栗‘遵玉’雌雄花发育的形态学和组织学特性,为遵化地区板栗育种和生殖生物学研究提供资料,为生产者在板栗园栽培过程中进行适当的花期管理,创造良好的发育条件提供科学依据。
1 材料与方法
试材‘遵玉’来自河北省遵化市魏进河林场板栗品种对比园,树龄15 a,树势良好。2012年5~6月,每天上午9:00~10:00采集树冠外围大小均等的雌花以及雄花序各12个,每3天1次,用FAA固定液固定24 h,固定充分,并对雄花序外部形态特征进行记录。
切片制作采用常规石蜡制片法,将固定材料经不同梯度乙醇逐级脱水、浸蜡、包埋后切片,切片机型号为Leica RM2265,切片厚度10 μm,铁矾-苏木精染色,0.5%曙红溶液对染,树胶封片[12-13],使用奥林巴斯光学显微镜对切片进行观察、照相。
2 结果与分析
2.1 雄配子体发育
2.1.1 花药壁发育(雄花序增长—雄花成熟)
板栗品种‘遵玉’花药有4个药室,药室左右基本对称,在5月中旬处于造孢细胞期,雄蕊原基分裂并增长(见图1-A~D),上部发育成分生组织,通过尖端生长和边缘生长,分生组织形成嫩花药。随着花药原始体的出现,花药四角细胞发生变化,胞内细胞核增大,原生质稠密,发育为孢原细胞(见图1-E),共计4组。孢原细胞出现在原基表皮下,经平周分裂后由外而内发育成初生壁细胞和初生造孢细胞2层细胞(见图1-F)。次生壁细胞为2层同心圆细胞,由初生壁细胞经平周分裂发育而成,在花粉母细胞(小孢子母细胞)开始分裂前,次生壁细胞先行分裂,由外而内形成表皮、药室内壁、中层和绒毡层(见图1-G、H、I、L、O)。发育类型为基本型。表皮细胞较大,药室内壁细胞较小,中层细胞有1~2层,有的甚至没有中层细胞,可能随着小孢子母细胞的减数分裂,中层细胞逐层解体,绒毡层细胞在花粉母细胞减数分裂期间,细胞变大、细胞核增大、细胞质稠密,直至四分体时期,绒毡层开始消散(见图1-O、Q)。当形成单核花粉粒后,药室内壁细胞的细胞壁纤维状明显增厚(见图1-R、U、V)。二细胞花粉粒形成时,中层细胞几乎全部解体,随着花粉粒成熟,绒毡层细胞也完全解体,整个发育过程中始终维持在原来的位置,属于腺质型(见图1-U、V)。花药壁此时只有褶皱的表皮层和加厚的药室内壁。
2.1.2 减数分裂以及雄配子体发育(雄花序增长—雄花成熟)
5月25日前后,花粉母细胞经过初生造孢细胞多次有丝分裂形成,细胞质较浓,无明显液泡形状,从不规则多边形逐渐呈圆形(见图1-G、H)。随后花粉母细胞开始减数分裂,分裂时期持续时间短。经过减数分裂前期(见图1-I)、减数分裂I中期(见图1-J)、后期(见图1-K)、末期,减数分裂第1阶段结束,细胞内形成2个子核,胞质不分裂,不形成二分体(见图1-L);随后依次进入减数分裂Ⅱ中期(见图1-M)、后期(见图1-N)、末期(见图1-O),细胞内分裂为四核,进入四分体时期,在形成四分体的同时,新的细胞壁形成,将花粉母细胞分裂成4个细胞,即小孢子细胞,小孢子并未马上分离,而是包裹在同一个胼胝质细胞壁内,其中四面体型普遍存在,多数四分体属于此类型,少数为左右对称型(见图1-P、Q);进入小孢子时期,胼胝质开始发生消融,随后小孢子从四分体中慢慢游离出去,即为花粉粒,共计4枚且单核(见图1-R)。根据‘遵玉’板栗小孢子减数分裂情况,确定其胞质分裂类型为同时型。随着小孢子离体四分体,并充斥于小孢子囊中,标志着雄配子体时期的到来。小孢子刚离体四分体时,细胞整体偏小,具有壁薄质密的特点,细胞核位于胞体中心,随后细胞体变大,细胞质液泡化,小孢子发育进入单核时期(见图1-R);随后液泡逐渐增大,将细胞核挤到花药壁的一侧,花粉发育进入单核靠边期(见图1-S);此后,贴近花药壁位置的花粉细胞核进行1次不均等的分裂,形成1个体积较大的营养细胞和较小的生殖细胞,随后花粉粒的体积明显增大,生成淀粉、脂肪等营养物质,中央大液泡逐渐消失,花粉粒渐次成熟,呈现圆形或椭圆形,出现明显发芽孔,‘遵玉’板栗成熟的花粉为二细胞花粉(见图1-T);花粉粒发育临近成熟,紧邻的花粉囊的囊壁开始破碎,花粉囊彼此连通(见图1-U、V)。

