红曲霉发酵高温豆粕高产可溶性多肽
2015-12-20倪春蕾程建军朱秀清
石 琳,尹 园,王 帅,齐 惠,倪春蕾,程建军,*,朱秀清
(1.东北农业大学食品学院,黑龙江 哈尔滨 150030;2.黑龙江省大豆技术开发研究中心,黑龙江 哈尔滨 150086)
红曲霉发酵高温豆粕高产可溶性多肽
石 琳1,尹 园1,王 帅1,齐 惠1,倪春蕾1,程建军1,*,朱秀清2
(1.东北农业大学食品学院,黑龙江 哈尔滨 150030;2.黑龙江省大豆技术开发研究中心,黑龙江 哈尔滨 150086)
通过利用红曲霉发酵高温豆粕产生可溶性多肽,确定菌株对变性蛋白质有一定的分解作用;并采用紫外诱变红曲霉,经初筛和复筛,最终筛选出能高产可溶性多肽的菌株。结果表明:红曲霉发酵可分解高温豆粕中的蛋白质,并且产生分子质量介于7.8~20.1 kD的可溶性多肽。发酵120 h可产生(13.41±0.20) mg/mL的可溶性多肽,是原豆粕的3.60 倍。15 W紫外线灯35 cm处,搅拌条件下照射50 s,红曲霉的致死率为(82.70±2.20)%。在此诱变条件下得到一株高产可溶性多肽的突变红曲霉0501100菌株。该菌株在发酵豆粕96 h时产生的可溶性多肽含量达到(17.20±0.18) mg/mL,是原菌株在同等发酵时间条件下产生可溶性多肽的1.47 倍,是原豆粕的4.61 倍,缩短了发酵时间并且有较好的遗传稳定性。
红曲霉;高温豆粕;可溶性多肽
大豆经浸提脱油后的碎片状或粗粉状的副产品--豆粕,是目前使用最多、最广泛的植物性蛋白质饲料原料。其粗蛋白含量高(40%~45%),氨基酸组成平衡,营养价值较高[1],因此豆粕蛋白质的高效利用就显得更为重要。高温豆粕中蛋白质的变性程度很大,其溶解性等性质较大豆分离蛋白差,可通过直接酶解法[2-3]或发酵法[1,4]生产大豆肽的方式加以利用,而发酵法既能去除大豆中各种致敏原[5]、增加蛋白质和氨基酸总含量[6],还能把蛋白酶的微生物生产和大豆肽的酶解生产结合在一起,降低了大豆多肽的生产成本,应用前景较好。
红曲霉(Monascus)在我国主要是用于制造传统食品,在酿酒、发酵食品、食品色素、中药等方面有着广泛应用[7]。它能利用多种有机碳源,可产生多种酶类,并有不少菌株可产生高活性蛋白酶[8]。诱变处理可使得微生物的突变机率增大,加上特定培养基的定向筛选,可筛选出高产某种特定产物(如蛋白酶)的遗传稳定菌株用于工业发酵。
关于红曲霉的研究主要集中在红曲色素[9-11]以及其他次级代谢产物,其中包括了具有降血脂功效的洛伐他汀(lovastatin)[12]、降血压功效的γ-氨基丁酸[13]、增强免疫力的红曲多糖[14]、VD的前体麦角固醇(ergosterol)[15],而对于红曲霉作为霉菌自身产生的蛋白酶的利用却较少。本研究利用高温脱脂后的大豆豆粕作为原料,期望通过初筛和复筛分离出能高效分解蛋白的优良红曲霉菌株,将变性豆粕蛋白转化为易于人体消化吸收的多肽,为进一步研究提供依据。
1 材料与方法
1.1 菌种与试剂
