甘肃白龙江干旱河谷不同海拔梯度植物α分析
2015-12-02陈国鹏黄旭东
王 飞,郭 星,陈国鹏,黄旭东
(甘肃省白龙江林业管理局林业科学研究所,甘肃兰州 730070)
干热河谷在世界统一划分的3大类干旱地区中,既不属于大陆中心荒漠,也不是副热带稀树草原,而是属于局部的干旱生境。干热河谷的出现和分布具有明显的自然背景,标示着自然地带性环境的特殊性。中国的干旱河谷区分布在嘉陵江的支流白龙江流域、岷 江上游、金沙江上游、大渡河上游、雅碧江上游等地区[1]。
白龙江流域干旱河谷是长江水系北部的生态脆弱带和敏感区,其植被生态系统严重退化,泥石流和滑坡等地质灾害频发,水土流失严重,治理困难,具有向荒漠化发展趋势。干旱河谷也是白龙江流域人口和城镇分布密集的地带,其恶劣的生态环境和逐渐增多的自然灾害严重影响当地及下游的工农业生产和人民群众的生活,已成为制约该地区经济社会可持续发展的主要因素[2-3]。由于白龙江中下游地区地质构造较复杂,地形、地貌的差异引起了水、热的再分配,形成了不同的生物、气候带的自然景观,对土壤形成和发展、植被类型的演替也起到了支配作用。现存植被均为旱生灌丛、草丛,层次结构单一,生态景观特征呈现为旱生半荒漠化景观。气候干燥,降水少而不均,蒸发量极大[4-6]。针对白龙江干旱河谷地带的自然特点,研究不同海拔植物生物α多样性,以期为白龙江干旱河谷生态治理提供数据支撑。
1 试验材料与方法
1.1 自然概况
白龙江发源于甘、青、川3省交界处的郎木寺附近,南以岷山与四川分界,西接积石山高原,北以迭山与黄河水系一级支流洮河分水,向东南流至四川昭化汇入嘉陵江,全长约600 km,为长江水系北部的主要二级支流。流域整体位于青藏高原东北边缘,白龙江流经的地域广、落差大,河道全长576 km,其位于东经 102°46′~ 104°52′,北纬 33°04′~35°09′,属典型的西南高山地形,温度、降水具有明显的垂直梯度和水平差异[7]。由于坡陡谷深、降水量偏少、峡谷地貌对太阳辐射的反复折射和地面强烈辐射造成的增温作用及焚风效应,加之樵采、垦荒、放牧、筑路等人为干扰的强度和频度相对较大,在沿江海拔2 100 m(迭部县尼傲乡)往下至760 m(文县口头坝乡)和主要支流(阿夏沟、岷江、拱坝河、北峪河、羊汤河、白水江)两岸的河谷及浅山地带,形成以气候干暖少雨、植被稀疏残败、地形破碎、土地石漠化及岩漠化为主要特征的干旱河谷景观,泛称为干热河谷。植被主要由旱中生小叶落叶具刺灌木及耐旱草本(禾草为主)植物组成.根据气候要素并参考植被特征,白龙江干旱河谷主要为干暖河谷类型,局部为干温河谷类型[8-9]。白龙江干旱河谷地区的土壤特征表现为土层浅薄、土壤结构差、土壤石质、粗骨性强或表土紧实、土壤干旱等,土壤质地比较差。土壤水分性质是植物生长和恢复的关键制约因素,土壤干旱是土壤水分性质恶化的结果,土壤水分特性是影响该区植被恢复的重要因素[10-12]。
1.2 研究方法
1.2.1 外业调查
取样于白龙江干旱河谷地区样地海拔793 m~2 106 m之间,每100 m为一海拔阶梯,每一阶梯选择代表性地段布设3个~5个样方,共计73个样方,每个样方地点的选择兼顾不同的群落类型,所选取的样方具有代表性,能反映白龙江干早河谷植被的主要类型,生境条件等。
植被调查:(1)群落灌木层设置5 m×5 m样方,记录样方内植物名称、经纬度、海拔高度、坡度、坡向等因子,测量每一物种高度、基径、株丛数、多度、盖度。
(2)草本层取样,在灌木样方四角和中央各取一个1 m×1 m草本样方,记录样方内植物名称,测量每一物种高度、基径、株丛数、多度、盖度等因子。
1.2.2 数据计算
