无蜡质芥蓝突变型叶表面蜡质超微结构观察
2015-12-21张德双赵学志孙2张凤兰何洪巨汪维红苏同兵卢桂香
张德双赵学志孙 雷,2张凤兰何洪巨汪维红苏同兵卢桂香
于拴仓1余阳俊1赵岫云1
(1北京市农林科学院蔬菜研究中心,北京 100097;2烟台大学,山东烟台 264005)
无蜡质芥蓝突变型叶表面蜡质超微结构观察
张德双1赵学志1孙 雷1,2张凤兰1何洪巨1汪维红1苏同兵1卢桂香1
于拴仓1余阳俊1赵岫云1
(1北京市农林科学院蔬菜研究中心,北京 100097;2烟台大学,山东烟台 264005)
以亮叶无蜡质紫色芥蓝突变型紫.中花芥蓝和多蜡质紫色芥蓝野生型为试材,对芥蓝叶片发育过程中最外层的表面蜡质进行系统地观察,并对无蜡质突变型紫.中花芥蓝、多蜡质野生型芥蓝和普通绿色芥蓝(CK)的干物质量、VC、可溶性糖、粗纤维、蛋白质、花青苷等主要营养成分含量进行测定分析。结果表明:在叶片发育过程中,多蜡质野生型芥蓝的叶面蜡质明显多于无蜡质突变型紫.中花芥蓝;野生型芥蓝和紫.中花芥蓝叶片背面的蜡质均明显多于叶片腹面;野生型芥蓝叶片背面的蜡质退化速度明显慢于叶片腹面。无蜡质突变型紫.中花芥蓝的VC含量高于野生型芥蓝,而粗纤维(DW)含量低于野生型芥蓝,说明紫.中花芥蓝有利于改良现有芥蓝的营养品质,有望育成高VC含量、高花青苷含量、以生食为主的芥蓝品种。芥蓝叶片的无蜡质性状对多蜡质表现为隐性遗传,紫色对绿色表现为显性遗传。
芥蓝;无蜡质突变体;超微结构;花青苷
芥蓝(Brassica alboglabra L. H. Bailey)是十字花科芸薹属甘蓝种一年生或两年生草本植物,原产于我国华南地区,为我国著名的特产蔬菜之一。芥蓝栽培历史悠久,不仅深受国内消费者喜爱,还出口到日本及东南亚地区。芥蓝多以幼嫩、肉质的花薹和嫩叶为主要食用器官,肉质脆嫩清甜,营养丰富,富含硫代葡萄糖苷(Glucosinolate),尤其是4-甲基硫氧丁基硫苷(Zhang et al.,1994)。因此,芥蓝在营养品质、萝卜硫素提取及食品加工业等领域有着重要的研究价值。
植物叶片表皮细胞外覆盖表皮,表皮由3层明显的结构组成,最内层为表皮层(Cuticular layer,CL),由角质、角质层内蜡质和多糖组成;中间层为角质层本体(Cuticle proper,CP),由表面蜡质、角质层内蜡质和角质组成;最外层为平滑、结晶的表面蜡质(Epicuticular waxes,EW),具有很高的疏水性(图1)。植物叶表面的蜡质主要为特长链饱和脂肪酸的衍生物,在表皮细胞的内质网上合成,具有防止非气孔的水分散失、病虫害的侵入和太阳光辐射等生物学功能。

