不同三疣梭子蟹混养系统能量收支的研究*
2015-12-02田相利董双林
奉 杰,田相利,董双林,张 凯,董 佳
(中国海洋大学水产学院,山东 青岛 266003)
多种类混养是我国海水池塘养殖的主要养殖方式之一,也是中国海水池塘养殖的特色之一。它不仅可以增加池塘生态系统的结构和空间成层性,提高池塘中生物群的多样性和环境的稳定性,从而提高投入物质的利用率,增强养殖水体的自净能力[1]。在中国沿海的海水池塘养殖中,梭子蟹是主要的养殖种类之一[2],而梭子蟹与对虾和贝类的混养则是比较常见的养殖模式之一。从生态学原理来讲,梭子蟹与对虾混养具有良好的生态学作用,不仅可以充分利用残饵,提高饲料利用率,还对抑制虾蟹类疾病的发生具有一定的作用[3]。而贝类的滤食作用和生物沉积作用则可以改善养殖水体的水质[4-8],加速养殖水体内有机质的循环利用,在一定程度上提高水体的养殖容量[9]。然而,在目前的实际生产中,梭子蟹、对虾和贝类放养密度和比例的确定多基于经验,随意性较大,缺少相关的理论指导。
物质循环和能量流动状况是衡量一个生态系统是否稳定的重要指标,一直是生态学者们进行生态学研究的焦点[10]。自1970年以来,国内外众多学者无论对于不同养殖系统的能量流动状况,还是养殖生物的个体能量学都做了大量的研究工作,例如,对综合养殖池塘的能量利用和转化情况[11-16]、高产单养对虾池[17-18]和工厂化养殖系统的能量收支和流动状况[19],以及温度[20-21]、盐度[22]和密度[23]等因素对养殖个体能量收支的影响等,都进行了广泛而深入的研究。关于混养生态系统的能量收支,国内外目前已有较多关于虾贝藻混养[15],虾鱼混养[24]的相关研究,不过,目前关于蟹、对虾和贝类混养系统的能量收支和转化效率的研究还少见报道。
鉴于此,本研究以海水池塘陆基围隔实验法[25]比较研究了三疣梭子蟹(Portunustrituberculatus)、凡纳滨对虾(Litopenaeusvannamei)、菲律宾蛤仔(Rudi-tapesphilippinarum)不同混养系统的能量输入与输出状况,以及能量利用和转化效率,以期通过对优化、筛选出具有较高能量利用率兼较高养殖产量的混养模式,从而为中国梭子蟹的高效生态养殖技术的提高提供相关的理论依据。
1 材料与方法
1.1 实验池塘与实验材料
本研究于2012年7~10月在江苏省连云港市赣榆县佳信水产养殖场进行,养殖池塘为泥沙质底,主要以海水池塘陆基实验围隔为实验设施进行研究,围隔大小大约为5m×5m。围隔的具体结构参见文献[25]。
实验用三疣梭子蟹、凡纳滨对虾、菲律宾蛤仔均购自连云港市赣榆县佳信水产养殖场。梭子蟹放养时平均甲宽(2.13±0.17)cm,平均甲长(1.09±0.09)cm,平均体重(0.57±0.13)g;凡纳滨对虾放养时平均体长(1.41±0.22)cm,平均体重(0.045±0.010)g,菲律宾蛤仔放养时平均壳长(1.55±0.13)cm,平均壳宽(2.34±0.16)cm,平均壳高(1.00±0.08)cm,平均体重(0.91±0.12)g。
1.2 实验设计
