唐家河国家级自然保护区扭角羚环境容纳量
2015-12-02官天培谌利民符建荣宋延龄
官天培,谌利民,符建荣,宋延龄
(1.绵阳师范学院生态安全与保护四川省重点实验室,四川绵阳 621000;2.四川唐家河国家级自然保护区,四川青川 628100;3.四川省林业科学研究院,四川成都 610081;4.中国科学院动物研究所,北京 100101)
扭角羚(Budorcas taxicolor),又称羚牛、牛羚、盘羊、野牛等,隶属偶蹄目,牛科,羊亚科,扭角羚属(吴家炎1990,胡锦矗2007),是仅分布在亚洲山地森林的珍稀有蹄类,被IUCN认定为易危,红色物种名录附录Ⅱ,国家一级重点保护野生动物(汪松2009,宋延龄等2001)。四川扭角羚是目前分布面积最广,现存种群数量最多的亚种(胡锦矗1984)。岷山是四川扭角羚的主要分布产区,而唐家河自然保护区是岷山山系扭角羚遇见率最高的区域(Guan et al.2015)。扭角羚是唐家河国家级自然保护区的重点保护对象。
一般而言,在栖息地完好的情况下,野生动物数量会随着时间推移持续增长(Dasmann,1964)。但是,这种增长会受到天敌捕食、疾病和容纳量的制约。老年和患病个体将会由自然死亡和天敌捕食淘汰,过高的种群密度将可能通过疾病传播和食物短缺两个途径加以控制(孙儒泳2001)。此外,极端气候引发的疾病和食物短缺也可能导致动物的死亡。在唐家河,扭角羚是该区域内体型最大的哺乳动物,且缺乏天敌捕食(胡锦矗等2004)。由于保护力度不断的提升,区内扭角羚数量呈现逐年增长的态势。然而,保护区在近年冬末春初的监测中发现了较多的死亡扭角羚,且与偷盗猎和天敌捕食后的痕迹特征不符(虎绍勇等2011)。据生态学原理和扭角羚的生态习性分析,疾病和饥饿均有可能是扭角羚异常死亡的原因。但是这两种因素的根源都与种群数量过大而导致的密度制约效应相关(孙儒泳2001)。因此,我们有必要评价唐家河扭角羚的种群数量是否达到了饱和状态,即是否存在种群密度过高的情况。
野生动物和生态系统的管理离不开对栖息地质量的评估,而栖息地内野生动物的容纳量往往是评价野生食草动物栖息地质量的依据。因此,不论是从物种保护的角度还是从栖息地保护的角度判断,环境容纳量的评估都是至关重要的。容纳量是野生动物管理学中的一个重要概念,其定义为“在生态系统不发生退化、动物体况保持良好的前提下,给定的生态系统在一年中环境最恶劣季节能够维持的最大种群数量”(Dasmann,1964)。确定某种野生动物的容纳量涉及到目标动物种群生态学特性和栖息地特征,需要充分了解动物的采食、消化、能量需求、空间、移动能力等方面的知识(Gates&Hudsong,1978)。
对唐家河扭角羚环境容纳量的研究是种群管理和栖息地管理的基础,也是自然保护区管理能力提升的重要途径。为了实现这个目标,自2012年冬季起,历时两年,笔者与唐家河自然保护区共同完成了扭角羚环境容纳量的外业工作,并于2014年11月完成内业分析。在此,我们对唐家河扭角羚环境容纳量调查进行总结。
1 研究内容和方法
1.1 研究区域和样地选择
唐家河国家级自然保护区(以下简称唐家河,东经 104°36'~104°56',北纬 32°32'~32°41')位于四川省广元市青川县,毗邻甘肃文县和四川平武县,是位于岷山山脉摩天岭南麓的大熊猫(Ailuropoda melanoleuca)保护区。唐家河属于低山丘陵向高山峡谷过渡的地带,最低海拔1 100 m,最高海拔3 864 m。植被类型随海拔梯度变化明显,从低海拔至高海拔依次是河谷灌丛及灌木林,落叶阔叶林,针阔混交林,针叶林以及高山草甸。区内主要分布有3种箭竹,分别是缺苞箭竹(Fargesia denudata)、糙花箭竹(Fargesia scabrida)和青川箭竹(Fargesia rufa),主要集中分布于1 800 m~2 600 m的林下。区内属于温带气候,根据保护区内气象记录(海拔1 700 m),最低气温发生在1月份(-11°C)。