图1 植物叶表皮横断面示意图(Bernard & Joubès,2013)
自然界中已发现的植物叶表无蜡质突变体较多,研究者普遍关注控制蜡质合成的相关基因,但有关无蜡质突变体叶表面蜡质超显微结构的报道较少,尤其研究无蜡质芥蓝突变体的报道更少。普通芥蓝的叶表面均覆有不同程度的蜡质,当用手涂抹叶片表面后,蜡质便可部分减少。无蜡质亮叶型芥蓝最早由初莲香等(1998)报道,来源于无蜡质亮叶结球甘蓝,采用香港白花普通芥蓝为母本与无蜡质亮叶结球甘蓝父本杂交,F1与香港白花回交、再自交,经连续转育5代,并通过反复选择而育成。无蜡质亮叶型芥蓝丰富了芥蓝种质资源,在增加芥蓝产量等方面具有十分重要的意义。同时,无蜡质亮叶结球甘蓝突变体也具有一定的抗虫性(Lin et al.,1984;Stoner,1990)。
对蜡质突变体的叶表面超微结构进行研究,有助于了解突变型蜡质的发生机制以及蜡质的发育过程。本试验以亮叶无蜡质紫色芥蓝突变型紫.中花芥蓝和多蜡质紫色芥蓝野生型为试验材料,对芥蓝叶片最外层表面蜡质的发育过程及变化情况进行系统地观察和比较,以期在超显微水平上解析无蜡粉芥蓝突变体的叶表面结构;同时,测定了无蜡质紫色芥蓝突变型紫.中花芥蓝、多蜡质紫色芥蓝野生型和普通绿色芥蓝叶片的水分、干物质量、VC、可滴定酸、可溶性糖、粗纤维、蛋白质、花青苷等主要营养成分的含量,旨在为亮叶无蜡质紫色芥蓝在芥蓝育种中的应用及尽快选育出亮叶无蜡质紫色芥蓝一代杂种提供指导。
1 材料与方法
1.1 试验材料
亮叶无蜡质紫色芥蓝突变型紫.中花芥蓝来源于多蜡质台选中花芥蓝×紫甘蓝的F1与台选中花芥蓝(为父本)连续回交5代(每代均进行综合性状筛选)的回交群体中发现的突变型个体,自交后性状稳定遗传,主要表现为:植株叶表面光亮,有光泽,无蜡质覆盖,叶片薄而质地脆嫩,叶片、叶脉、菜薹均为紫色,且营养品质优良,又具有一定的抗性;多蜡质紫色芥蓝野生型是台选中花芥蓝×紫甘蓝的F1与台选中花芥蓝(为父本)连续回交5代(每代均进行综合性状筛选)获得的自交系,主要表现为:叶片多蜡粉覆盖,灰绿色,叶脉、菜薹均为紫色;普通绿色芥蓝(CK)为台选中花芥蓝人工自交授粉5代的自交系。上述材料均来源于北京市农林科学院蔬菜研究中心大白菜课题组。
2014年1月5日,在日光温室中播种芥蓝材料,3月15日定植到日光温室,开花后采用人工授粉方式获得自交种子。同时,以紫.中花芥蓝为母本,以普通多蜡质绿色芥蓝为父本进行杂交,获得F1种子。8月20日,在大棚中继续播种芥蓝材料,9月10日定植到四季青农场(露地),采用喷灌浇水,保持土壤湿润,9月30日追1次复合肥;10月20日调查紫.中花芥蓝突变型和野生型芥蓝、普通绿色芥蓝的叶片和菜薹颜色、蜡质有无等性状,初步研究芥蓝紫色和蜡质的遗传规律。
1.2 试验方法
1.2.1 超微结构观察 2014年4月,在日光温室中播种无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝,待芥蓝齐口期,分别选取无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝自交系4个发育阶段的叶片,即新叶、初生叶、次生叶和老叶。分别在中国农业科学院农产品加工研究所和北京生物技术研究中心电镜室进行观测,扫描电镜样品制作方法参考牟香丽等(2013)的方法并加以改进:用流水多次冲洗芥蓝新鲜叶片,再用ddH2O水清洗,以保证叶片表面干净;避开叶脉部分,用剪刀将叶片切成长方形小块,并用镊子摆放在覆有滤纸的干燥器中(滤纸上记有编号),将贮有四氧化锇的小瓶放在干燥器内,密闭熏蒸样品72 h,晾干;最后,采用E-1010(HITACHI)型离子溅射镀膜仪镀金膜,利用S-3400N和S-450(HITACHI)型扫描电子显微镜对样品叶片腹面、背面的蜡质结构进行观察。
为了加强四氧化锇熏蒸效果,四氧化锇需要装在开口的小容器内,并同样品一起放于密闭的干燥器中(图2);样品应该尽量小,这样不仅保证了样品熏蒸和干燥充分,而且样品也不容易卷曲,不会影响观测结果。

图2 四氧化锇熏蒸样品
1.2.2 营养品质测定 2013年10月,选取在露地种植的无蜡质突变型紫.中花芥蓝、多蜡质野生型芥蓝和普通绿色芥蓝(CK),每份材料3株,混合取样,送交农业部蔬菜种子质量监督检验测试中心,测定水分、干物质量、VC、可滴定酸、可溶性糖、粗纤维、蛋白质和花青苷等营养成分含量,样品的全部成分测定结果于2014年完成。
2 结果与分析
2.1 无蜡质突变型与多蜡质野生型芥蓝植物学性状比较
无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝自交系的植株性状见图3。紫.中花芥蓝植株叶表面光亮,有光泽,无蜡质,叶片、叶脉、菜薹均表现为紫色,叶片薄而质地脆嫩;野生型芥蓝叶片灰绿色,有蜡质,叶脉、菜薹均表现为紫色。而紫.中花芥蓝×普通多蜡质绿色芥蓝的F1表现为叶片紫色、叶面有蜡质。说明,芥蓝叶片无蜡质性状对多蜡质为隐性遗传、紫色性状对绿色为显性遗传。