数据统计采用Excel软件、SPPS18.0软件等。采用以下指数对物种多样性进行测定:生物多样性指数H(Shannon-Wiener指数),H=-∑(PilnPi);优势度指数SP(Simpson指数),SP=1-(∑pi2),均匀度指数Sw(Pielou指数),Sw=(-PilnPi)/lnS,式中:S为样地内的物种数;丰富度指数D(Margalef指数),D=(S-1)/LnN,N为观察到的个体数总数,S为群落中的总数目;Pi为物种的相对重要值[13]。
2 结果与分析
2.1 不同海拔梯度上植物种群组成和分布的动态变化
通过实地调查统计,将白龙江干旱河谷地带按不同海拔分成13个不同的小组。
植物的海拔梯度被认为是影响物种多样性格局的重要因素之一。与纬度梯度相似,作为地理梯度的海拔梯度由于包含了温度、湿度和光照等诸多环境因子而成为生物多样性梯度格局研究的重要方面[14]。并且环境因子沿海拔梯度的变化要比沿纬度梯度快1 000倍[15],因而成为生态学家的重要研究对象。白龙江干热河谷灌木和草本植物种数随海拔变化如图1所示。不论灌木还是草本植物种数都随海拔的升高表现出先略升高然后降低最后又升高的趋势,在海拔1 850 m左右灌木的种数最少(14种),在海拔2 100 m左右则最多(36种);而草本植物种数在海拔1650m左右最少(2种),在海拔1 350 m左右最多(25种);不管海拔怎么变化,同一海拔处灌木的种数总是大于草本植物的种数(参见图1)。

图1 不同海拔植物种数Fig.1 Plant species at different altitude
2.2 相同海拔梯度上坡面及坡向植物种群生物多样性分析
坡面及坡向不同位置上的α多样性分析如图2所示。坡向是山地景观的重要特征因素,阳坡和阴坡在温度和水分条件方面的差异,往往造成不同坡向上植物群落的类型和特征差别明显,成为限制植物生长的重要因素[10]。不同坡向的群落多样性,从图2中可以看出,灌木层的Shannon指数大小为:阴坡>半阴半阳坡>阳坡,而草本层为:半阴半阳坡>阴坡>阳坡。无论是灌木层还是草本层,多样性指数阳坡均最低。这是由于阴坡的土壤水分含量高于阳坡,阳坡较阴坡有温度高、湿度小、蒸发量大、土壤的物理风化和化学风化都强的特点,因而土壤有机质积累少,也较干燥和贫膺,植被发育较差,这在土壤水分是植被生长状况的主要限制因素的干早河谷来说,表现的更明显。

图2 不同坡形坡面的Shannon指数Fig.2 Slope surface Shannon index on different slopes
从不同坡形上的多样性分析无论是灌木层还是草本层,多样性大小为:凹坡>平坡>凸坡。从地形方面看,凹面不同于平面和凸面,凹面是凸面和平面养分水分流失的聚集点,养分经过淋溶集中在凹面的位置,因此凹面有较高的土壤养分和水分,支持着较高的多样性。
2.3 海拔梯度上物种α多样性分析
通过对白龙江干旱河谷不同海拔的α多样性分析,灌木层的平均Shannon-wiener指数达到2.85,在海拔1 350 m处达到峰值,为3.26;在海拔1 850 m处最小为1.91。草本层的平均Shannon-wiener指数为1.85,在海拔1 450 m处最高,为2.68。在海拔850 m处达到最小值,为1.19。无论是草本还是灌木群落,随着海拔的增加,Shannon-wiener指数都呈现出先增加后减小的趋势,灌木层的α多样性明显高于草本层的α多样性,但是变化情况没有