唐家河自然保护区管辖面积约400 km2,由于面积较大,不能做到全面取样,因此调查样地的选择是按照采用分层抽样法的方式进行。具体步骤包括:(1)按照扭角羚冬季栖息地的海拔范围,确定越冬地总面积;(2)将研究海拔段划分成100 m×100 m的栅格后按照每种植被型所占的栅格比例,以1%的强度进行抽样,作为野外调查样地。(对于部分植被型在所选择海拔段内不足100个栅格,我们则抽取这种类型植被的1个栅格)。
一般来说,在任何有限的空间内,如果为动物提供的食物、水源和隐蔽地资源中的某一项成为短板,也就成为决定种群最容易达到饱和的主要因素。例如,水源是荒漠生境的短板、洞穴是穴居动物的短板,食物则是大型草食动物的短板(尤其在灾难性的气候中,如干旱、降雪等),这些因素均可成为制约种群数量的因素。因此,对扭角羚而言,冬季是食物条件和气候条件最恶劣的时期,成为制约扭角羚种群的短板。据此,本研究野外调查时间设定在冬季(12月至次年2月),结合2006年~2009年实施的扭角羚迁移特征研究(官天培,2012),将保护区境内扭角羚冬季适宜海拔段定为2 000 m~2 500 m,总面积约8 230 hm2。按照分层抽样原则,我们将保护区划分为面积为100 m×100 m的样地。因此在适宜海拔段内共有8 230个样地。按1%的抽样强度,本研究共需在82样地内进行大样方和小样方的调查。各植被型中,样地数量如下:桦木-铁杉45个样地,栎类-青冈林12个样地,柳树林两个样地,槭树-杨树林两个样地,山核桃-盐肤木12个样地,云杉-冷杉林9个。抽样过程中所涉及的地理信息处理均在ArcGIS 9.2中完成(图1)。
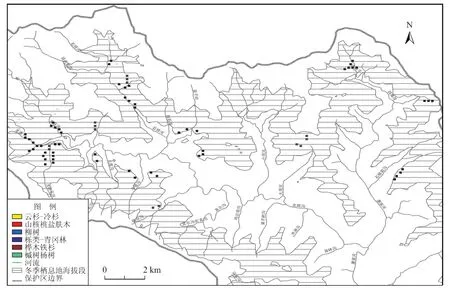
图1 唐家河扭角羚容纳量植物样方调查示意
1.2 样方调查及样品处理方法
在随机选择的调查样地中,我们分别设置4 m×4 m和1 m×1 m的样方各3个。在进行4 m×4 m大样方的调查时,我们需要记录灌木和草本的种类。基于已知扭角羚的采食习性(Schaller et al.1986),具体的野外操作流程是:用枝剪剪取灌木当年新生的、距离地面高度小于2.5 m的枝条;将所有枝条充分混合后称取200 g~400 g,用牛皮纸A4信封保存并标注样方序号,备用于进一步烘干处理;收割1 m×1 m小样方内所有的地表草本、并称重。把获得的草本样本充分混合,抽取200 g用牛皮纸A4信封保存并标注样方序号,备用于进一步烘干处理。
1.3 圈养扭角羚冬季能量需求调查
在动物园的协助下,从2013年1月10日至1月18日对成都动物园内圈养扭角羚进行了冬季营养需求的外业数据收集。该园扭角羚共7只(雄性4只、雌性两只、亚成体1只)圈养于面积约80 m2的围栏中。所有圈养个体在白天进行混合饲养,在夜晚分圈饲养。我们要求动物园饲养员在维持正常的投喂习惯的同时,在每次投料前进行饲料的称重、记录。晚上待扭角羚不再采食后,将剩余的食物移出并称重、记录。在连续喂养3 d、待动物适应新的环境后,我们用上述相同的方法连续收集5 d的数据。在投喂前,将草料剪成10 cm的草段,充分混合后,取其中200 g按日装信封保存、标记,烘干后测定粗蛋白、粗脂肪、粗纤维、灰份含量。
1.4 扭角羚现有种群数量估计
扭角羚仅在夏季繁殖期集成大群,是唯一可以高效进行种群数量调查的季节。笔者分别在2013年夏季和2014年夏季对大草堂区域的扭角羚数量进行计算(n),基于大草堂区域扭角羚密度(d),推算保护区范围内高山草甸夏季种群数量(N)。具体计算公式如下:
