华贵栉孔扇贝四种壳色选育系F1遗传多样性的AFLP分析
2015-12-02孙宗红刘志刚欧燕燕朱晓闻刘锦上
孙宗红 , 刘志刚 , 欧燕燕 , 朱晓闻 刘 超 , 刘锦上
(1. 广东海洋大学 水产学院, 广东 湛江 524025; 2. 广东高校热带海产无脊椎动物养殖工程研究中心, 广东 湛江 524025; 3. 湛江银浪海洋生物技术有限公司, 广东 湛江 524025)
扩增片段长度多态性(Amplified Fragment Length Polymorhism, AFLP)是1993年由荷兰Keygene公司科学家发明的一种 DNA分子标记技术[1], 该技术既继承了RFLP的稳定性, 又具有PCR反应快速、灵敏的特点, 同时克服了RFLP和RAPD的缺点, 且扩增的带纹多(50~100条)。AFLP的扩增片段与基因组的单一位置相对应, 且该标记为孟德尔式遗传, 可以作为联系遗传图谱和物理图谱的一个桥梁。
贝类壳色作为一个可遗传的质量性状, 已经被相关学者[2]成功应用到实际工作中。华贵栉孔扇贝是我国南方海域重要经济贝类, 近年来, 一些学者[3-7]对其生理和育种进行了大量研究。但相对于其他经济贝类如马氏珠母贝来讲, 采用分子标记来对华贵栉孔扇贝辅助育种的研究则非常少[8-9], 而对其不同壳色选育系 AFLP分析的研究尚未见报道。本文应用 AFLP标记技术对华贵栉孔扇贝四种壳色家系进行了遗传多样性分析, 旨在为该贝壳色的定向选育提供遗传理论基础, 加快从家系到壳色品系的建立, 并期望以此为基础, 筛选出与华贵栉孔扇贝颜色性状连锁的基因, 为华贵栉孔扇贝遗传图谱的构建提供参考。
1 材料方法
1.1 材料
1.1.1 样品采集
实验材料为本实验室选育的紫顶枣褐壳色(PBB)、枣褐壳色(BB)、紫白壳色(PW)和橘黄壳色(O)4种壳色选育系F1代(见图1)。随机挑选以上4种壳色华贵栉孔扇贝选育系F1代个体各30只, 清洗干净, 取少量闭壳肌(约 50 mg)用于基因组 DNA的提取, 剩余部分闭壳肌分装浸泡于无水乙醇中, –80℃保存。
1.1.2 引物及主要药品
本试验所用引物(表1)均由上海生工生物工程有限公司合成; Taq 酶、T4 DNA连接酶、EcoRⅠ内切酶、MseⅠ内切酶、DNA Maker DL2000、100 bp等均购自Takara公司; DNA提取试剂盒购自北京艾德莱生物科技有限公司。
1.2 AFLP实验步骤
1.2.1 基因组DNA的提取、酶切及连接反应
本实验使用DNA提取试剂盒提取基因组DNA,1%的琼脂糖凝胶检测, 用 Eppendorf核酸蛋白测定仪测定其纯度, 将DNA提取物稀释至300 ng/μL左右, 4℃保存。采用EcoRⅠ和MseⅠ双酶切方法酶切基因组 DNA, 37℃条件下EcoR I酶切 3 h, 65℃30 min终止反应, 将酶切产物65℃MseⅠ酶切3 h, 80℃30 min终止反应后马上进行连接反应, 16℃过夜, 65℃处理10 min终止反应。