紫红曲霉(Monascus purpureus)3.5833 中国普通微生物菌种保藏管理中心(China General Microbiological Culture Collection Center,CGMCC)。
豆粕 黑龙江省阳霖油脂集团有限公司;偶氮酪蛋白(azocasein) 美国Sigma公司;脱脂奶粉 市售;其他试剂均为国产分析纯。
1.2 培养基
斜面培养基(麦芽汁琼脂培养基):蛋白胨4 g/L、葡萄糖10 g/L、酵母浸粉3 g/L、麦芽粉10 g/L、琼脂粉13 g/L,pH值自然。
液体培养基:葡萄糖20 g/L、蛋白胨15 g/L、酵母提取物5 g/L,pH值自然。
初筛培养基[16]:脱脂奶粉25 g,溶于250 mL水中;琼脂10 g,溶于250 mL水中,分开灭菌,待冷却至45~50 ℃后混合。
复筛培养基(液态发酵培养基):高温豆粕与水之比为1∶9(m/V),pH值自然。
基础发酵条件:80 g/500 mL的装料量,107CFU/mL的接种量,(28.0±0.5)℃、180 r/min的摇床培养条件。
除初筛培养基采用115 ℃灭菌15 min外,其他培养基均采用121 ℃灭菌20 min后备用。
1.3 仪器与设备
5430R小型台式高速离心机 艾本德中国有限公司;SW-CJ-1D型超净工作台 苏州净化设备有限公司;HZQ-F100振荡培养箱 哈尔滨市东联生化仪器有限公司;TU-1800紫外-可见分光光度计 北京普析通用仪器有限责任公司。
1.4 红曲霉发酵高温豆粕产可溶性多肽的研究
1.4.1 高温豆粕中蛋白质的测定
采用国标GB 5009.5-2010《食品中蛋白质的测定》中的凯氏定氮法测定豆粕中蛋白质含量。
粉碎高温豆粕后过80 目筛,取筛下物测定原豆粕中粗蛋白含量。在室温条件下,将筛下物按照料水比1∶9,180 r/min摇床振荡提取2 h,而后7 000×g离心15 min,取上清液,经0.45 μm微孔滤膜过滤后,测定原豆粕中可溶性蛋白含量。
将筛下物在121 ℃热处理20 min后,7 000×g离心15 min,取上清液,经0.45 μm微孔滤膜过滤后,测定加热处理后高温豆粕中可溶性蛋白含量。
1.4.2 生长曲线的测定
将活化后的菌种斜面制成孢子悬液,接种于液体培养基,按照40 mL/250 mL的装料量,106CFU/mL的接种量,在(28.0±0.5)℃、180 r/min的摇床培养条件下振荡培养192 h。每24 h取培养基梯度稀释涂布,每个稀释度涂3 个麦芽汁琼脂平板,(28.0±0.5)℃恒温箱中培养40~48 h,按照菌落总数计数法测定活菌数。
1.4.3 总蛋白酶活力的测定
取液体培养基,4 ℃、12 000×g离心10 min,而后取少量上清液,参照Hirano等[17]方法测定总蛋白酶活力。一个酶活力单位定义为在37 ℃每小时吸光度升高0.01为1 U。
1.4.4 红曲霉发酵高温豆粕发酵液的制备
将红曲霉菌株孢子悬液接种于液体培养基,培养72 h左右。接种于液态发酵培养基,发酵数天后7 000×g离心15 min,取上清液,经0.45 μm微孔滤膜过滤后,测定发酵液中可溶性多肽含量和分子质量。
1.4.5 可溶性多肽含量的测定