图1 雄配子体发育的显微观察Fig. 1 Microscopic observation of male gametophyte development
2.2 雌配子体发育
2.2.1 雌花发育
早在冬季休眠结束后,混合花序基部逐渐出现突起,并且慢慢扩大,形成半圆形结构,这便是雌花原基,到5月初雌花原基发生变化,再次形成突起,形成花朵原基,随着花朵原基不断发育,其顶部逐渐变得平钝,其边缘慢慢出现突起,即花萼原基,花萼原基不断伸长并卷曲,将雌花的内部结构包被其中,同时内部形成雌蕊原基(见图2-A),至5月中旬,雌蕊原基慢慢伸长并变大,形成花柱原基(见图2-B),花柱原基形成后,不断伸长,最后突破花萼的包被,并形成花柱和柱头(见图2-C),而花柱顶部裂开的部分即为柱头,分化期一般在5月末至6月初,随着花柱的停止生长,位于花柱下方的细胞,不断裂解,于是出现中空的结构,这便是子房的雏形(见图2-D),此阶段最早出现在6月5日左右,随后缝隙不断地扩大,形成子房,并且有胚珠原基的出现(见图2-E),倒生于中轴胎座之上,随后进入大孢子发生及雌配子体形成阶段。
2.2.2 大孢子发生及雌配子体形成
板栗珠被为双珠被,由于外珠被较内珠被发育快,在外珠被和内珠被之间出现一定程度的空隙(见图2-F),随后珠心内部出现1颗孢原细胞,此细胞明显不同于周围细胞结构,具有体大、质密的特征,孢原细胞直接可以发育成为大孢子母细胞(见图2-G);胚囊母细胞(即大孢子母细胞)随后通过2次减数分裂,分别发育为大孢子的二分体(见图2-H)和四分体(见图2-I),但是一般只有1个大孢子能够发育为胚囊(即功能大孢子),其它3个大孢子逐渐退化并消失;接着功能大孢子逐渐发育为胚囊,起初为单核(见图2-J),随后通过3次有丝分裂逐渐发育为2核、4核直至8核胚囊,2核胚囊中央有大液泡(见图2-K),2核位于上下两端,2核变4核(见图2-L),直至胚囊上下各有4核,随后上下各有1核向中央移动,即2个极核,2枚助细胞、1枚卵细胞呈品字排列,位于珠孔端,而3枚反足细胞位于合点端,至此,胚囊完全发育为成熟胚囊。
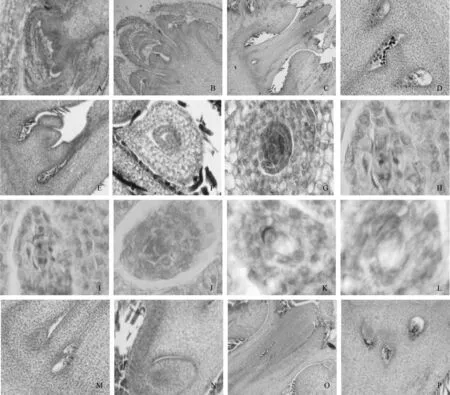
图2 雌配子体发育的显微观察Fig. 2 Microscopic observation of female gametophyte development
2.3 雌雄花发育的不同步性
通过观察发现同一时期的不同取材或是相同取材,大小孢子发生及雄配子体发育并非完全同步。例如,取自不同单株的雄花,1个处在成熟花粉粒时期,1个处在减数分裂I后期(见图1-W、X);同一雄花的不同花药,1个处在四分体时期,1个处在减数分裂I后期(见图1-Y);而不同单株的雌花,1个处于子房形成期,1个处于胚珠形成期(见图2-M、N);同一单株的不同雌花,1个处于花柱伸长期,1个处于子房形成期(见图2-O、P)。
2.4 雌雄花发育与外部特征的相关性