图1 四种不同壳色华贵栉孔扇贝Fig. 1 Four kinds of Chlamys nobilis with different shell colors

表1 实验所使用的引物Tab. 1 All the primers used in the experiment
1.2.2 预扩增及选择性扩增反应
将连接产物进行预扩增反应, 94℃预变性2 min;94℃ 30 s, 56℃ 30 s, 72℃ 2 min, 共 23 个循环; 60℃延伸30 min, 产物4℃保存, 将产物稀释20倍进行选择性扩增反应, 选择扩增反应体系: 1.5μL10×PCR Buffer(含 Mg2+), 1.2 μL dNTPs(2.5 mmol/L), 1.0 μLEcoRⅠ引物 E (5 μmol/L), 1.0 μL MseⅠ引物 M(5 μmol/L), 0.1 μL Taq 酶(5 U/μL), 1.0 μL 预扩增稀释产物, 9.2 μL dd H2O。94℃预变性 2 min; 94℃ 20 s,66℃ 30 s, 72℃ 2 min, 然后每个循环降低1℃, 经10个循环后退火温度降低到 56℃, 再进行 20个循环, 条件为94℃ 20 s, 56℃ 30 s, 72℃ 2 min; 60℃延伸30 min。
1.2.3 变性聚丙烯酰胺凝胶电泳
选择性扩增产物用 5%的聚丙稀烯酰胺凝胶进行电泳分离、银染, 然后统计扩增位点数。
1.3 数据分析处理
将图谱条带按有记为“1”, 无记为“0”, 转换成 0, 1矩阵, 本实验按照显隐性将扩增位点上有带的视为显性基因型, 无带的视为隐性纯合基因型。利用Popgene1.31软件[10]计算多态位点比例、Shannon多样性指数、群体间遗传分化系数和遗传距离, 用MEGA4.1作图, UPGMA法进行聚类分析, 用SPSS17.0软件, ANOVA法进行方差分析。
2 结果
2.1 AFLP扩增结果
本实验扩增片段主要集中在150~1 000 bp之间。根据引物的筛选条件在64个引物组合中选出扩增条带数较多且清晰的10对AFLP引物组合, 在100~1 100 bp范围共扩增出414个位点, 四种壳色的华贵栉孔扇贝均在62个位点上出现扩增带, 多态位点数为 352个, 引物的多态性比例分布为79.31%~89.47%,平均为 85.02%, 多态信息含量较高。平均每个引物组合得到41个扩增位点, 但不同引物组合得到的扩增位点总数也存在很大差异, 比如 E3M5组合只得到 27个扩增位点(图 2、表 2), 而 E1M8得到 62个扩增位点(表2)。
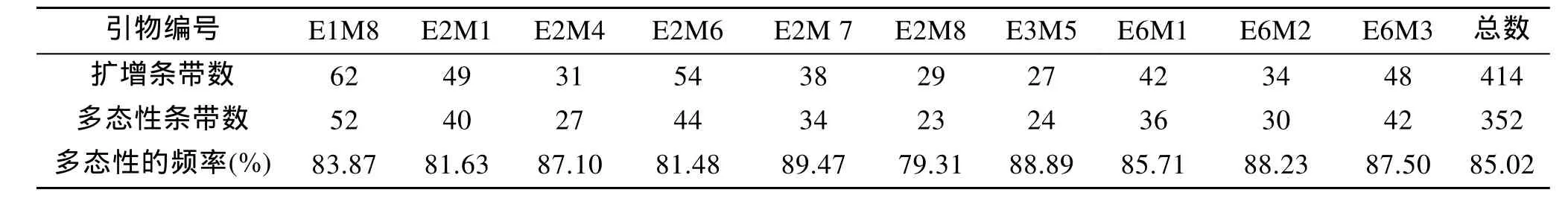