可溶性多肽的含量以酸可溶性多肽计算,取1 mL发酵液加入1 mL质量分数10%的三氯乙酸(trichloroacetic acid,TCA)溶液,以去除酸不溶性的蛋白质和长链肽的沉淀,取上清液根据双缩脲法[18]测定酸可溶性多肽的含量。
将恒质量的酪蛋白作为标准蛋白质绘制标准曲线。
1.4.6 可溶性蛋白质(多肽)分子质量分布的测定
采用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodiam dodecyl sulfate-polyacrylamide gelelectrophoresis,SDS-PAGE)法对可溶性蛋白质(多肽)分子质量分布进行测定,参考并修改Laemmli[19]和Schägger[20]的方法。
将处理后的发酵液于14 000×g离心10 min后测定可溶性蛋白分子质量分布。
取1 mL发酵液加入1 mL 10%的TCA溶液,室温放置30 min后5 000×g离心10 min,取上清液14 000×g离心10 min后测定可溶性多肽分子质量分布。
1.5 红曲霉菌株的筛选
1.5.1 紫外诱变条件的确定
调整孢子浓度为106CFU/mL。取孢子悬液5 mL置于15 W紫外线灯35 cm处,磁力搅拌条件下照射10~70 s,而后稀释液涂布于麦芽汁琼脂平板,于(28.0±0.5)℃恒温避光培养48 h后记录菌落数。以未经紫外处理的孢子悬液稀释涂布平板作为对照。
计算致死率,选择致死率在80%左右的紫外照射时间作为最适诱变剂量。

1.5.2 初筛
将最适诱变剂量处理后的孢子悬液接种于液体培养基,按照基础培养条件培养24 h后,稀释涂布于麦芽汁琼脂平板上,在(28.0±0.5)℃条件下培养48 h后,将各菌株点种于初筛培养基上,以未处理的原菌株作为对照。每12 h用卡尺测量每株菌的水解圈直径和菌落直径,将其比值(水解圈直径与菌落直径的比值,HC值)显著大于对照的突变菌株转接到麦芽汁琼脂斜面上培养7 d,以备复筛使用。

1.5.3 复筛
将初筛菌株制成孢子浓度为107CFU/mL的孢子悬液,接种于液体培养基培养72 h左右。而后按照基础发酵条件接种于复筛培养基,将培养数天后的液体培养基按照基础发酵条件接种于液态培养基中,发酵数天后取发酵液7 000×g离心15 min后,取上清液,经0.45 μm微孔滤膜过滤后,测定发酵液中可溶性多肽含量,将其多肽含量显著高于原菌株的突变菌株定义为高产菌株。
1.5.4 遗传稳定性的测定
筛选出的突变菌株经4~6 次复筛培养基传代,测定培养96 h后发酵液中可溶性多肽含量的变化,比较其遗传稳定性。
1.6 数据处理与分析
每个指标测定重复3 次。并采用SPSS 17统计软件进行单因子方差分析,结果以±s表示。
2 结果与分析
2.1 加热处理前后高温豆粕中可溶性蛋白含量变化的比较
采用GB 5009.5-2010中的凯氏定氮法测定高温豆粕中的粗蛋白、可溶性蛋白及加热处理后可溶性蛋白含量。高温豆粕中粗蛋白含量为(39.26±0.77)%,虽然含量很高,但可溶性蛋白的含量较少,仅为(8.70±0.09)%,占粗蛋白含量的22.16%左右,这为红曲霉发酵豆粕产生更多可溶性蛋白提供了条件。加热处理后的高温豆粕中可溶性蛋白含量有所上升,达到(19.19±0.14)%,这是由于高温高压处理促使不溶性蛋白部分转变成可溶性蛋白。