通过观察发现雌雄花的发育进程与其形态变化密切相关,雌雄花的不同形态能够代表某一发育时期,虽然没有完全吻合,但大部分是处于同一时期,此结果说明如下取样标准可行,同时说明大小孢子母细胞发育的不同步性。雄花序出现15 d左右,长度大约6 cm,雄花扁平,颜色呈嫩绿,为造孢细胞期;20 d左右,长度大约10 cm,雄花膨大,颜色呈绿色,为小孢子母细胞期;27 d左右,长度大约17 cm,并且慢慢停止伸长,雄花逐渐饱满,颜色呈绿色,为雄配子体形成期;32 d左右,少数出现花粉孔,为花粉粒成熟期。雌花在直径为0.2~0.3 cm时,颜色呈嫩黄色,为花萼原基及雌蕊原基形成时期;当总苞直径为0.7 cm时,花柱刚刚露出,为子房雏形出现期;当总苞为1.0 cm,刺苞变硬,花柱角度为90°时,为胚珠原基出现期;总苞为1.3 cm时,进入胚囊形成时期。
3 结论与讨论
(1)‘遵玉’板栗的花药为4室,花药壁由外向内分别为表皮、药室内壁、中层和绒毡层,发育属于基本型,与任中立等人报道不一致[7],推测其原因可能是中层细胞在小孢子母细胞减数分裂进程中分解,其分解原因还需进一步研究。绒毡层为腺质绒毡层,花药发育过程中不断提供营养,直至花粉成熟,绒毡层消失。之前有报道花粉母细胞分裂形成的四分体呈典型的四面体型排列[14],本研究中发现‘遵玉’板栗大多符合这种类型,但仍存在少数呈左右对称排列,小孢子母细胞胞质分裂为同时型,2-细胞花粉粒,具有明显萌发孔,这些特征与之前报道基本一致[8,14],也与壳斗科其它植物十分相似[15-16],说明板栗‘遵玉’小孢子及雄配子体发生发育不存在异常,雄配子体发育正常,花粉可育,可以作为授粉树。
(2)‘遵玉’板栗雌花发育依次形成花朵原基、花萼原基、雌蕊原基、花柱原基、子房、胚珠、胚囊,其中花萼原基与雌蕊原基同时出现,但花萼原基发育较快,将雌蕊原基包被其中,这与之前张林平等的报道有所不同[17],‘遵玉’板栗胚囊为蓼型胚囊,大孢子四分体呈直线排列,只有1个胚囊发育为成熟胚囊。
(3)经观察发现,无论是不同单株,同一单株的不同花序,同一花序上不同位置的雌雄花,还是同一雄花的不同花药,雌雄花整个发育过程并非完全一致,甚至在同一视野下,可以看到发育过程的不同时期,这种不同步性正好延长了板栗雌雄花花期的相交时间,增进了授粉的有效作用时间,此现象有利于种群繁殖后代,这是一种植物进化适应环境的表现,同时也体现出异花授粉植物比自花授粉植物同步性低的一般规律[18]。
(4)板栗花粉母细胞减数分裂时期历时较短,从花粉母细胞形成到减数分裂完成只有5~7 d[7],而从大孢子母细胞形成到成熟胚囊也仅5~6 d。如果没有在适合的时期取样,则无法观察到雌雄配子体的形成,或者错过雌雄花分化的关键期。经观察发现,板栗雌雄花发育与其外部形态有着密切关系,在雄花序出现15~20 d,花序长度为10~17 cm时,为最佳取样时期,而雌花在总苞直径为0.9~1.6 cm,同时也是雄花刚刚盛开后,为雌花的最佳取样时期。
[1]黄亚丽,王庆江,郭云龙.喷施5种叶面肥对板栗产量和品质的影响[J].经济林研究,2013,31(3):143-145.
[2]李广会,郭素娟,邹 锋.板栗叶片营养与土壤养分的动态变化及回归分析[J].中南林业科技大学学报,2012,32(9):41-46.
[3]白仲奎.板栗[M].北京:经济管理出版社,1997:1-2.
[4]唐时俊,李润唐,李昌珠,等,板栗丰产栽培技术[M].长沙:湖南科学技术出版社,1992.
[5]罗建谱.果树周年管理技术[M].长沙:湖南科学出版社, 1995.
[6]陈建华,何 钢,李志辉.促进板栗雌花芽分化的研究[J]. 中南林学院学报,2002, 22(1):27-30.
[7]任立中,杨其光,杜国华.板栗花性别和器官分化的研究[J].安徽农学院学报,1981,(2):41-48.