表2 实验选用的10对AFLP引物及扩增结果Tab. 2 The amplification results of selected ten pairs AFLP primers
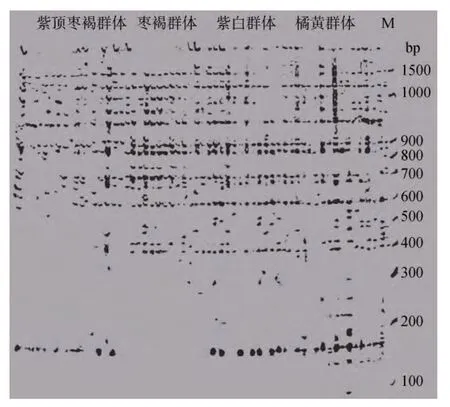
图2 引物E3M5在4种壳色华贵栉孔扇贝群体中的选择性扩增结果Fig. 2 The selective amplification results with primer E3M5 in 4 populations of the Chlamys nobilis
2.2 四种壳色华贵栉孔扇贝群体内遗传多样性
表3显示枣褐群体的多态性最大为86.04%, 橘黄群体的多态性最小为67.35%; 10对引物共扩增出414个位点, 而每个群体的总扩增条带数都小于414,与总条带数相比, 橘黄群体相差最多(71个), 而枣褐群最少(27个)。四个群体的Nei’s(H)指数在0.307 1~0.349 4之间, 其中, 枣褐群体的遗传变异最大, 其次是紫白、橘黄, 最小是紫顶枣褐群体。有效等位基因数(Ne)在1.519 1~1.616 6。Shannon(I)指数分布在0.452 5~0.513 9之间。四个群体的平均等位基因数和Shannon(I)与多态位点比例具有相同的变化趋势, 均为枣褐群体>紫白群体>紫顶枣褐群体>橘黄群体;平均有效等位基因和Nei’s基因多样性则是枣褐群体>紫白群体>橘黄群体>紫顶枣褐群体。遗传参数显示,Ht为0.395 4;Hs为0.326 6,Gst为 0.173 9,Nm为2.374 4(表 4), 说明群体间有一定分化, 不同群体间存在遗传差异, 各种群间存在一定的基因流动。
2.3 四种壳色华贵栉孔扇贝群体间的遗传多样性分析
2.3.1 四个群体间的遗传距离
四个群体间 Nei’s遗传距离(表 5)。由表可知,华贵栉孔扇贝四个群体间之间遗传距离(D)的变异范围在0.119 4~0.172 0之间。遗传距离最远的是紫顶枣褐群体和紫白群体, 最近的是枣褐群体和紫白群体。
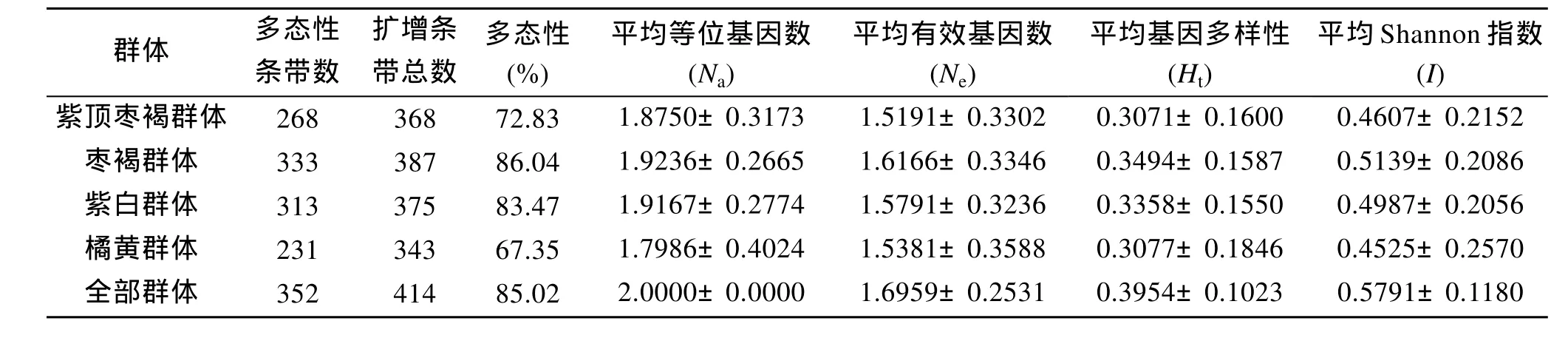
表3 四种不同壳色华贵栉孔扇贝群体的遗传结构Tab. 3 Genetic structures of 4 populations of Chlamys nobilis with different shell colors

表4 四个群体内的基因多样性和基因流Tab. 4 Gene diversity and gene flow of four populations
利用UPGMA法构建4个群体间的遗传关系聚类图(图 3)。聚类系统发生树均分为两个大支, 其中紫顶枣褐群体聚为一支, 其他三个群体聚为一支,以枣褐和紫白群体遗传关系最近, 紫顶枣褐群体与其他三个群体遗传关系最远。亲缘关系由近及远顺次为紫白壳色(PW)、枣褐壳色(BB)、橘黄壳色(O)和紫顶枣褐壳色(PBB)。