图3 无蜡质突变型紫.中花芥蓝与多蜡质野生型芥蓝自交系植株
2.2 叶片超微结构观察
2.2.1 叶片腹面蜡质的比较 多蜡质野生型芥蓝的新叶、初生叶、次生叶和老叶无蜡质突变型腹面的蜡质均明显多于无蜡质突变型紫.中花芥蓝。新叶时,野生型芥蓝叶片腹面的蜡质为不规则颗粒和针状,紫.中花芥蓝为不规则颗粒(图4);初生叶时,野生型芥蓝叶片腹面的蜡质为圆柱状、片状晶体,具有规则的三维结构,紫.中花芥蓝为不规则颗粒和针状,聚集成团(图5);次生叶时,野生型芥蓝叶片腹面的蜡质为片状晶体,具有规则的三维结构,紫.中花芥蓝为不规则颗粒和针状,聚集成团,个体增大(图6);老叶时,野生型芥蓝叶片腹面的蜡质为颗粒状,聚集成团,形状不规则,紫.中花芥蓝为瘤状突起(图7),此时野生型芥蓝和紫.中花芥蓝叶片腹面的蜡质均退化,这与叶片衰老、功能丧失有关。由此可见,随着叶片的发育,野生型芥蓝和紫.中花芥蓝叶片腹面的蜡质逐渐增多、三维结构增强;至老叶时,因叶片老化,蜡质随之退化。

图4 无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝新叶腹面超微结构

图5 无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝初生叶腹面超微结构

图6 无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝次生叶腹面超微结构

图7 无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝老叶腹面超微结构
2.2.2 叶片背面蜡质的比较 多蜡质野生型芥蓝的新叶、初生叶、次生叶和老叶无蜡质突变型背面的蜡质均明显多于无蜡质突变型紫.中花芥蓝。新叶时,野生型芥蓝叶片背面的蜡质为管状、针状晶体,紫.中花芥蓝为片状、针状晶体,个体大,三维结构明显(图8);初生叶时,野生型芥蓝叶片背面的蜡质为片状、针状,三维结构增强,紫.中花芥蓝为片状、针状(图9);次生叶时,野生型芥蓝叶片背面的蜡质为片状、针状,三维结构进一步增强,紫.中花芥蓝为片状,聚集成团,开始退化(图10);老叶时,野生型芥蓝叶片背面的蜡质为密集的片状、针状,三维结构最强,紫.中花芥蓝为瘤状,退化严重。由此可见,随着叶片的发育,野生型芥蓝和紫.中花芥蓝叶片背面的蜡质逐渐增多,但紫.中花芥蓝的蜡质退化速度比野生型芥蓝更快。
2.2.3 叶片腹面、背面蜡质的比较 由图4~11可见,野生型芥蓝和紫.中花芥蓝叶片背面的蜡质多于腹面,背面蜡质退化的速度慢于腹面。野生型芥蓝叶片背面与腹面相比,背面蜡质退化速度极显著地慢于腹面,4个时期叶片背面的蜡质一直在增多、发育,在老叶时发育最好,而腹面的蜡质在老叶时开始退化;紫.中花芥蓝叶片背面与腹面相比,新叶背面的蜡质发育最好,随后开始退化,而腹面的蜡质在4个时期发育都比较差,老叶时完全退化。

