皱肋文蛤胚胎、幼虫及稚贝的发育
2015-12-02栗志民钱佳慧刘志刚刘锦上李艳红
栗志民, 钱佳慧, 刘志刚, 刘锦上, 李艳红
(广东海洋大学 水产学院, 广东 湛江 524025)
皱肋文蛤(Meretrix lyrata), 又名越南文蛤、琴文蛤,隶属软体动物门(Mollusca), 瓣鳃纲(Lamellibranchia),异齿亚纲(Heterodonta), 帘蛤目(Veneroida), 帘蛤科(Veneridae), 文蛤属(Meretrix)。主要分布在中国台湾、广西和广东, 以及越南和菲律宾[1], 是潮间带中低潮区典型的埋栖型双壳经济贝类。20世纪90年代后期中国从越南引进天然种苗进行养殖后, 在雷州半岛沿海各地出现了小规模的自然种群。由于该贝适应性强、养殖周期短、产量高、肉质鲜美, 因此深受国内外市场欢迎, 市场需求越来越大, 价格一路攀升, 依靠进口天然种苗养殖已经不能满足市场需要。近年来, 广东海洋大学攻克种苗规模化生产技术难关, 已能够生产出大量种苗供应市场, 在雷州半岛沿海初步实现了皱肋文蛤的规模化育苗。
国外学者先后研究了皱肋文蛤对微量金属元素的生物富集和净化作用[2]; 评估了其营养成分[3]; 调查了其在越南海域生存的环境因子、养殖生物学及群体动态分布模式[4-6]; 分析了自然条件与皱肋文蛤栖息场的形成和发展之间的关系[7]。国内学者在皱肋文蛤的人工育苗[8]、养殖技术[9]、摄食生理[10]、呼吸生理[11], 以及对温、盐度的适应性[12]等方面开展了研究, 但有关皱肋文蛤的早期胚胎发育过程, 以及幼虫和稚贝的形态学特征和行为方式, 尚未见报道。作者在皱肋文蛤人工育苗过程中, 详细观察了其早期发育的形态学特征和行为方式, 旨在为皱肋文蛤的人工育苗和养殖提供理论指导, 以及为该贝胚胎、幼虫和稚贝的质量评价提供基础研究资料。
1 材料和方法
1.1 材料
实验于2013年6~7月在湛江市遂溪县湛江银浪海洋生物技术有限公司水产种苗场进行。皱肋文蛤亲贝1~2龄, 采自湛江港北月村滩涂养殖区, 平均壳长(51.54±1.63)mm、平均壳高(44.03±1.34)mm、平均壳宽(22.61±1.15)mm、平均个体鲜质量(37.15±1.27)g。
1.2 方法
1.2.1 催产、孵化及幼虫培育