2.2 菌株的生长曲线
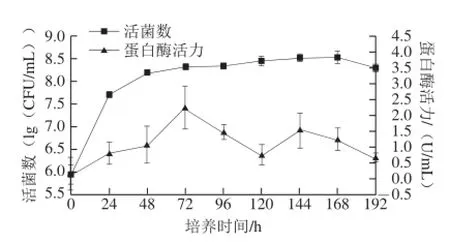
图1 红曲霉菌株生长曲线Fig.1 Growth curve of Monascus purpureus
由图1可知,红曲霉在液体培养基中培养48~72 h后达到对数生长末期,而后进入稳定期,此时活菌数为1.48×108CFU/mL(lg活菌数为8.32)。由于在对数生长末期时,菌体数量最多,而进入稳定期以后,菌体数量趋于稳定,不再增加。因此在微生物实验中,一般采用对数生长末期的菌悬液作为种子液接种到筛选培养基中[21]。与此同时,菌株的蛋白酶活力在0~72 h呈上升趋势,而后又下降,在72 h达到2.18 U/mL,更有利于豆粕中蛋白质的分解,因此采用培养72 h后的液体培养基作为种子接种到菌株的液态培养基上。
2.3 红曲霉发酵高温豆粕的研究
2.3.1 发酵对高温豆粕中可溶性蛋白(多肽)分子质量的影响
通过SDS-PAGE分析红曲霉发酵高温豆粕后可溶性蛋白和多肽分子质量的变化,其结果如图2所示。加热处理后的高温豆粕中,分子质量大于43.0 kD的可溶性蛋白明显减少,而分子质量小于14.4 kD的可溶性蛋白有所增加,红曲霉的发酵作用可使分子质量在31.0~43.0 kD的可溶性蛋白彻底分解(图2A中泳道3)。随着菌株发酵时间的延长,发酵24、48 h时发酵液中分子质量小于31.0 kD的可溶性蛋白明显减少(图2A中泳道3和4),而72、96 h时又明显增加(图2A中泳道5和6)。这是因为0~72 h内红曲霉处于生长阶段,可溶性蛋白在红曲霉分泌的蛋白酶作用下,分解成小分子的蛋白质用于自身生长代谢。而后,蛋白酶作用于不溶性的蛋白质,分解产生的可溶性蛋白质含量增加,并且大于自身生长代谢所消耗的量,产生大量的积累。
在这个过程中可溶性多肽的含量也在不断地积累,如图2B所示,红曲霉发酵高温豆粕可大量产生分子质量7.8~20.1 kD的可溶性多肽,且大部分集中在14.4 kD左右。在发酵120 h时分子质量为14.4~20.1 kD的可溶性多肽明显减少(图2B中泳道6),这说明大分子的多肽在不断地分解成小分子的多肽。
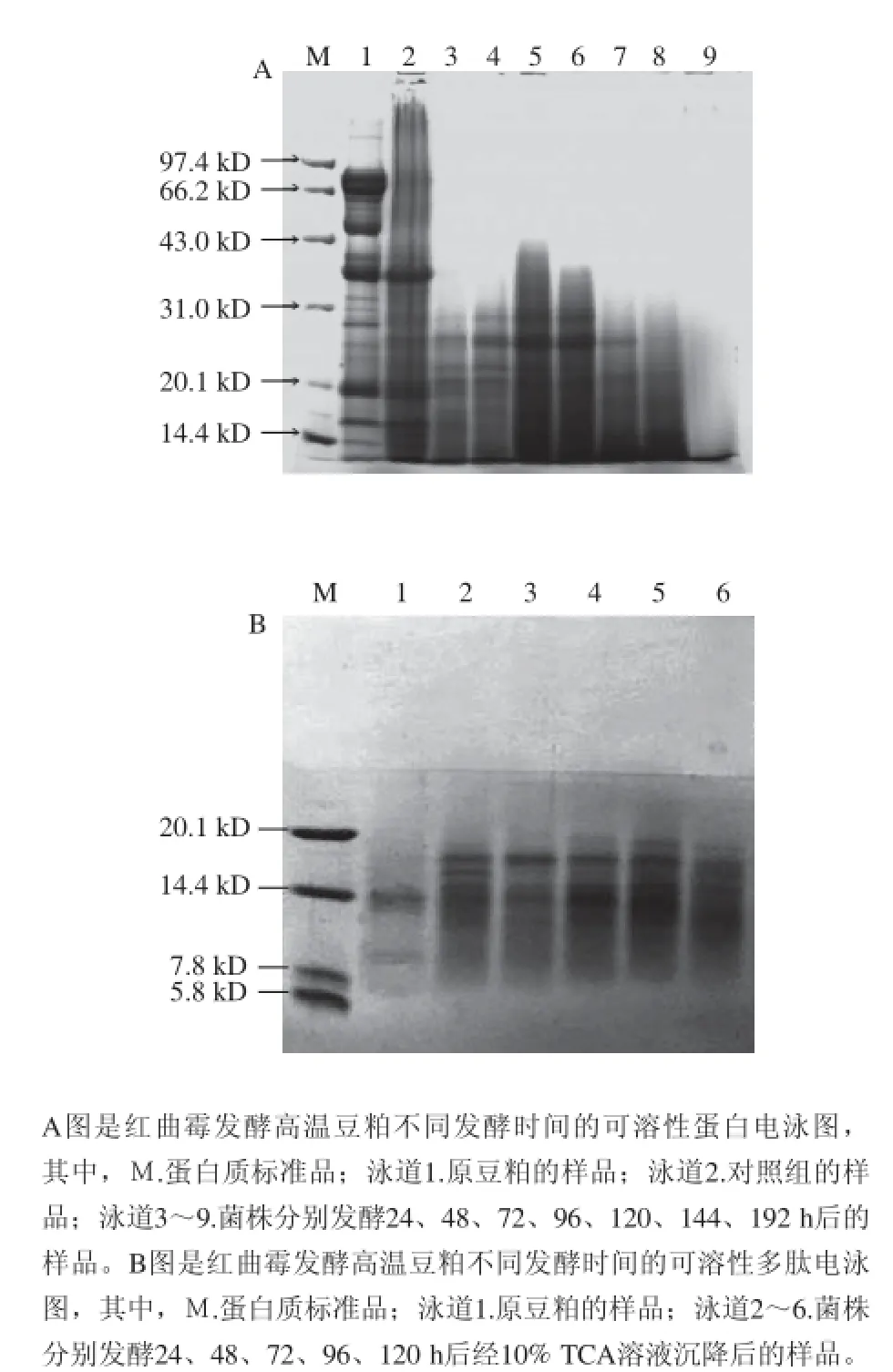
图2 红曲霉发酵高温豆粕产可溶性蛋白(多肽)的电泳图Fig.2 Electrophoresis patterns of soluble protein (polypeptide) from Monascus purpureus fermented high-temperature soybean meal
2.3.2 发酵对高温豆粕中可溶性多肽含量的影响
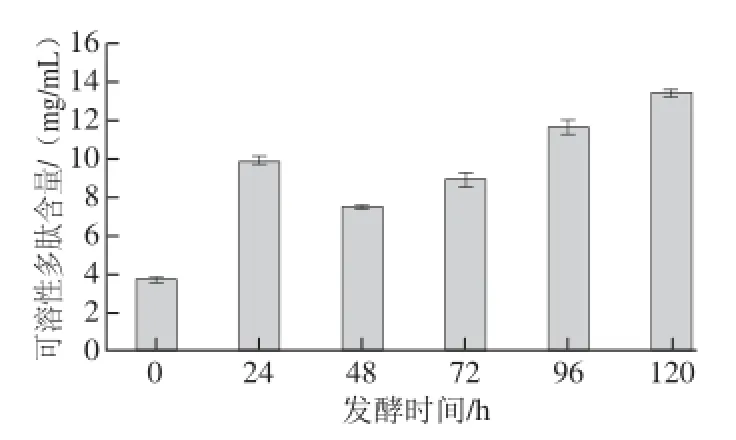