图8 无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝新叶背面超微结构
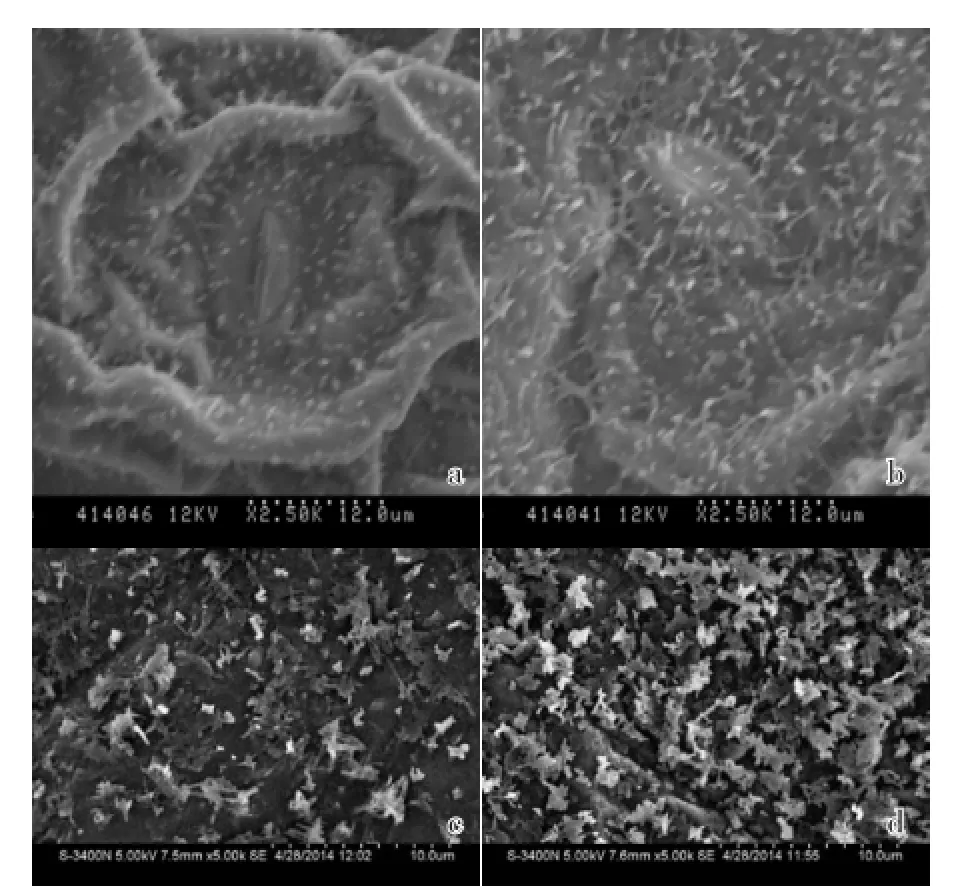
图9 无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝初生叶背面超微结构

图10 无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝次生叶背面超微结构

图11 无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝老叶背面超微结构
2.3 营养品质
由表1可见,无蜡质突变型紫.中花芥蓝和多蜡质野生型芥蓝的VC、花青苷含量均高于对照普通绿色芥蓝(CK),而紫.中花芥蓝和野生型芥蓝的花青苷含量相近;紫.中花芥蓝的VC含量、干物质量高于野生型芥蓝,可滴定酸、可溶性糖、粗纤维(DW)含量低于野生型芥蓝。由此可见,亮叶无蜡质突变型紫.中花芥蓝不仅增加了VC含量,而且降低了粗纤维(DW)含量;与对照相比,尤其是增加了花青苷含量。