皱肋文蛤在每年的 6~7月份进入繁殖盛期, 选取性腺饱满的亲贝, 将亲贝置于室内催产池(长7.0 m×宽3.5 m×高0.9 m)中, 采用阴干结合流水刺激催产,一般30 min后雄贝开始排精, 紧接着雌贝产卵。卵子密度控制在4~5 个/mL, 催产、孵化和幼虫培育均在原池中进行, D形幼虫培育密度2个/mL。当幼虫平衡囊明显可见, 达到 30%时即倒池移入室外已铺好附着基的池中, 池顶加盖遮阳网。幼虫附着期密度0.5 个/mL, 附着期一般2~3 d, 从受精卵至附着完毕约需9 d。附着后36 d左右, 稚贝壳长达到0.8~1.0 mm时即可出池。培育期间用石英砂作为变态幼虫的附着基和稚贝的栖息底质。
孵化期间保持微充气, 水温 28.5~30℃, 盐度23.6~25.4, pH 8.1~8.2。幼虫期的管理参考栗志民等[8]方法。前3 d通过加水方式改善育苗池水质, 开口饵料选择湛江等边金藻(Isochrysis zhanjianggensis), 投饵密度 5000 个/mL。3 d后开始换水, 每次换水20%~30%, 保持水质新鲜, 换水网目随幼虫的生长做相应调整, 换水温差不超过2℃。每天7: 30和17: 00各投饵一次, 依幼虫的摄食情况调整投饵密度, 一般保持湛江等边金藻(1.5~2)×104个/mL, 牟氏角毛藻(0.4~0.8)×104个/ mL。每天测量水温、盐度, 以及幼虫的密度、壳长和壳高。
1.2.2 胚胎、幼虫及稚贝形态特征观察与测量
胚胎、幼虫和稚贝发育过程在光学显微镜下观察, 并用数码相机显微摄影。胚胎期用目微尺测量其长径和短径, 并记录发育期。幼虫及稚贝期测量其壳长和壳高, 并记录发育情况。
1.2.3 数据处理
实验数据采用平均值(M)±标准差(SD)表示, 用Excel软件作图。
2 结果
2.1 胚胎发育
皱肋文蛤成熟卵细胞呈圆球形, 胚泡已经破裂,直径(80±0.87)μm。受精后, 受精卵表面隆起极薄而透明的受精膜(图版Ⅰ-1), 卵周隙约 8~10μm, 受精膜在后续的胚胎发育中逐渐膨胀, 最终在多细胞期破裂。在水温28.5~30℃, 盐度23.6~25.4条件下, 受精卵经过20 min左右第一极体排出(图版Ⅰ-2), 经过30 min第二极体排出(图版Ⅰ-3)。受精后50 min左右,卵进行第一次不等分裂, 分裂线与卵子动植轴平行偏于一侧, 纵裂成 2 个大小不等的分裂球, 即为 2细胞期, 二者之间的卵裂沟十分明显(图版Ⅰ-4)。第二次不等卵裂开始于受精后 1h 10 min 左右, 分裂面与第一次垂直, 与卵轴方向平行, 但在较大分裂球上的分裂线稍偏一边, 形成一大三小的 4个卵裂球, 进入4细胞期(图版Ⅰ-5)。受精后1h 50 min, 进行第三次卵裂, 分裂面位于赤道附近, 与卵的动植轴垂直, 每个分裂球上半部向右旋转, 分裂后分裂球继续位移, 把上部4个分裂球移向顶端, 形成8细胞期(图版Ⅰ-6)。