图3 红曲霉发酵高温豆粕对可溶性多肽含量的影响Fig.3 Effect of fermentation time on the content of soluble protein in high-temperature soybean meal fermented by Monascus purpureus
图3为对2.3.1节中可溶性多肽(图2B)的定量分析。原豆粕中可溶性多肽含量为(3.73±0.03) mg/mL,而发酵24 h时达到(9.90±0.20) mg/mL,其可溶性多肽含量显著增加。随着发酵时间的延长,发酵液中可溶性多肽含量呈先下降而后上升的趋势,在48~120 h的发酵过程中,可溶性多肽含量不断地上升,这与图2B显示的结果一致。当发酵48 h时达到最低(7.46±0.05) mg/mL,之后120 h时上升至(13.41±0.20) mg/mL,是原豆粕的3.60 倍,这说明红曲霉发酵高温豆粕能显著地提高可溶性多肽的含量。
2.4 紫外诱变剂量的选择
采用紫外线作为诱变剂,目的是增加红曲霉菌株突变率,提高其产蛋白酶的变异幅度。紫外照射时间与致死率的关系见图4。在该条件下辐照10 s,红曲霉菌株致死率达到(22.97±12.58)%,随着辐照时间的不断延长,致死率逐渐增加,当照射时间为70 s时已达到(95.14±0.53)%。通常随着辐照剂量的增加,微生物的突变率升高,但达到一定程度后,辐照剂量再增加突变率反而下降,研究结果证明,在辐照剂量偏低、致死率偏低的情况下,紫外线等诱变剂引起的正变率高,更容易分离到高产菌株[22],因此本实验采用致死率在80%左右的诱变剂量作为诱变剂的最适剂量。即在15 W紫外线灯35 cm处,搅拌照射50 s为最适剂量,此时致死率为(82.70±2.20)%。
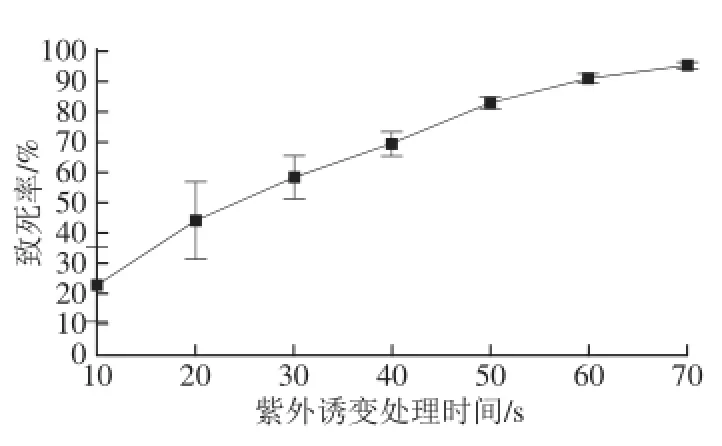
图4 紫外线诱变红曲霉菌株结果Fig.4 Mutagenesis of Monascus purpureus by ultraviolet light
2.5 突变菌株筛选结果
2.5.1 初筛结果
将最适诱变剂量处理后的菌株点种于初筛培养基上,菌株产生的蛋白酶可水解培养基中酪蛋白,由于HC值与蛋白酶活力之间成正相关[23],以产生的水解圈与菌落直径的比值(HC值)来衡量菌株分解蛋白质的能力,即是蛋白酶活力的比较。菌株在初筛培养基上培养48 h时才能准确测定水解圈与菌落的直径,并且随着培养时间的延长,菌落不再生长或是出现形态上的变化,不再适合继续培养和测定。因此,实验的测定结果采取培养48~144 h内、每12 h测定的水解圈与菌落直径的比值表示,初筛菌株HC值测定结果如表1所示。

表1 初筛菌株HC值Table 1 HC values of mutant strains in initial screening experiments
如表1所示,各菌株随着培养时间的延长,HC值主要呈不断减小的趋势。其中0310717菌株的HC值在培养48~144 h的测量时间范围内均显著高于原菌株的HC值。而在培养72~132 h内,除72 h时0409300、0501100、0501132菌株和84 h时0200104菌株的HC值外,其他时间的突变菌株的HC值均显著高于同时间的原菌株HC值,并且96 h时0310717菌株的HC值是同时间的原菌株1.25 倍。这说明诱变后得到的突变菌株在初筛培养基上培养72~132 h内产生的蛋白酶活力高于原菌株。