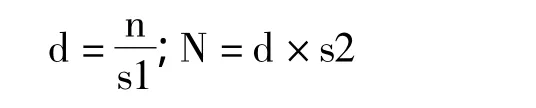
s1=大草堂高山草甸面积;s2=保护区内高山草甸面积
1.5 能量分析和容纳量估计
所调查的82个样地中,共计263份植物样品在收集后均放于烘干箱内在80℃下恒温干燥4h,其间不时进行翻转,确保均匀受热。在样品完全干燥后,我们使用专用粉碎机将样品研磨成粉末状,并滤去较大的颗粒。最后,在称重后将每个样品使用自封袋密封保存,送往实验室进行营养成分分析。营养成分分析是使用近红外漫反射光谱法进行测定的,该部分工作由英国爱丁堡动物园营养分析师 Andy Bear完成。所有营养分析在2014年10月完成。
能量估计是基于粗纤维、粗蛋白每克16.74(KJ)和粗脂肪每克37.66(KJ)的标准计算的(宋延龄等2003)。容纳量估计是在获取每种植被类型单位面积能量值与单只扭角羚冬季能量消耗总值之后进行的。由于圈养个体无法获取各个年龄段冬季能量消耗数值,仅能获得均值。因此,本研究尝试使用了两种方法进行容纳量估算:(1)依据圈养秦岭各年龄段及性别的扭角羚能量消耗标准并结合唐家河扭角羚种群结构,估算可容纳种群总数;(2)仅依据成都动物园能量分析结果,不分年龄段和性别估算可容纳成年个体总数。两种方法估算时都是以扭角羚消耗20%可利用食物的安全标准进行的(宋延龄等2003)。基于2006年~2009年的研究,发现扭角羚冬季栖息地地形较缓和极少活动在坡度40°以上的较陡峭生境,因此将坡度超过40°的区域排除在扭角羚冬季适宜栖息地的范围外。
2 结果
2.1 各植被类型冬季提供的能量值
保护区内冬季扭角羚种群主要分布的植被类型有6种,分别是桦木-铁杉林、栎类-青冈林、柳树林、槭树-杨树林、山核桃-盐肤木林、云杉-冷杉林,适宜扭角羚生存的冬季海拔段总面积8 230 hm2,以上 6种植被所占比例依次是 55.96%、14.79%、1.59%、1.11%、15.18%、11.13%。适宜冬季生境总面积约5 397 hm2,以上6种植被所占比例依 次是 51.55%、13.24%、1.56%、0.82%、22.24%和10.59%。冬季适宜海拔段内6种植被可提供总能量4.49±2.14×010(KJ)。冬季适宜栖息地中6种植被可提供的总能量值是2.82±1.31×1010(KJ)。
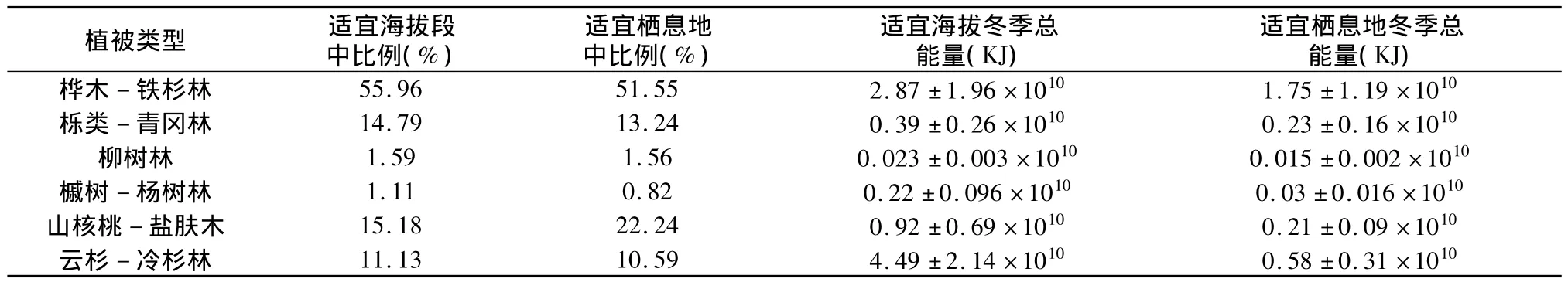
表1 唐家河自然保护区各植被类型能量
2.2 动物园圈养扭角羚冬季能量消耗
依照成都动物园的饲喂方式:白天扭角羚混合放养,仅饲喂新鲜草料和干草;夜间分圈喂养,主要投喂苜蓿颗粒和动物园专门定制的颗粒饲料,计算连续5 d投喂食物与剩余食物,得到每只成年扭角羚单日需消耗2 780 g草料(群体喂养)、3 780 g±980 g苜蓿颗粒饲料、1 800 g±360 g精饲料的结果。经能量换算,冬季每只扭角羚单日需摄入60 338(KJ)的能量。我们将每年的11月15日至次年3月15日定为唐家河的冬季,共计120 d,即冬季每只成年扭角羚需要总能量为7.24×106(KJ)。
2.3 环境容纳量
2.3.1 基于群结构的环境容纳量估计
经2007年~2009年野外观察,唐家河扭角羚种群由雄性 22.55%、雌性 39.25%、亚成体29.73%、幼体8.45%构成。计算过程如下:
设唐家河可容纳扭角羚数量为x,那么计算容纳量的公式为:
x=(0.2255×5.48+0.3925×3.62+0.297×2.9+0.0845×2.12)×104×120天 × E(保护区可提供总能量)×20%;依适宜生境计算,x1=1 232±506只;依适宜海拔计算,的x2=2 025±967只。