2.3.2 群体遗传多样性来源的分子方差(AMOVA)分析
应用分子方差分析(AMOVA)方法对4个群体的遗传多样性来源进行分析(表6)。由表可知群体内的变异占80.16%, 群体间的变异占19.84%。Nei’s遗传多样性分析方法与分子方差分析(AMOVA)得出的结论一致, 群体内遗传变异大于群体间。
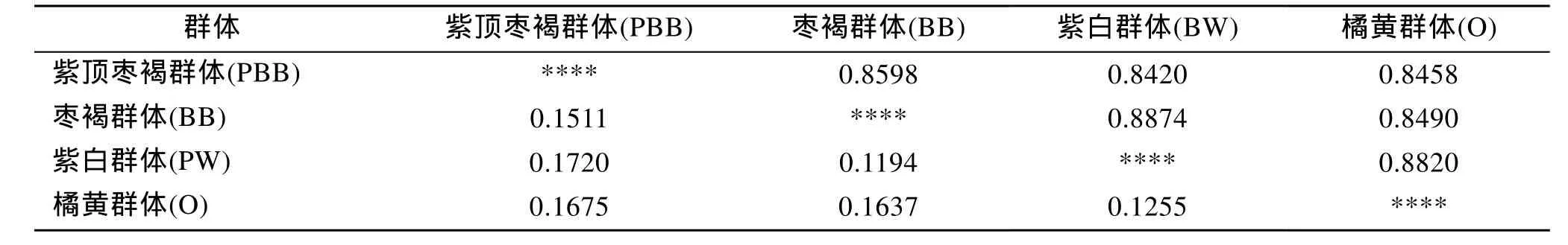
表5 华贵栉孔扇贝群体间的遗传距离及两两种群的遗传分化指数Tab. 5 Nei’s genetic distance and pairwise Fst among the stocks of Chlamys nobilis
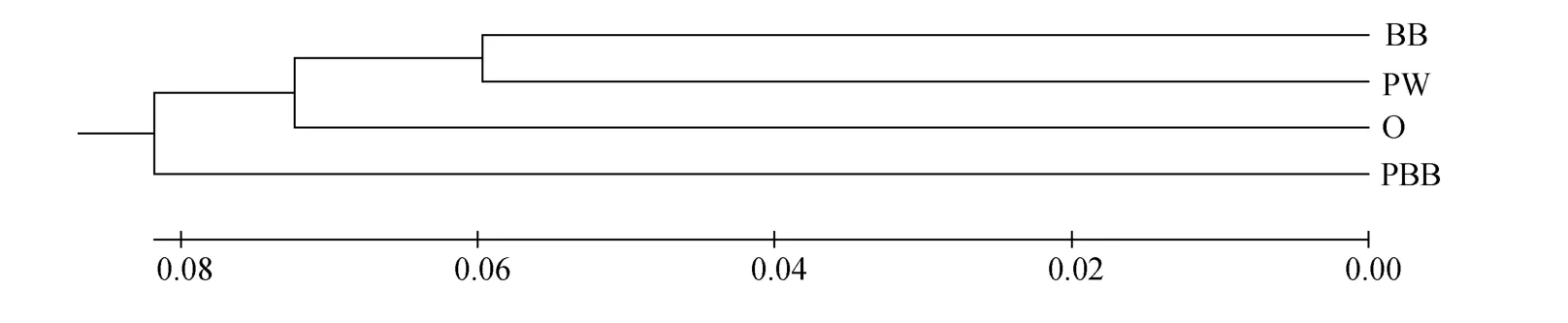
图3 四种不同壳色华贵栉孔扇贝群体间的聚类关系Fig. 3 Phylogenesis of four kinds of Chlamys nobilis with different shell colors
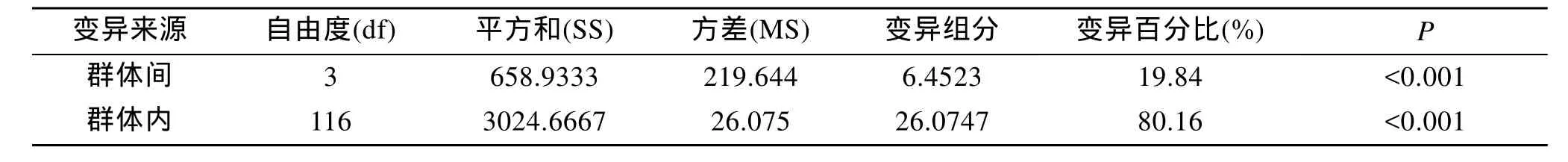
表6 四个群体的AMOVA分析Tab. 6 AMOVA analysis of four populations
3 讨论
3.1 四种壳色华贵栉孔扇贝种群内遗传多样性分析
薛钦昭等[11]的研究表明海洋贝类具有较高的遗传变异, 多态位点比例平均为 46%, 脊椎动物多态性平均值为24.7%, 且AFLP分子标记很适用于对亲缘关系及遗传上区别不大的种类的检测。本实验就是通过 AFLP检测分析出华贵栉孔扇贝物种水平上的多态位点比例达85.02%(表2、表3), 群体内总体的遗传分化Ht为0.3954(表4), 说明了群体多态性较高, 群体间存在一定的遗传分化。从欧燕燕等[12]的研究可知, 橘黄壳色群体本身较多, 同时因市场需求及其经济价值较高, 人们普遍留选橘黄色个体做繁殖亲本, 这可能是导致其遗传多样性低于其他群体的原因。而大量研究[13-15]也证实, 选育群体的遗传多样性会低于自然群体或野生群体。同时也有研究[15-16]证实, 只要保证亲本数量, 选育群体仍可保持较高的遗传多样性。本实验的样本是来自实验室培育的4种壳色的华贵栉孔扇贝选育系F1代, 通过遗传多样性分析表明了群体内的遗传多样性是较为丰富的,均具有较高的遗传多样性。
华贵栉孔扇贝的遗传多样性应该是怎样的水平才算合理, 迄今没有系统的研究, 也没有参考的标准, 本研究的数据可以作为衡量华贵栉孔扇贝各群体遗传变异的一个相对指标。
3.2 四种壳色华贵栉孔扇贝种群间遗传多样性分析
在分子生物学研究方面, 张涛[17]用5个SSR多态性位点对 3种壳色华贵栉孔扇贝分析发现各壳色间具有较高的遗传多态性, 存在一定分化。Yuan等[18]对华贵栉孔扇贝的养殖和野生群体的线粒体 16S rRNA和COI基因片段序列进行了研究, 表明了野生群体的基因差异水平高于养殖群体; Yuan等[9]用373个AFLP和9个微卫星标记, 通过拟测交策略分别构建了华贵栉孔扇贝雌性和雄性的遗传连锁图谱。