[8]Roberto Botta, Grazia Vergano, Giovanni Me. Floral biology and embryo development in chestnut [J]. Hort Sci, 1995, 30(6):1283-1286.
[9]Cevriye Mert, Arif Soylu. Flower and stamen structures of malefertile and male-sterile chestnut[J]. Soc Hortic Sci,2006, 131(6):752-759.
[10]张恒悦.板栗胚珠和胚的发育[J].山东农业大学学报,1986,17(2): 19-25.
[11]许慧玲,曹慧娟,谭晓风.板栗的胚胎学研究[J].北京林业大学学报,1988,10(1):10-16.
[12]袁德义,邹 峰,段经华.“金丝4号”枣大小孢子发生及雌雄配子体发育[J].经济林研究,2013,31(2):26-31.
[13]袁德义,邹 峰,李天生.油茶花芽分化及雌雄配子体发育的研究[J].中南林业科技大学学报,2011,31(3):65-70.
[14]Feng Zou, Su-Juan Guo. A Morphological and Histological Characterization of Male Flower in Chestnut (Castanea) Cultivar‘Yanshanzaofeng’[J]. Advance Journal of Food Science and Technology,2013,5(9):1192-1197.
[15]曾春霞,孙卫邦.三棱栎开花结实习性、小孢子发生及雄配子体发育[J].武汉植物学研究,2004,22(2):98-104.
[16]郑诚乐,俞晓曲,潘东明.锥栗雌配子体形成与胚胎发育[J].福建农林大学学报,2009,38(6):585-589.
[17]张林平,李保国,白志英.板栗雌花簇分化过程观察[J].果树科学,1999,16(4):280-283.
[18]辛培尧,孙正海,罗思宝,等.大麻花粉母细胞减数分裂染色体行为观察[J].河南农业科学,2008,(5):45-48.
[19]王永宏,孙益知,李方向.板栗花芽分化特点与调控技术研究进展[J].西北农业学报,2001,10(2):115-118.
[20]王云尊,马元考,陈维峰.珍惜板栗新品种浮来无花的性状及栽培技术[J].林业科技开发,2001,15(5):31-32.
[21]刘丽华.板栗雄性不育生理学机制研究[D].保定:河北农业大学,2007.
[22]冯永庆,秦 岭,杨东升,等.板栗短雄花序芽变的主要特征特性研究[J].北京农学院学报,2005,20(3):1-5.
[23]刘国彬,兰彦平.板栗雄性不育研究进展[J].中国农学通报,2011, 27(16):75-78.
Study on generation and development of megaspores, microspores, and female and male gametophytes in chestnut ‘Zunyu’
CHEN Fang, SU Shu-chai
(The Key Laboratory for Silviculture and Conservation of Ministry of Education, Beijing Forestry University,Beijing 100083, China)
In order to systematically understand the development process of male and female flowers in chestnut,taking a fine chestnut cultivar ‘Zunyu’ as materials, generation and development of megaspores, microspores, and female and male gametophytes were researched. The results showed that anther wall layers were composed of epidermis,endothecium, middle layer, and tapetal layer from outside to inner side, respectively, in which the tapetal layer belonged to glandular type. Cytokinesis of microspore belonged to simultaneous type. Most of the tetrads were typical tetrahedral type, and others were symmetrical type. Male gametophyte had only once meiosis process, mature pollens belonged to two-cell type. Embryo-sac developed into polygonum type, and only one embryo sac mother cell developed into embryo sac, and tetrad laid out in linear fashion. The entire development process was closely related to the external characters of male and female flowers. The optimal observation period formeiosis was when male inflorescence length was 10-17 cm and involucre diameter was 0.9-1.6 cm. No abnormal appearance was observed during development of microspores and male gametophyte in ‘Zunyu’, which indicated male gametes normal developed, were fertile, and could be used as pollinizers.
chestnut; male gametophyte; female gametophyte
10.14067/j.cnki.1003-8981.2015.03.008 http: //qks.csuft.edu.cn
2014-04-11
国家十一五科技支撑专题“板栗优质高产栽培技术研究”(2006BAD01A1703-1)。
陈 昉,硕士研究生。
苏淑钗,教授,博士。E-mail:sushuchai@sohu.com
陈 昉,苏淑钗.板栗‘遵玉’大小孢子发生及雌雄配子体发育研究[J].经济林研究,2015,33(3):44-49.
S664.2
A
1003—8981(2015)03—0044—06
[本文编校:闻 丽]