2.5.2 复筛结果

表2 各菌株发酵后样品中可溶性多肽含量Table 2 Content of soluble polypeptide in the sample subjected to fermentation by each strain mg/mL
将初筛得到的红曲霉突变菌株接种到复筛培养基中发酵120 h,各发酵液中可溶性多肽含量测定结果如表2所示。随着发酵时间的延长,可溶性多肽的含量呈现先下降的趋势,在发酵48 h时到达最低点,其中0501100菌株在发酵48 h时可溶性多肽含量最低,仅为(5.39±0.45) mg/mL,但仍是原豆粕的1.45 倍。而后可溶性多肽的含量又逐渐升高,在发酵96 h时部分菌株发酵液中可溶性多肽含量达到最高点,之后又再次下降;而另一部分的菌株仍然继续上升。这说明各菌株之间产生蛋白酶的时间和酶的活力不同,导致分解蛋白质产生多肽含量的不同,并且达到最高含量的时间也是不同的。各菌株发酵液中可溶性多肽含量最高的是0501100菌株发酵96 h时,为(17.20±0.18) mg/mL,显著高于其他菌株(P<0.05),是同等发酵条件下原菌株发酵样品的1.47 倍,是原豆粕的4.61 倍。因此选用0501100菌株进行遗传稳定性实验。
2.6 突变菌株遗传稳定性

表3 0501100菌株各代发酵96 h样品中可溶性多肽含量Table 3 Content of soluble polypeptide in the sample fermented by each generation of mutant strain 0501100 for 96 h
复筛得到的0501100菌株经过连续5 代接种到复筛培养基上发酵培养后,其发酵样品中可溶性多肽含量测定结果如表3所示。该菌株连续5 代发酵样品中可溶性多肽含量变化不显著(P>0.05),因此该菌株有较好的遗传稳定性。
3 结 论
采用红曲霉液态发酵高温豆粕,将其中变性蛋白转化成了可溶性蛋白,增加了高温豆粕中蛋白质的利用率,并且可溶性多肽的含量显著增加,发酵120 h时可到达(13.41±0.20) mg/mL,是原豆粕的3.60 倍。这为红曲霉发酵高温豆粕生产可溶性多肽提供了依据。
在该发酵条件下,筛选出一株高效分解高温豆粕中变性蛋白的0501100菌株,同时该菌株在发酵96 h后产生的可溶性多肽含量达到(17.20±0.18) mg/mL,是原菌株同等发酵时间条件下可溶性多肽的1.47 倍,是原豆粕的4.61 倍。缩短了发酵时间并且有较好的遗传稳定性。
[1] 朱秀清, 王玲. 微生物发酵高温豆粕菌种筛选及发酵工艺优化[J].食品与发酵工业, 2012, 38(4): 82-88.
[2] 张静, 李理. Alcalase蛋白酶酶解高温豆粕制备水溶性大豆多肽[J].食品工业科技, 2012, 33(10): 212-215.