刘广绪[19]对栉孔扇贝、华贵栉孔扇贝及其二者的种间杂交子代、种内近交子代进行了遗传关系的分析, 发现杂交子代群体具有较高的遗传多样性,且两种杂交子代间也具有巨大的差异。邓岳文等[20]研究发现华贵栉孔扇贝桔黄壳色群体的各经济性状均值均显著大于紫顶枣褐壳色群体。
本研究在 4种壳色华贵栉孔扇贝群体中, 用 10对选择性扩增引物共扩增得到 414个位点, 其中多态性位点 352个, 多态性比例为 85.02%, 通过计算分析, 其群体间遗传分化系数(Gst)为 0.173 9, 基因流动系数(Nm)2.374 4亲缘关系依次为紫白壳色>枣褐壳色>橘黄壳色>紫顶枣褐壳色, 具有较高的遗传多样性, 表明了 AFLP技术适合用于华贵栉孔扇贝种群的遗传多样性研究。卢建峰等[21]对大口黑鲈选育群体遗传多样性进行了AFLP分析, 92.48%的遗传变异来自于群体内, 表明了大口黑鲈在遗传上的稳定性, 其群体具有一定选育潜力。而本实验应用分子方差分析方法对 4个群体的遗传多样性来源进行分析, 结果显示群体内遗传变异占 80.16%,说明各壳色群体具有一定选育潜力, 可继续进行人工选育。
3.3 壳色可作为贝类育种标记
在海洋贝类壳色的分子研究方面, 陈静, 朱晓闻, 代悦等[22-24]各自对马氏珠母贝 4种壳色的不同选育系进行了研究, 为其定向选育奠定了理论基础。程鹏[25]研究表明了华贵栉孔扇贝群体内遗传结构的差异, 造成了不同壳色家系在幼虫阶段生长性状上的不同。上述研究证实了在海洋贝类的选育中, 利用壳色作为依据可行, 且在实际应用中也得到了验证[2]。陈金涛[26]研究表明了华贵栉孔扇贝不同壳色子代在生长、存活率、出柱率(AP)及性状参数间的相关性方面存在显著差异。本实验也是以华贵栉孔扇贝的 4种壳色作为育种标记, 从分子水平上阐明了这 4种壳色选育系 F1代间存在遗传差异, 群体内的遗传多样性较高, 群体间存在一定分化, 可继续进行选育,为其定向选育提供了理论基础。
[1] 田舜, 李韬. AFLP技术操作流程的变革及其衍生技术[J]. 扬州大学学报, 2006(4): 62-65.
[2] 张跃环, 闫喜武, 张澎, 等.贝类壳色多态的研究概况及展望[J]. 水产科学, 2008, 27(12): 680-683.
[3] 栗志民, 刘志刚, 谢丽. 体重和温度对华贵栉孔扇贝(Chlamys nobilis)耗氧率和排氨率的影响[J]. 海洋与湖沼, 2010, 41(1): 99-105.
[4] 郑怀平, 孙泽伟, 张涛, 等. 华贵栉孔扇贝 1龄贝数量性状的相关性及通径分析[J]. 中国农学通报, 2009,25(20): 322-326.