表1 3种芥蓝主要营养品质
3 结论与讨论
3.1 野生型芥蓝的叶面蜡质多于无蜡质突变型紫.中花芥蓝
紫.中花芥蓝的叶片不仅具有蜡质结构,而且背面的蜡质尤为明显,因此紫.中花芥蓝叶片无蜡质是相对的。多蜡质紫色芥蓝野生型的新叶、初生叶、次生叶和老叶腹面、背面的蜡质均明显多于紫.中花芥蓝4个时期的叶面蜡质。位于叶片背面的蜡质,无论是多蜡质野生型芥蓝还是紫.中花芥蓝均多于腹面,这一结果与周小云(2006)、曾爱松等(2009)和牟香丽等(2013)报道的甘蓝叶片腹面的蜡质多于背面的结论不同,这可能与试验的材料及取材部位有关。芥蓝不包球,以幼嫩、肉质的花薹和嫩叶为主要食用器官(除老叶外,均可食用),且芥蓝叶片与茎形成的角度接近45°;而甘蓝包球,以叶球为食用器官(球叶以外的叶片,一般不可食用)。牟香丽等(2013)是在甘蓝的结球期和成熟期选取最内层外叶为试材,此时甘蓝的叶片为弯曲、抱合的球叶,不同于接近45°的芥蓝叶片。今后,应将亮叶无蜡质甘蓝、无蜡质亮叶芥蓝(初莲香 等,1998)和紫.中花芥蓝种植在同一条件下,再进一步比较三者叶片腹面、背面蜡质的异同。
3.2 紫.中花芥蓝有利于提高芥蓝品质
紫.中花芥蓝不仅提高了VC含量、降低了粗纤维(DW)含量,而且与普通绿色芥蓝相比,含有花青苷成分等优点更为突出,因此亮叶无蜡质紫色芥蓝突变型紫.中花芥蓝有利于改良现有芥蓝的营养品质。由于花青苷具有水溶性,VC遇高温容易降解等特点,建议育成的高营养品质紫色芥蓝新品种以生食为主,如凉拌、配菜等。
3.3 有关紫.中花芥蓝的选育工作
笔者已经获得了紫.中花芥蓝×普通多蜡质绿色芥蓝的6个世代种子。2015年秋季将在露地种植这6个世代,调查每个群体各单株紫色和蜡质的有无等分离情况,进一步开展蜡质、紫色等遗传规律研究;同时,利用紫.中花芥蓝×普通多蜡质绿色芥蓝配制的BC1回交群体,采用分离群体分组分析法(bulked segregant analysis,BSA),开展与紫色、蜡质相关基因的初步定位研究,并建立分子辅助育种技术体系,在苗期能够区分纯合、杂合蜡质和紫色基因型,为快速育成亮叶无蜡质且叶片、叶脉和菜薹均为紫色的芥蓝一代杂种提供依据。
初莲香,王秋艳,王英明,王永昌.1998.无蜡粉亮叶芥蓝的选育及利用.中国蔬菜,(4):30-31.
牟香丽,王超,王帅.2013.甘蓝无蜡粉突变体叶表皮蜡质超微结构观察.中国蔬菜,(4):32-37.
曾爱松,刘玉梅,方智远.2009.甘蓝耐裂球性与叶表面微形态及细胞组织结构的关系研究.华北农学报,(2):41-45.
周小云.2006.水稻叶表皮蜡质发育及蜡质相关转录因子基因OsWTF1和OsWTF2的克隆与鉴定〔博士论文〕.长沙:湖南农业大学.
Bernard A,Joub è s J.2013.Arabidopsis cuticular waxes:advances in synthesis,export and regulation.Prog Lipid Res,52(1):110-129.
Lin J,Dickson M H,Eckenrode C J.1984.Resistance of Brassica lines to the diamondback moth(Lepidoptera:Yponomeutidae)in the field and inheritance resistance.Journal of Economic Ent omology,77:1293-1296.
Stoner K A.1990.Glossy leaf wax and plant resistance to insects in Brassica oleracea under natural infestation.Environmental Entomology,19(3):730-739.
Zhang Y,Kensler T W,Cho C G,Posner G H,Talalay P.1994.Anticarcinogenic activities of sulforaphane and structurally related synthetic norbornyl isothiocyanates.Proc Natl Acad Sci,91(8):3147-3150.
Ultra-microstructure of Epicuticular Wax on Waxless Mutant of Chinese Kale(Brassica alboglabra L. H. Bailey)
ZHANG De-shuang1,SUN Lei1,2,ZHANG Feng-lan1,HE Hong-ju1,YU Shuan-cang1,YU Yang-jun1,ZHAO Xiu-yun1,ZHAO Xue-zhi1,WANG Wei-hong1,SU Tong-bing1,LU Gui-xiang1
(1Beijing Vegetable Research Center,Beijing Academy of Agricultural and Forestry Sciences,Beijing 100097,China;2Yantai University,Yantai 264005,Shandong,China)
Epicuticular waxes of leaves at 4 developing stages in Chinese kale(Brassica alboglabra L. H. Bailey)waxless mutant named‘Zi.zhonghuajielan’and wild type(WT)were observed by scanning electron microscope(SEM).Meanwhile,crude weight,VC,soluble sugar,crude fiber,protein and anthocyanins were measured. The results showed that epicuticular waxes at 4 growth periods of wild type were apparently higher than those of mutant. Epicuticular waxes at leaf abaxial of mutant and WT were higher than those of leaf adaxial.Epicuticular waxes at leaf abaxial of WT degenerated slowly than those of leaf adaxial. VC in waxless mutant was higher than that of WT,and dry weight of crude weight in waxless mutant was lower than that of WT.Thus,waxless mutant in Chinese kale can supply higher nutrition of VC and anthocyanins.Gene controlled wax biosynthesis in Chinese kale waxless mutant was recessive,while gene or genes controlled anthocyanins biosynthesis were dominant,compared to WT.Furthermore brilliant green,waxless and purple Chinese kale uncooked is more suitable for eating.
Chinese kale(Brassica alboglabra L. H. Bailey);Waxless mutant;Ultra-microstructure;Anthocyanins
张德双,男,博士,研究员,专业方向:蔬菜遗传育种及分子生物学,E-mail:zhangdeshuang@nercv.org
2015-07-14;接受日期:2015-10-08
北京市农林科学院科技创新能力建设专项(KJCX20140111-3),大宗蔬菜产业技术体系专项(CARS-25-A-11),北京市叶类蔬菜创新团队资助项目(blvt-01)