[3] 许晶, 江连洲, 李杨, 等. 酶水解高温豆粕制备高水解度大豆肽的研究[J]. 食品与发酵工业, 2009, 35(4): 98-102.
[4] 叶倩. 发酵豆粕深度酶解生产低分子大豆多肽[D]. 武汉: 华中农业大学, 2009.
[5] 黄颖. 微生物发酵法降解大豆致敏原的研究[D]. 无锡: 江南大学, 2012.
[6] 马文强, 冯杰, 刘欣. 微生物发酵豆粕营养特性研究[J]. 中国粮油学报, 2008, 23(1): 128-131.
[7] 李钟庆, 杨晓暾, 郭芳. 综观红曲与红曲菌[M]. 北京: 中国轻工业出版社, 2009: 1-3.
[8] 袁国兴. 红曲米中蛋白酶、酯化酶的活性研究[D]. 济南: 山东轻工业学院, 2012.
[9] MEINICKE R M, VENDRUSCOLO F, MORITZ D E, et al. Potential use of glycerol as substrate for the production of red pigments byMonascus ruber in submerged fermentation[J]. Biocatalysis and Agricultural Biotechnology, 2012, 1(3): 238-242.
[10] PRAJAPATI V S, SONI N, TRIVEDI U B, et al. An enhancement of red pigment production by submerged culture of Monascus purpureus MTCC 410 employing statistical methodology[J]. Biocatalysis and Agricultural Biotechnology, 2014, 3(2): 140-145.
[11] KIM C, JUNG H, KIM J H, et al. Effect of Monascus pigment derivatives on the electrophoretic mobility of bacteria, and the cell adsorption and antibacterial activities of pigments[J]. Colloids Surfaces B: Biointerfaces, 2006, 47(2): 153-159.
[12] SERAMAN S, RAJENDRAN A, THANGAVELU V. Statistical optimization of anticholesterolemic drug lovastatin production by the red mold Monascus purpureus[J]. Food and Bioproducts Processing, 2010, 88(2): 266-276.
[13] 胡珊, 王小波, 戴甄, 等. 产γ-氨基丁酸红曲菌种的筛选及其发酵工艺条件研究[J]. 食品与发酵科技, 2011, 47(4): 71-74.