约2h 30 min, 行第四次分裂, 分裂球旋转方向与前一次相反, 即左旋分裂, 形成 16细胞期(图版Ⅰ-7)。随着细胞不断分裂, 分裂球数量不断增多, 而胚胎总体积并不增大, 分裂球体积越来越小, 受精4h后成为桑椹胚(图版Ⅰ-8)。桑椹胚进一步发育, 约4h 30 min, 大分裂球逐渐陷入胚体之内,小分裂球包围大分裂球, 形成一囊状的结构, 此期为囊胚期, 囊胚的表面布有纤毛, 开始孵化游动(图版Ⅰ-9)。由于外胚层小细胞下包, 以及大细胞内陷,受精后5h, 发育形成一个具有两胚层的原肠胚(图版Ⅰ-10)。受精后7h 30 min, 胚体渐变梨形, 顶端膨大,细胞加厚, 长出一圈纤毛环, 中央具有一丛纤毛, 为顶纤毛束, 早期幼虫喜浮游水表面和明晰透亮的地方, 具有较强的游泳能力, 此期为担轮幼虫期(图版Ⅰ-11)。皱肋文蛤早期胚胎发育时间和生长见表1。
2.2 幼虫及稚贝发育
根据幼虫和稚贝的规格、行为特征、足部发育状况、平衡囊及鳃丝出现时间、面盘退化时间、足丝分泌阶段、水管形成情况、壳色及其透明度等特征, 把皱肋文蛤幼虫发育阶段分为 D形幼虫、壳顶初期幼虫、壳顶中期幼虫、壳顶后期幼虫、变态期幼虫、附着期幼虫, 把稚贝阶段划分为无管期稚贝、单管期稚贝、双管期稚贝(表 2)。依显微观察, 幼虫和稚贝各个时期形态结构特征如下。
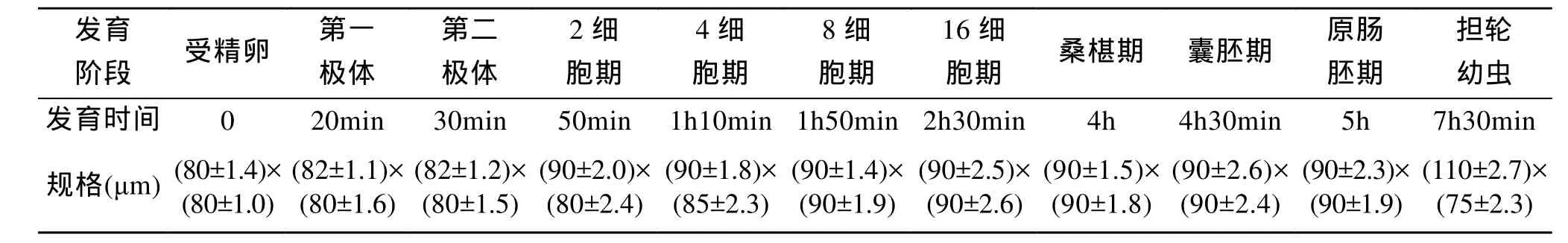
表1 皱肋文蛤早期胚胎发育时间和生长Tab.1 Developmental time and growth of early embryo of Meretrix lyrata
(1) 直线铰合幼虫(D 形幼虫): 受精后 16h, 胚体两侧覆盖 2片较硬而透明的半圆形的幼虫壳, 呈“D”形, 1 d 后, 幼虫壳长(110.8±1.9)μm, 壳高(87.7±2.5)μm。两壳在背部成直线铰合, 胚体顶端呈椭圆盘状, 形成运动器官面盘, 其四周细胞被有纤毛, 贝壳开壳依赖位于铰合线下方的前后闭壳肌, 消化器官分化尚未完善, 胃和消化盲囊不发达(图版Ⅰ-12)。
(2) 壳顶初期幼虫: 3 d时, 幼虫贝壳两侧靠近中央处稍稍隆起, 铰合线成微弧形, 形成微隆起的幼虫壳顶, 内脏团下方出现圆球状足原基。幼虫消化盲囊较发达, 对食物的消化吸收能力大大增强, 消化盲囊颜色变深, 呈黄褐色(图版Ⅰ-13)。
(3) 壳顶中期幼虫: 壳顶微隆起, 变化不显著,壳透明无色, 壳长较初期幼虫, 生长明显。足原基发育成靴状足, 足基部出现1对中空的细胞团, 称平衡囊, 中央包含1粒灰褐色耳石(图版Ⅰ-14)。