2.3.2 基于动物园圈养个体的环境容纳量估计
已知动物园圈养个体冬季成体需要7.24×106(KJ)的能量。唐家河冬季适宜海拔段可提供能量4.49±2.14×1010(KJ),可容纳成年扭角羚x3=1238±591只。唐家河冬季适宜生境可提供能量2.82±1.31×1010(KJ),可容纳成年扭角羚x4=778±362只。
2.4 现存扭角羚数量估计
调查显示,大草堂区域单日不重复计数情况下,最大种群数量102只。已知大草堂计数区域面积约41 hm2,可得夏季高山草甸扭角羚密度约2.48只·hm-2。保护区内高山草甸总面积月534 hm2,可得夏季高山草甸扭角羚数量约1 324只。
3 结论
3.1 环境容纳量
本研究采用两种标准进行容纳量的估算,分别得出适宜生境和适宜海拔段两种情况下可容纳扭角羚数量。虽然,目前仍无研究将地形因素做为环境容纳量计算因素,但环境容纳量是在动物自由选择适宜栖息地的情况下可承载的数量。此外,在适宜海拔段中的适宜地形中,还同域分布着斑羚、鬣羚等其他食草动物以及豹等天敌,真正可被称为适宜栖息地的面积会进一步降低、且可提供能量也因其它草食动物的消耗而相应减少。大量研究证明,过量的食草动物将改变森林生态系统的结构和功能,进而影响物种多样性(Reimoser&Gossow 1996)。因此,将扭角羚种群数量保持在适度的范围是非常必要的。在陕西佛坪保护区内秦岭扭角羚的容纳量的研究中,研究者也将一定的海拔范围作为扭角羚越冬栖息地的面积(宋延龄等,2003)。综合多种因素,笔者认为,将保护区适宜海拔段作为计算可容纳扭角羚数量做为种群管理的依据,具有合理性。将保护区内扭角羚的种群数量控制在约1 500只,即5.4只·km-2,能够保证扭角羚种群的生存安全。
3.2 种群数量
本调查显示夏季栖息于高山草甸扭角羚数量约1324只,表面上种群已经接近保护区的容纳量。但是,由于与唐家河自然保护区毗邻的自然保护区还包括北面的白水江国家自然保护区、东面的东阳沟省级自然保护区和南面的老河沟自然保护区,而这些保护区均有扭角羚分布,且高山草甸均分布在保护区边界区域。因此,无法确定夏季高山草甸扭角羚数量有多少是来自于唐家河自然保护区。但是,笔者认为调查结果所显示的高山草甸扭角羚数量不能够代表唐家河自然保护区目前的种群数量水平。因此,结合环境容纳量结果,笔者推断唐家河扭角羚数量尚未达到最大可容纳数量。为了更加科学管理和保护唐家河的扭角羚种群,准确高效的数量调查和长期开展监测夏季高山草甸区域扭角羚的来源非常有必要。
容纳量是野生动物管理学所涉及的基本问题。影响容纳量的因子非常多,如栖息地的初级生产力决定了可利用植物的生长量,而植物的生长量又受到当年的气温和降水量的影响,可利用植物的营养成分和含量则受到栖息地土壤的肥力的控制(Hobbs et al.,1985)、植物的可利用又与冬季积雪厚度及其对动物移动能力的限制有密切关系(Hobbs et al.,1982)。所以,以往研究对这个问题的议论远多于实例的报道。尽管,唐家河保护区多年对扭角羚种群的监测为本项研究提供了一定的基础,但是保护区现有的资料还不能提供保护区内扭角羚栖息地特征的细节。基于越冬栖息地能够提供的可利用食物资源总量得出的容纳量数值,只能是一个粗略的估计值,存在高估或低估的可能。更准确的容纳量数据,还需要更深入细致的研究。
致谢
扭角羚环境容纳量调查历时两年,多部门和单位为该项目的实施给予的大力的支持,包括唐家河保护区管理处、各保护站、保护科和科研科、中国科学院动物研究所、绵阳师范学院生态安全与保护四川省重点实验室、成都动物园、英国爱丁堡皇家动物学会等,在此表示谢忱。由于本项目的野外调查主要在寒冷的冬季和炎热夏季,保护区员工进行大量艰苦的野外调查,对项目的完成做出了非常重要的工作,在此表示感谢!同时也感谢绵阳师范学院的研究生和本科生对大量样品的预处理中的努力。最后,感谢英国皇家动物学会和Andy beer博士对大量植物进行的营养分析。