草本层明显,但总体上灌木层和草本层有着非常相似的变化趋势。说明在海拔1 350 m和海拔1 450 m处灌木层和草本层所含的物种信息量是最大的,复杂程度最大。不管是灌木层还是草本层所含的物种信息量都是随着海拔的增加先增加后减小,且灌木层的物种信息量不管在哪个海拔都大于草本层信息量。Shannon-wiener指数与海拔的关系如图3所示。

图3 Shannon-Wiener指数随海拔高度的变化Fig.3 Changes of Shannon-Wiener indexes along with altitude
对于灌木层和草本层的Pielou指数在海拔1 850 m和1 650 m达到最大值为0.115和0.251;在海拔1 450 m处都达到最小值0.032和0.015;平均值为0.05和0.11。灌木层和草本层的均匀度指数随海拔的变幅较小,与草本层相比,灌木层的均匀度较小,变化不明显,基本保持在0.05左右,但草本层的Pielou指数总体大于灌木层,且变化明显。说明在白龙江干旱河谷地带草本层物种分布比灌木层的物种分布均匀,在海拔1 850 m和1 650 m达到最大值且在这两个海拔带处灌木和草本的分布是最均匀的,相反在海拔1450m处的分布是最不不均匀的,总体上看灌木的物种分布变化比草本大。Pielou指数与海拔的关系如图4所示。
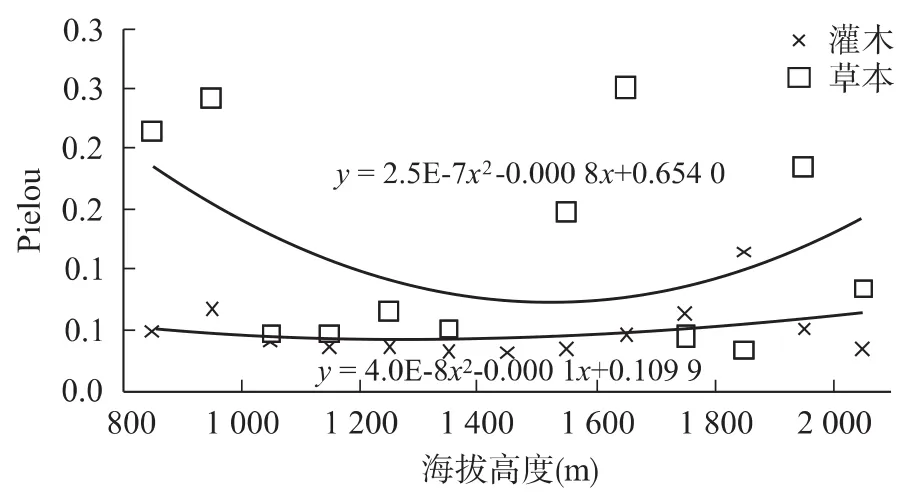
图4 Pielou指数随海拔高度的变化Fig.4 Changes of Pielou indexes along with altitude
灌木层和草本层的Simpson指数随海拔的变幅较小都保持在1附近。灌木层优势度指数在海拔1 850 m处达到最大值为0.98,在海拔950 m处为最小0.91,灌木层优势度指数平均为0.95;草本层在海拔1 350 m处优势度指数最大为0.98,在海拔1 650 m处最小为0.33,草本层的平均优势度为0.73.与灌木层相比,草本的优势度指数小于灌木层指数,但是变化趋势比灌木层明显。说明白龙江干旱河谷的灌木层在各个海拔梯度上都有较明显的优势物种,在海拔1 850 m处达到最大,在海拔950处为最小;草本层的这种优势不太明显,而且随海拔的增加优势物种的变化比较大,在海拔1 350 m处达到最大,在海拔1 650 m处达到最小,也是先升高后降低。Simpson指数与海拔的关系如图5所示。

图5 Simpson指数随海拔高度的变化Fig.5 Changes of Simpson indexes along with altitude
灌木层的Margalef指数随海拔的变化在海拔1 650 m处达到最大值为4.62,在海拔1 850 m处为最小1.12,灌木层优势度指数平均为3.45;草本层在海拔1 350 m处优势度指数最大为3.85,在海拔1 650 m处最小为0.31,草本层的平均丰富度为1.74。不管是灌木群落还是草本群落,变化趋势一样都是随着海拔的升高先升高后又降低,但是总体灌木层的丰富度指数大于草本层,变化都比较明显。说明不管是灌木层还是草本层物种丰富度都是随海拔的升高先增加后减小,且在海拔1 650 m和1 350 m处达到最大值及在这两个海拔处物种是最丰富的,在海拔1 850 m和1 650 m处最小及在这两个海拔处的物种是最少的。Margalef指数与海拔的关系如图6所示。

图6 Margalef指数随海拔高度的变化Fig.6 Changes of Margalef indexes along with altitude
3 结论与讨论