(4) 壳顶后期幼虫: 壳顶微隆起, 变化仍不显著。靴状足延长为棒状。平衡囊愈加显著, 清晰可见, 耳石颗粒增大。在足的基部两侧, 形成 1对成管状的内鳃丝。值得一提, 在皱肋文蛤幼虫发育过程中没有出现眼点, 幼虫成熟的判断依据为壳长及平衡囊(图版Ⅰ-15)。
(5) 变态期幼虫: 从 D 型幼虫到变态期幼虫约需6 d时间。此期幼虫面盘逐渐萎缩, 浮游能力逐渐尚失, 最终下沉至底部, 在底部一边依靠尚未脱落的面盘做间歇性游动, 一边利用足在底部做匍匐运动。变态期幼虫面盘上的纤毛逐渐脱离面盘, 最终使幼虫失去游动能力, 完全进入底栖生活, 完成从浮游向底栖的变态(图版Ⅰ-16)。
(6) 附着期幼虫: 壳顶隆起变圆, 变化十分明显。幼虫进入底栖生活时, 幼虫足部分泌黏状足丝附着在沙粒上, 面盘消失。内鳃丝逐渐增多, 开始用鳃呼吸, 靠鳃丝的摆动滤食水中食物颗粒。幼虫壳色与前面各期均为透明无色(图版Ⅰ-17)。
(7) 无管期稚贝: 贝壳外观卵圆形。稚贝的胚壳与次生壳的衔接处平滑, 无明显的交界, 两者透明度相同, 但壳色在次生壳部分为红褐色。从胚壳到次生壳, 壳面生长线明显且隆起。水管尚未形成, 靠进出水孔进行摄食与排泄(图版Ⅰ-18)。
(8) 单管期稚贝: 壳顶逐渐隆起变尖。在壳背后方长出出水管, 管口缺乏触手。此期稚贝肉眼明显可见, 外观呈红褐色, 育苗生产上把此期称为“红壳期”(图版Ⅰ-19)。
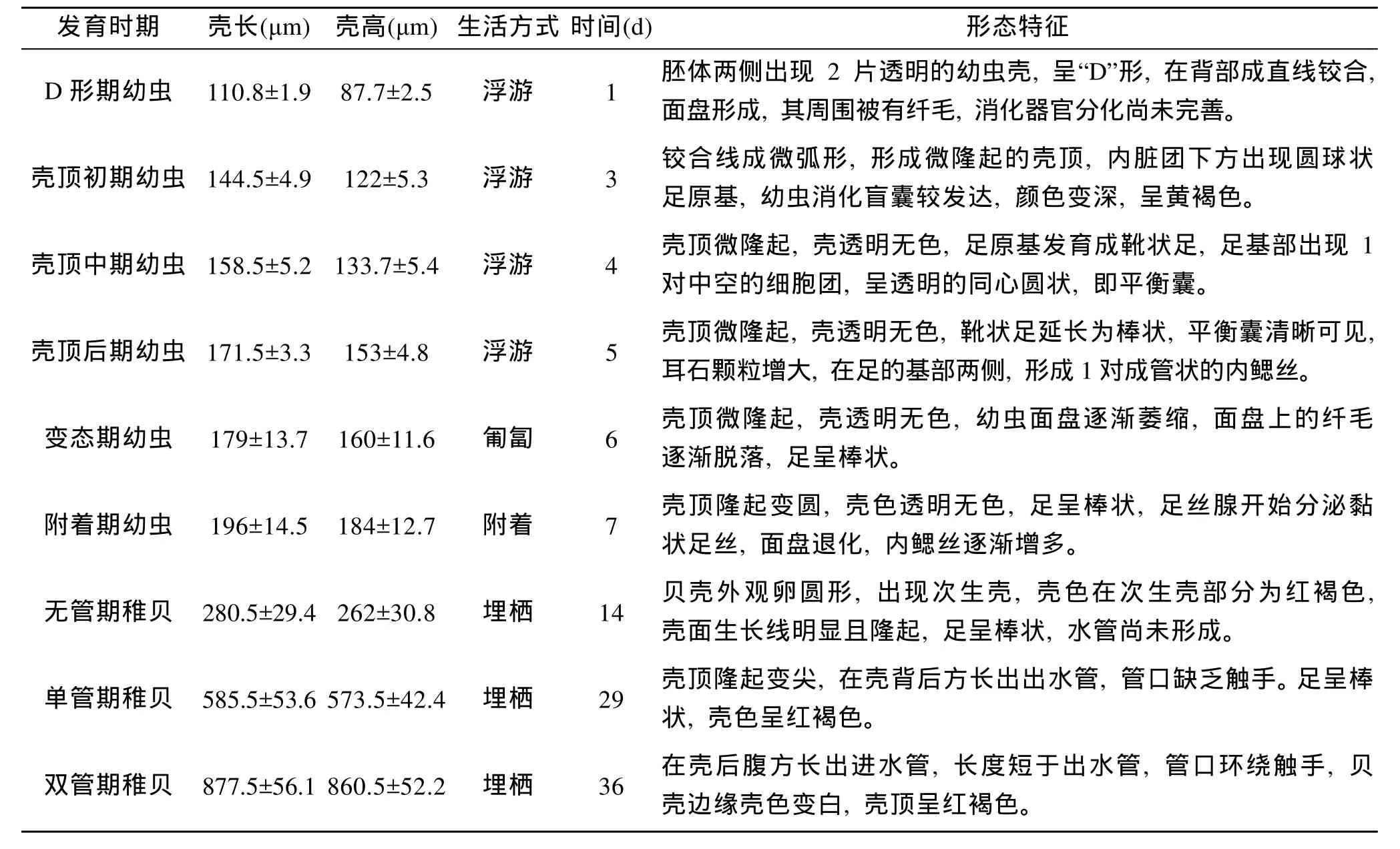
表2 皱肋文蛤幼虫和稚贝的形态特征和行为方式Tab.2 The morphological character and behavioral manner of larvae and spat of Meretrix lyrata
(9) 双管期稚贝: 稚贝在壳后腹方长出进水管,长度短于出水管, 管口环绕触手。此期稚贝贝壳边缘壳色变白而不透明, 育苗生产上把此期称为“白壳期”(图版Ⅰ-20)。此期稚贝各器官已发育完善, 外部形态与成体基本相同, 但壳顶仍呈红褐色, 不同于成体的壳顶。育苗生产早期培育结束, 可进入种苗中间培育阶段。
2.3 皱肋文蛤胚胎、幼虫和稚贝的生长
如表 1所示, 皱肋文蛤胚胎发育阶段从受精卵开始直至担轮幼虫, 该贝类受精卵大小为(80×80)μm,随着胚胎不断发育, 细胞数目迅速增多, 约 5h胚胎进入原肠期, 其胚胎大小为(90×90)μm, 受精后 7h 30 min, 胚体发育至担轮幼虫期, 其胚胎大小为(110×75)μm。