[1]官天培.扭角羚生态学研究-分布、迁移、家域及行为学研究[D].北京师范大学,2012.
[2]胡锦矗,王酉之.四川资源动物调查(第二卷兽类)[M].成都:四川科学技术出版社,1984.
[3]胡锦矗,胥晓,张君.四川唐家河自然保护区综合科学考察报告[M].成都:四川科学技术出版社,2005.
[4]胡锦矗.哺乳动物学[M].深圳:中国教育文化出版社,2007.
[5]宋延龄,党高第,李俊生,等,赵雷刚.陕西佛坪国家级自然保护区扭角羚容纳量[J].动物学报,2003,49:445~450.
[6]宋延龄,曾治高.秦岭扭角羚的生存现状及其面临的问题[J].生物多样性(香港),2001,2:94~100.
[7]孙儒泳.动物生态学原理[M].北京:北京师范大学出版社,2001.
[8]汪松,解焱.中国物种红色名录[M].北京:高等教育出版社,2009.
[9]王应祥.中国哺乳动物种和亚种分类名录与分布大全[M].北京:中国林业出版社,2003.
[10]吴家炎.中国扭角羚[M].北京:中国林业出版社,1990.
[11]虎绍勇,谌利民,杨建东.唐家河自然保护区扭角羚死亡监测探讨[J].四川动物,2011,30(6):982~985.
[12]Dasmann R F.Wildlife Biology[M].Newyork:John wiley& Sons Inc,1964.
[13]Gates C,Hudson R J.Energy cost of locomotion in Wapiti[J].Acta Theriologica,1978,23,365~370.
[14]Guan T P,Wang F,William J M,et al.Nature reserve requirements for landscape-dependent ungulates:the case of endangered takin(Budorcas taxicolor)in Southwestern China[J].Biological Conservation,2015,182:63~71.
[15]Hobbs N T,Baker D L,Ellis J E,et al.Energy and nitrogenbased estimates of elk winter range carrying capacity[J].Journal of Wildlife Management,1982,46(1):12~21.
[16]Hobbs NT,Swift DM.Estimates of habitat carrying capacity incorporating explicit nutritional constraints[J].The Journal of wildlife management,1985:814~822.
[17]Reimoser F,Gossow H.Impact of ungulates on forest vegetation and its dependence on the silvicultural system[J].Forest ecology and Management,1996,88(1):107~119.
[18]Schaller G B,Deng Q T,Pan W S,et al.Feeding behavior of Sichuan takin(Budorcas taxicolor)[J].Mammalia,1986,50:311~322.