[5] 邓岳文, 王庆恒, 黄荣莲, 等. 华贵栉孔扇贝基础群体内大小分化个体的淀粉酶和纤维素酶活力比较[J].海洋学研究, 2008, 26(2): 37-40.
[6] 刘志刚, 朱晓闻, 王辉, 等.华贵栉孔扇贝壳色遗传规律的研究[J].海洋与湖沼, 2012, 43 (1): 62-66.
[7] 刘志刚, 王辉, 吕文刚, 等. 华贵栉孔扇贝壳色与闭壳肌颜色的遗传规律[J].海洋与湖沼, 2012, 43(2):237-243.
[8] 张涛, 郑怀平, 孙泽伟, 等. 华贵栉孔扇贝不同壳色后代早期发育阶段性状比较[J]. 中国农学通报, 2009,25(23): 478-484.
[9] Yuan T, He M, Huang L, et al. Genetic linkage maps of the noble scallop chlamys nobilis reeve based on aflp and microsatellite markers[J]. Journal of Shellfish Research, 2010, 29(1): 55-62.
[10] 王学颖, 高远镇, 杜晓东, 等.马氏珠母贝金黄壳色系 F3 和基础群体遗传结构比较[J].海洋通报, 2012,31(3): 324-328.
[11] 薛钦昭, Stiles Sheila, 张福, 等. 海湾扇贝不同种群在磷酸葡萄糖变位酶基因位点的遗传结构与性状[J].海洋与湖沼, 1993, 30(4): 80-83.
[12] 欧燕燕, 刘志刚, 刘锦上. 流沙湾养殖华贵栉孔扇贝体色多态性研究[J].南方水产科学, 2012, 8(5): 15-24.
[13] 张国范, 张福绥. 贝类遗传多样性及其永续利用(I)[J]. 海洋科学, 1993(5): 17-21.
[14] 郭俊, 邓岳文, 杜晓东, 等. 马氏珠母贝黄壳色选育系G_1遗传结构的ISSR分析[J]. 广东海洋大学学报,2009, 29(1): 87-90.
[15] 赵晓霞, 邓岳文, 杜晓东, 等. 利用微卫星标记分析马氏珠母贝 4个养殖群体遗传结构[J]. 基因组学与应用生物学, 2010, 29(5): 879-884.
[16] Yu H, Li Q. Genetic variation of wild and hatchery populations of the pacific oysterCrassostrea gigasassessed by microsatellite markers.journal of genetics and genomics, 2007, 34(12): 1114-1122.
[17] 张涛. 华贵栉孔扇贝养殖群体数量性状及壳色遗传的初步研究[D].汕头: 汕头大学, 2010.
[18] Yuan T, He M, Huang L. Intraspecific genetic variation in mitochondrial 16S rRNA and COI genes in domestic and wild populations of Huaguizhikong scallop Chlamys nobilis Reeve[J]. Aquaculture, 2009, 289(1-2): 19-25.
[19] 刘广绪. 栉孔扇贝、华贵栉孔扇贝及其种间杂交子代、种内近交子代的ISSR分析[D].青岛: 中国海洋大学, 2004.
[20] 邓岳文, 张凌飞, 杜晓东, 等. 华贵栉孔扇贝两种壳色群体生长和消化酶活力比较[J]. 广东海洋大学学报, 2008, 28(3): 20-23.
[21] 卢建峰, 白俊杰, 李胜杰, 等. 大口黑鲈选育群体遗传多样性的AFLP分析[J]. 淡水渔业, 2010, 40(3): 3-7.
[22] 陈静, 刘志刚, 孙小真, 等. 马氏珠母贝 4种壳色选育系 F_3的生长特性和遗传多样性比较[J]. 上海海洋大学学报, 2010, 19(5): 588-595.
[23] 朱晓闻, 刘志刚, 王辉, 等. 马氏珠母贝四种壳色选育系 F5的生长及遗传多样性[J].海洋与湖沼, 2012,43(1): 160-165.
[24] 代悦, 喻子牛, 赵晓霞, 等. 马氏珠母贝 4个壳色选育系F_1遗传结构的AFLP分析[J]. 基因组学与应用生物学, 2010, 29(2): 252-258.
[25] 程鹏, 杨爱国, 周丽青, 等. 不同壳色虾夷扇贝家系F_1幼虫生长及遗传结构的比较分析[J]. 中国水产科学, 2010, 17(5): 960-968.
[26] 陈金涛. 华贵栉孔扇贝不同壳色选育系的建立及其生长发育研究[D]. 湛江: 广东海洋大学, 2011.