如图 1所示, 从 D形幼虫期开始直至附着期幼虫, 该阶段幼虫体积出现较大幅度增长, D形幼虫规格(110.8×87.7)μm, 达附着期幼虫时, 规格为(196×184)μm。幼虫期壳形变化也较明显, D形幼虫期铰合部呈直线铰合, 附着期幼虫壳顶明显, 隆起变圆。而壳颜色仍保持透明无色。变态完成的稚贝进入埋期生活, 稚贝阶段包括无管期稚贝、单管期稚贝和双管期稚贝, 壳体积变化明显, 其中, 无管期稚贝(280.5×262)μm, 而双管期稚贝(877.5×860.5)μm。壳形由隆起变圆向隆起变尖过渡, 而且该阶段长出了明显的出、入水管。

图1 皱肋文蛤幼虫和稚贝的生长Fig.1 The growth of larvae and spat of Meretrix lyrata
如图 2所示, 皱肋文蛤从 D形幼虫到双管期稚贝的发育过程中, 壳长/壳高呈现先快速下降再缓慢下降的趋势。其中, 从D形幼虫到附着期幼虫, 壳长/壳高迅速下降, 而从附着期幼虫, 经无管期稚贝、单管期稚贝到双管期稚贝, 壳长/壳高呈缓慢下降趋势。
3 讨论
3.1 皱肋文蛤早期发育的分期
作者依据皱肋文蛤的早期发育状况, 将其早期发育分为胚胎发育阶段、幼虫发育阶段和稚贝发育阶段, 其中胚胎发育阶段从受精卵开始至担轮幼虫分8期, 即受精卵、第一极体、第二极体、卵裂期、桑椹期、囊胚期、原肠胚期、担轮幼虫期。依据幼虫的行为方式、壳顶形状、面盘和足将幼虫发育阶段分为 6期, 即 D形幼虫、壳顶初期幼虫、壳顶中期幼虫、壳顶后期幼虫、变态期幼虫和附着期幼虫。依据水管的有无和发育状况将稚贝发育阶段分为 3期, 即无管期稚贝、单管期稚贝和双管期稚贝。
3.2 皱肋文蛤胚胎发育
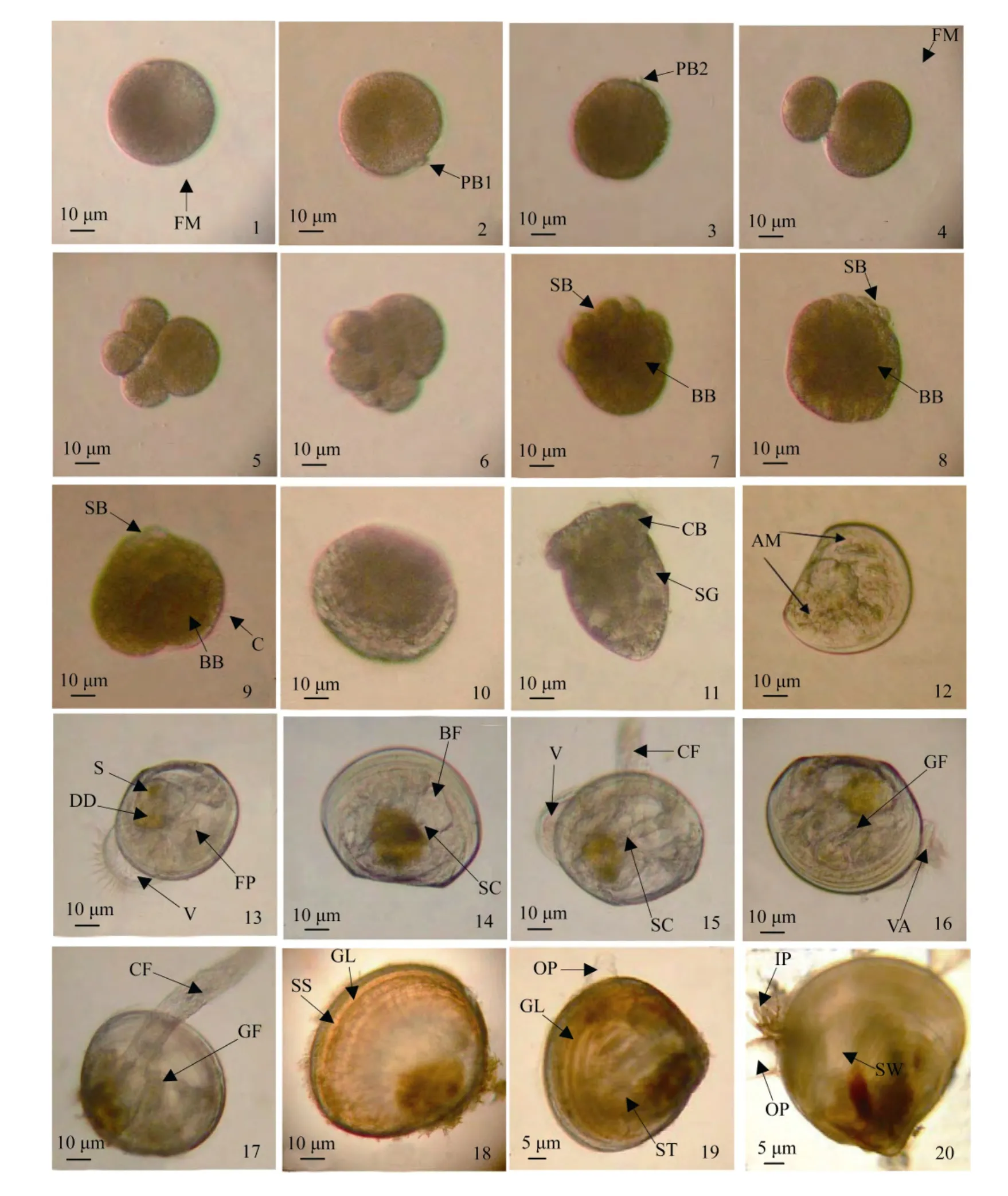
图版Ⅰ 皱肋文蛤胚胎、幼虫及稚贝发育过程的光镜观察PlateⅠ Microscopic observation on the development of embryo, larvae and spat of Meretrix lyrata
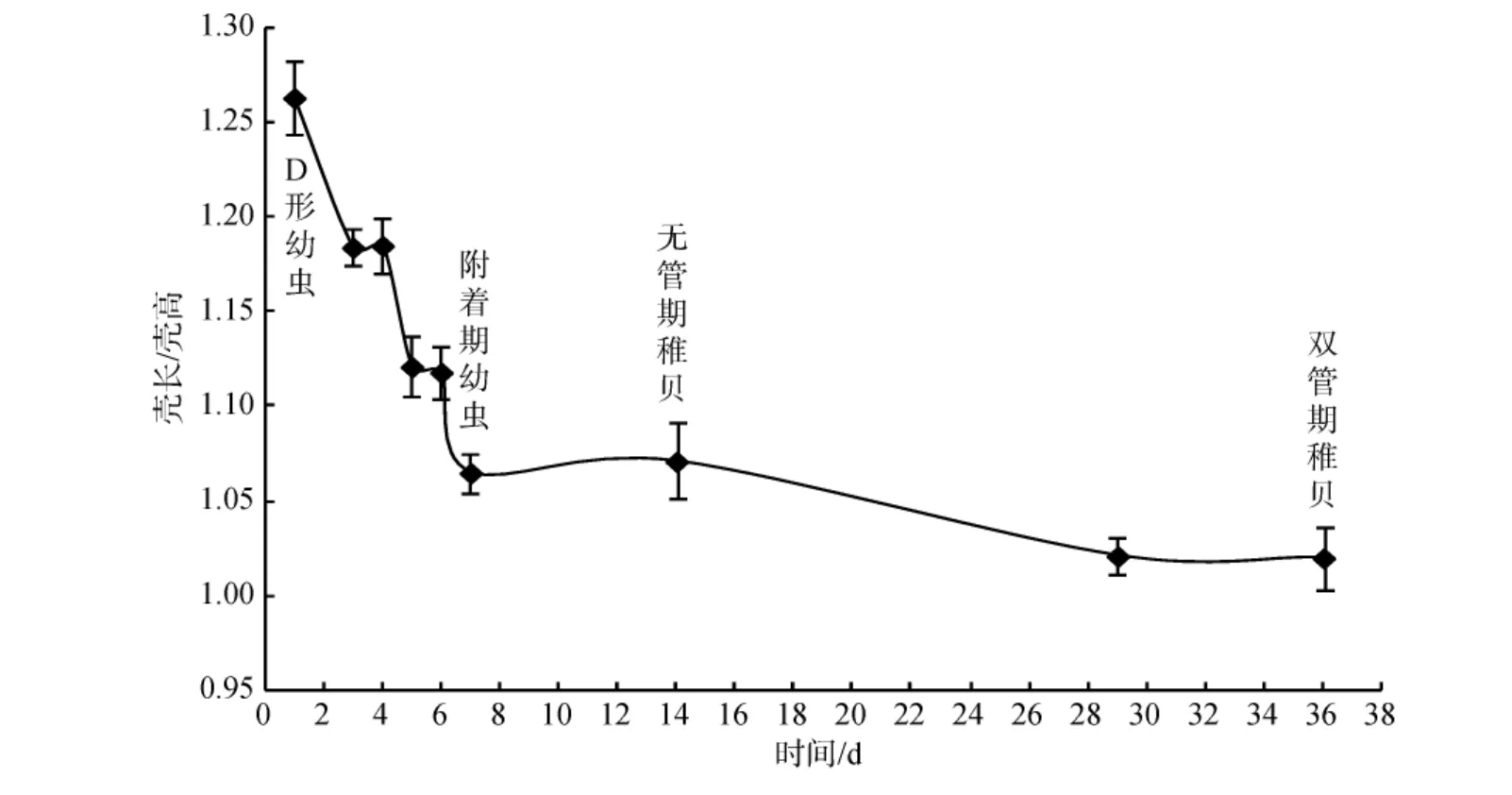
图2 皱肋文蛤幼虫和稚贝的壳长与壳高的比值Fig.2 The ratio of shell length and shell height for larvae and spat of Meretrix lyrata