植物种、植物群落的分布格局是不同尺度上,各种环境因子,如气候、土壤、地形等综合作用的结果。区域尺度上讲,气候、植物区系决定特定的植被类型;景观尺度上讲,如海拔这一环境因子,使植被沿局部气候条件的变化而导致植被类型的变化,进而决定植被分布格局[13],而对植物的分布和生长起重要作用的微生境、土壤养分等也由这些主导因子所控制;本文所做的生物多样性分析,揭示了白龙江干旱河谷地区植被与海拔、坡面、坡向的关系。
由于白龙江地区环境条件恶劣,特别是干旱缺水、过度放牧、人为破坏等因素的影响,决定了干旱河谷地区植被性质[8,9],表现出以灌木为本的生态系统,但随海拔的升高灌木和草本的物种数略显升高后又降低,最后升高,在海拔1 800 m达到最低,这一区域物种最少,植被破坏严重,气候条件差,生态系统更为脆弱。在这一区域更需要采取一定的措施恢复植被。
通过α多样性分析的;物种多样性和丰富度都随海拔的升高略增加后又降低,最后升高的趋势,说明白龙江干旱河谷地区的生态系统的复杂程度和所含信息量也是略增加后又降低,最后升高,在海拔1 850 m左右达到最小;物种优势度和均匀度变化都不大;不论海拔怎么变化灌木始终是白龙江干旱河谷地带的主要优势种。不同坡面位置上的多样性分析表明,无论是灌木层还是草本层,多样性指数阳坡均最低,阴坡和半阴半阳坡有着较高的多样性。无论是灌木还是草本,凹坡和平坡的多样性比凸坡高,结果与许多干热河谷的研究相似[4,10]。
通过研究白龙江干旱河谷地区不同海拔植物得出中海拔地区生物多样性低,低海拔和较高海拔生物多样性较高。
[1]赵琳,郎南军,郑科,等.云南干热河谷生态环境特性研究[J].林业调查规划,2006,31(3):114~11.
[2]中国科学院青藏高原综合科学考察队.横断山区干旱河谷[M].北京:科学出版社,1992.
[3]冯自诚,刘刚,刘谦和.白龙江中上游森林生长与立地条件的相关分析[J].甘肃农业大学学报,1993,28,317~324.
[4]刘醒华.岷江上游干旱河谷的形成条件与植被恢复的探讨[M]//宋达泉.森林与土壤(第三次全国森林土壤学术讨论会论文选编).北京:中国林业出版社,1985:166~180.
[5]岷江上游综合考察队.岷江上游森林生态问题综合考察报告[J].四川林业科技,1980(增刊):1~31.
[6]张荣祖.横断山区干旱河谷[M].北京:科学出版社,1992.
[7]邱祖青,杨永宏.白龙江干旱河谷木本植物多样性及其区系地理特征[J].甘肃农业大学学报2007,10:119~125.
[8]中国科学院青藏高原综合科学考察队.横断山区干旱河谷[M].北京:科学出版社,1992.
[9]杨兆平,常禹,布仁仓,等.岷江上游干旱河谷区域空间变化的定量判定[J].生态学报,2007,27(8):3250~3256.
[10]孙学刚,冯自诚,张承维.白龙江中上游森林植物区系研究[J].甘肃农业大学学报,1993,28(专辑):65~78.
[11]康永祥.白龙江流域木本植物区系特征[J].西北植物学报,1999,19(2):337~343.
[12]丁磊,孙学刚,刘晓娟,等.白龙江中上游林区种子植物物种多样性与区系特征[J].甘肃农业大学学报,2006,41(6):79~85.
[13]方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[M].生物多样性,2009,17(6):533~548.
[14]刘世荣.中国温暖带森林生物多样性研究[M].北京:中国林业出版社,1998.
[15]YODA K A.Preliminary survey of the forest vegetation of eastern-Nepal[J].Journal of College Art and Sciences,1997,2:153~162.