[14] 张建峰, 昌友权, 陈光, 等. 红曲多糖的免疫活性研究[J]. 食品科学, 2008, 29(2): 391-393.
[15] 葛锋, 王艳, 王剑平, 等. 红曲中主要活性成分的研究[J]. 昆明理工大学学报: 自然科学版, 2012, 37(2): 61-64.
[16] 赵寿经, 吴晓艳, 刘丽, 等. 产酸性蛋白酶微生物的筛选及其在玉米淀粉生产中的应用[J]. 食品与发酵工业, 2011, 37(7): 68-72.
[17] HIRANO S, KATO J Y, OHNISHI Y, et al. Control of the Streptomyces subtilisin inhibitor gene by adpa in the a-factor regulatory cascade in Streptomyces griseus[J]. Journal of Bacteriology, 2006, 188(17): 6207-6216.
[18] COTTON F, DELOBBE E, GULBIS B. Assessment of a Biuret method without concentration step for total protein determination in cerebrospinal fl uid[J]. Clinical Biochemistry, 1997, 30(4): 313-314.
[19] LAEMMLI U K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4[J]. Nature, 1970, 227: 680-685.
[20] SCHÄGGER G V J H. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa[J]. Analytical Biochemistry, 1987, 166(2): 368-379.
[21] 马宏颖. 中性蛋白酶高产菌株的诱变选育及发酵条件研究[D]. 保定: 河北农业大学, 2008.
[22] 居乃琥. 酶工程手册[M]. 北京: 中国轻工业出版社, 2011: 335.
[23] 赵伟华, 程建军, 杨秋萍, 等. 高温变性豆粕发酵菌株的选育[J]. 食品工业科技, 2012, 33(1): 152-154.
Production of Soluble Polypeptide from Fermented High-Temperature Soybean Meal by Monascus purpureus
SHI Lin1, YIN Yuan1, WANG Shuai1, QI Hui1, NI Chunlei1, CHENG Jianjun1,*, ZHU Xiuqing2
(1. College of Food Science, Northeast Agricultural University, Harbin 150030, China; 2. Soybean Technology Research and Development Center in Heilongjiang Province, Harbin 150086, China)
The molecular weight and content of soluble polypeptide in high-t emperature soybean meal subjected to Monascus purpureus fermentation were determined to ascertain whether denatured protein could be decomposed during the fermentation process. The strain was mutagenized with ultraviolet light. The mutant strains were inoculated into hightemperature soybean meal and scree ne d twice for the highest yield of soluble polypeptides. The results showed that proteins were decomposed in high-temperature soybean meal during Monascus purpureus fermentation, producing soluble polypeptides with molecular weight of 7.8-20.1 kD. The yield of soluble polypeptide from fermented soybean meal after 120 h of fermentation was (13.41 ± 0.20) mg/mL, which was 3.60 times higher than that from the original soybean meal. Monascus purpureus was irradiated by a 15 W UV lamp at a distance of 35 cm with stirring for 50 s, leading to a death rate of (82.70 ± 2.20)%. The mutant 0501100 could produce relatively high yield of soluble polypeptide under these conditions. The yield of soluble polypeptide from soybean meal fermented by the mutant strain 0501100 for 96 h was (17.20 ± 0.18) mg/mL, which exhibited a 1.47- and 4.61-fold increase when compared with that obtained from the original strain under the same fermentation conditions and the original soybean, respectively. Moreover, the fermentation time was shortened and the mutant strain had a good genetic stability.
Monascus purpureus; high-temperature soybean meal; soluble polypeptide
TS201.1
A
1002-6630(2015)19-0137-06
10.7506/spkx1002- 6630-201519024
2014-11-22
“十二五”国家科技支撑计划项目(2014BA22B00);黑龙江省应用技术研究与开发计划项目(GC13B208)
石琳(1989-),女,硕士研究生,研究方向为粮食油脂及植物蛋白工程。E-mail:sl89115@hotmail.com
*通信作者:程建军(1969-),男,教授,博士,研究方向为农产品加工。E-mail:cheng577@163.com