皱肋文蛤雌雄异体, 一般1龄性成熟, 成熟性腺分布在内脏团周围, 并延伸至足基部。雌性性腺呈乳白色, 雄性性腺呈浅黄色。精子属于鞭毛型, 成熟卵子充满大量卵黄, 正圆形, 呈现均黄卵和沉性卵特性。皱肋文蛤属体外受精, 精子入卵的部位是随机的,可以从卵的任意部位入卵, 这与栉孔扇贝[13]、太平洋牡蛎[14]、泥蚶[15]菲律宾蛤仔[16]等贝类相似。皱肋文蛤精子和卵子在水中结合完成受精, 受精卵大小均匀, 卵径80μm。受精后50 min左右, 受精卵进行分裂, 卵裂方式为完全不等分裂, 第一次分裂为纵裂,分裂线与卵子动植轴平行偏于一侧, 纵裂成 2 个大小不等的分裂球, 即为2细胞期, 二者之间的卵裂沟十分明显。第二次卵裂与第一次卵裂过程基本相同,也是纵裂, 不过与第一次卵裂方向垂直。在第一次和第二次分裂前均无极叶形成, 这与厚壳贻贝[17]、渤海鸭嘴蛤[18]、大獭蛤[19]胚胎发育不同, 与文蛤[20]、中国蛤蜊[21]和缢蛏[22]相同。受精后1h 50 min, 进行第三次卵裂, 仍为不等全裂, 但此后开始进行螺旋分裂, 左旋分裂和右旋分裂交替进行。
3.3 皱肋文蛤幼虫和稚贝发育
海洋软体动物主要有 3种幼虫发育形式[23-24]:一是浮游异养型, 幼虫主要以浮游植物为食; 二是卵黄营养型, 幼虫以自身的卵黄为营养, 极少数甚至不依赖浮游生物; 三是直接发育型, 整个胚胎和幼虫发育过程在卵壳内进行, 不经历浮游阶段而直接进入成年底栖生活。皱肋文蛤在发育过程中经历担轮幼虫期和面盘幼虫期, 属间接发育, 幼虫阶段营养来源依赖浮游植物, 因此, 皱肋文蛤幼虫发育形式属浮游异养型。
双壳贝类幼虫从壳顶后期进入变态附着时, 其外部形态、内部构造、生理机能和生态习性等方面发生较大的变化。皱肋文蛤壳顶后期进入变态附着时, 行为方式从浮游生活转变为短暂的匍匐生活,最后附着生活。壳顶由微隆起逐渐变圆, 面盘逐渐脱落, 开始以棒状足作为运动器官。鳃丝逐渐增多, 出现明显的平衡囊, 消化系统逐渐完善, 摄食能力增强。许多双壳贝类, 如牡蛎、扇贝、珍珠贝等[25]在结束浮游生活即将进入变态附着时, 可以看到在鳃原基的背部形成 1对球形的由黑色素聚集起来的感觉器官, 称为眼点。因眼点是贝类幼虫即将附着变态时显而易见的特征, 所以一般以眼点的出现作为幼虫附着变态的标志。但对皱肋文蛤幼虫附着变态阶段的观察结果表明, 该贝在幼虫附着变态时并未出现眼点, 进一步证实了作者此前的结论[8]。双壳贝类在附着变态阶段没有眼点的现象也出现在施氏獭蛤[26]中。皱肋文蛤幼虫附着变态过程中虽然没有眼点出现, 但有明显的平衡囊, 在光镜下可清晰地看到平衡囊呈透明的同心圆状, 位于足的基部, 其可作为皱肋文蛤幼虫附着变态的标志。
皱肋文蛤在担轮幼虫初期发育阶段, 胚体背部后半段细胞加厚并逐渐陷入, 成为壳腺。以后壳腺分泌出2片透明而柔软的薄膜, 紧贴胚体的两侧, 背部铰合, 腹部开启, 薄膜钙化后形成两片且半透明的贝壳, 此壳称为胚壳或初生壳。随着皱肋文蛤幼虫的继续发育, 外套膜逐渐形成, 在附着变态期之后, 在胚壳的基础上, 外套膜开始分泌形成新的贝壳, 该种贝壳称为次生壳。皱肋文蛤次生壳的形成, 标志着该贝类已发育到稚贝期。无管期稚贝的壳色在次生壳部分为红褐色, 壳面生长线明显且隆起。当稚贝壳长达到近 600μm 时, 在壳后背方长出出水管, 管口缺乏触手, 称为单管期稚贝。该期稚贝壳顶逐渐隆起变尖, 外观呈红褐色, 育苗生产上把此期称为“红壳期”。随着稚贝在壳后腹方长出进水管, 管口环绕触手, 标志双管期稚贝的形成, 此期稚贝贝壳边缘壳色变白而不透明, 育苗生产上把此期称为“白壳期”。此期稚贝器官和外部形态已接近成体。
3.4 皱肋文蛤早期发育阶段的生长规律
皱肋文蛤胚胎发育阶段从受精卵开始直至原肠胚, 虽然随着细胞不断分裂使细胞数目迅速增多,但胚胎体积变化较小。自受精卵经卵裂, 孵化至担轮幼虫, 胚体长径仅增到原有长度的 1.38倍。幼虫阶段从担轮幼虫开始直至附着期幼虫, 尽管 D形幼虫之后, 幼虫的消化道已经形成并开始摄食, 但由于摄食和消化等内部器官发育并不完善, 摄食量较小,导致幼虫阶段皱肋文蛤生长依然缓慢。从D形幼虫经壳顶初期幼虫、壳顶中期幼虫和壳顶后期幼虫变态发育到附着期幼虫, 其壳长和壳高分别增加到原有壳长和壳高的1.77倍和2.10倍(表1和表2)。皱肋文蛤稚贝阶段从无管期稚贝至双管期稚贝, 该阶段稚贝内部器官发育完善, 摄食量增加, 稚贝生长速度明显加快, 自无管期稚贝经单管期稚贝发育到双管期稚贝, 稚贝壳长和壳高分别增加到原有壳长和壳高的3.13倍和3.28倍。由此可见, 皱肋文蛤早期发育阶段呈现胚胎和幼虫阶段缓慢生长, 稚贝阶段快速生长的规律。
[1] 蔡英亚, 谢绍河.广东的海贝[M].汕头: 汕头大学出版社, 2006: 344-345.
[2] Nugegoda D, Phuong P, Nguyen D, et al.Bioaccumulation and Depuration of Complexed and Uncomplexed TraceMetals by the Asian ClamMeretrix lyratafrom Vietnam[C]//Goteborg: 2009 Annual Meeting of the UK Branch of the Society of Environmental Toxicology and Chemistry (SETAC 2009), 2009.
[3] Luc N T, Thoa N V. Chemical composition and nutrient value lyrate hard clamMeretrix lyrata[J]. Fisheries Review: 2003, 4: 14-16.
[4] Hao N V, Hung N D, Thanh P C, et al. Environmental Parameters, Biology and Stocks ofMeretrix lyratain the Mekong Delta, Vietnam[C]∥ Proceedings of the 10th International Congress and Workshop of the Tropical Marine Mollusc Programme. Phuket: Phuket Marine Biological Center, 2000, 2(1): 272-273.
[5] Tri N N, Lin C K. Mollusc Culture System and Environmental Conditions in Can Gio District, Ho Chi Minh City, Vietnam[C]∥ Proceedings of the Ninth Workshop of the Tropical Marine Mollusc Programme.Phuket: Phuket Marine Biological Center, 1999, 19 (1):185-190.
[6] Thom P V, Tuan V S. Preliminary Studies on the Relationship Between Environmental Conditions and Distribution of Some bivalves in the Coastal Waters of South Vietnam[C]∥ Proceedings of th e Ninth Workshop of the Tropical Marine Mollusc Programme(TMMP). Phuket: Phuket Marine Biological Center,1999, 19(1): 249.
[7] Tong P H S, Nguyen T T. The Relationship between natural conditions and the formation and development of clam grounds (Meretrix lyrata) in the Mekong Delta[J]. Environmental Change and Agricultural Sustainability in the Mekong Delta, Advances in Global Change Research, 2011, 45: 301-333.
[8] 栗志民, 刘志刚, 欧阳志, 等.皱肋文蛤人工育苗技术的研究[J].南方水产科学, 2012, 8(2): 36-42.
[9] 刘志刚, 刘建勇, 刘付少梅.不同潮位、密度及季节对皱肋文蛤中间培育效果的影响[J].海洋科学, 2011,35(10): 34-41.
[10] 栗志民, 刘志刚, 徐法军, 等.温度、盐度、pH和饵料密度对皱肋文蛤清滤率的影响[J].渔业科学进展,2011, 32(4): 55-61.
[11] 栗志民, 刘志刚, 徐法军, 等.体重、温度和盐度对皱肋文蛤耗氧率和排氨率的影响[J].海洋科学进展,2011, 29(4): 512-520.
[12] 栗志民, 刘志刚, 姚茹, 等.温度和盐度对皱肋文蛤幼贝存活与生长的影响.生态学报, 2010, 30(13):3406-3413.
[13] 杨爱国, 王清印, 孔杰, 等.栉孔扇贝受精卵减数分裂的细胞学观察[J].中国水产科学, 1999, 6(3): 96-98.
[14] 任素莲, 王德秀, 王如才, 等.太平洋牡蛎受精过程中的精核扩散与成熟分裂[J].海洋湖沼通报, 1999, (1):34-39.
[15] 孙慧玲, 方建光, 王清印, 等.泥蚶受精过程的细胞学荧光显微观察[J].水产学报, 2000, 24 (2): 104-107.
[16] 毕克, 包振民, 黄晓婷, 等.菲律宾蛤仔受精及早期胚胎发育过程的细胞学观察[J].水产学报, 2004, 28(6):623-627.
[17] 顾忠旗, 倪梦麟, 范卫明.厚壳贻贝胚胎发育观察[J].安徽农业科学, 2010, 38(32): 18213-18215.
[18] 黄加棋.渤海鸭嘴蛤胚胎发育的研究及其与缢蛏胚胎的比较[J].厦门大学学报自然科学版, 1984, 23(2):224-231.
[19] 李琼珍, 童万平, 苏琼, 等.大獭蛤的胚胎、幼虫及稚贝的形态发育[J].广西科学, 2003, 10(4): 296-299.
[20] 董迎辉, 林志华, 柴雪良, 等.文蛤受精及早期胚胎发育过程的细胞学观察[J].动物学报, 2007, 53(4):700-709.
[21] 沈永忱, 郑宝太, 刘吉明, 等.中国蛤蜊的诱导产卵及胚胎发育[J].齐鲁渔业, 2005, 22(3): 1-3.
[22] 齐秋贞, 杨明月.缢蛏浮游幼虫、稚贝和幼贝的生长发育[J].台湾海峡, 1984, 3(1): 90-99.
[23] Thorson G.Reproductive and larval ecology ofmarine bottom invertebrates[J].BiolRev, 1950, 25: 1-45.
[24] Jablonski D, Lutz R A. Molluscan Larval Shellmorphology:Ecological and Paleontological Applications [M] //RhoaksD C, LutzR A. Skeletal Growth of Aquatic Organisms.New York: Plenum Press, 1980: 323-377.
[25] 王如才, 王昭萍, 张建中.海水贝类养殖学[M].青岛:中国海洋大学出版社, 2008: 134-136, 195-199,272-276.
[26] 刘永, 梁飞龙, 毛勇, 等.施氏獭蛤人工育苗技术的研究[J].湛江海洋大学学报, 2006, 26(03) : 98-101.