研究设置了7个不同处理,每个处理分别设置4个重复,一共使用了28个实验围隔。具体放养情况详见表1所示。
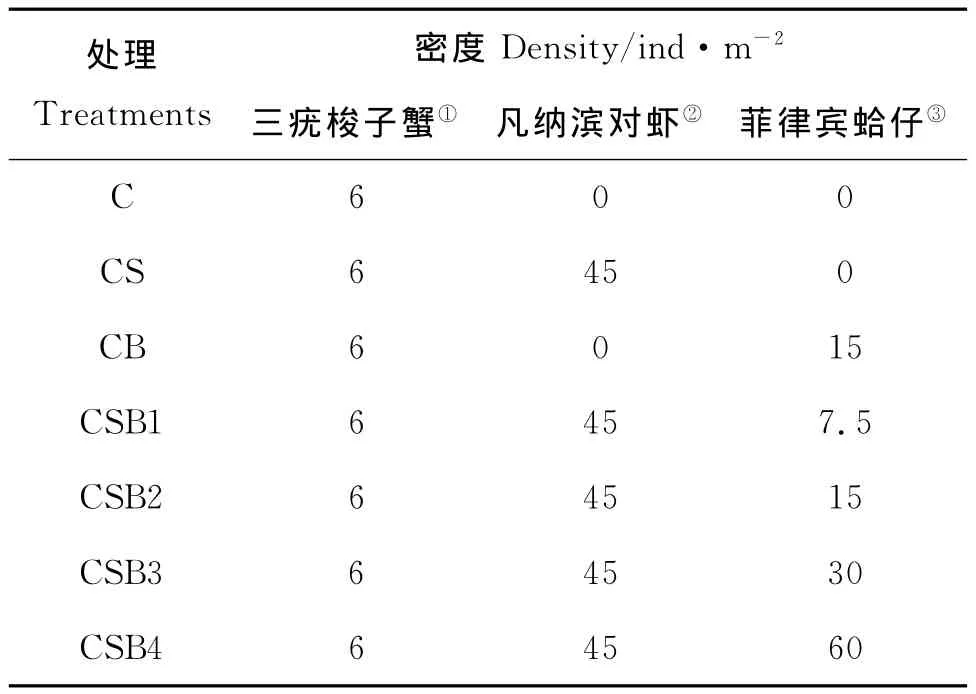
表1 各实验组的放养情况Table 1 Stocking information for different treatments
1.3 实验管理
实验开始前一个月用100g/m3漂白粉消毒,围隔于2012年3月18日进水完毕。在2012年7月2日放养三疣梭子蟹和菲律宾蛤仔苗,7月14日放养凡纳滨对虾苗;2012年10月3日开始对养殖生物进行收获。实验期间不换水,只补充蒸发和渗漏水,保持水深1.2~1.5m。7月中旬开始使用增氧机,根据天气状况,调整充气时间与时长。
实验期间每天7:00和19:00投喂2次。梭子蟹饲料主要投喂兰蛤(Aloidislaevis),对虾饲料选用配合饲料(青岛六和海洋科技有限公司,青岛)。围隔内设置投饵盘用于及时检查虾蟹的摄食情况。每隔10~15d检查1次生长情况,测定对虾体长,梭子蟹的甲长、甲宽,同时每天观察摄食情况与残饵量,根据生长情况并综合每天吃食情况、天气、水温及时调整和确定适宜的投喂量。
每天上午6:00用海水温度计测定水温(WT),每5 d用野外便携式溶氧仪(SX716,上海三信仪表厂)测定溶氧(DO),pH和盐度,每5d用便携式pH盐度计(Salinity &pH Monitor 2771,科立隆)测定1次。
1.4 水体初级生产力和能值测定及分析方法
浮游植物初级生产力测定:实验期间,每隔15d利用黑白瓶法测定一次各处理水体的初级生产力状况[26]。所用黑白瓶体积为250mL。每次水面下20cm处取水样5L混匀并充分曝气,经过200目(74μm)筛绢过滤,以去除大型浮游动物。将黑白瓶挂于水面下20cm处,挂瓶24h后,用Winkler法测定黑白瓶内溶氧含量的变化。具体操作和测定方法参考文献[27]进行。按照1mg O2=14.56J[28]的当量关系,计算水体浮游植物的初级生产力。
附生藻类初级生产力测定:围隔实验开始时在围隔内贴壁挂片收集附生藻类(挂片规格为25cm×150cm,挂片材料同围隔幔)。每15d进行1次采样,采样时,分别取位于水体表层、中层、底层围隔幔挂片剪取代表性样方[(1~2)cm×(1~2)cm]作为实验材料,置于250mL黑白瓶中,虹吸法注入取样围隔水样,在围隔内原位挂瓶培养,同时设原围隔水样挂瓶培养对照,测定围隔幔附生藻类的生产量。按照1mg O2=14.56J[28]的当量关系,计算水体附壁植物的初级生产力。
每隔7d测量1次照射到水体表面的光照辐射强度(TES-1330A型照度计,5:00~19:00每小时1次),并记录天气情况。
每15d收集1次水体沉积物,沉积物收集方法为:在各围隔内设3个沉降物收集管,开口距水体表面1.0m。沉降物质收集管采用圆柱形的PVC管(diameter:90mm,height:550mm),在管口处覆盖一块尼龙网(网眼0.8cm)以防止大型游泳生物进入。收集管在水体中放置7d后取出,带回实验室,低温静置。将上层水虹吸出,剩余的物质倒入大型培养皿(15cm)中,用蒸馏水洗去盐分,然后在60℃烘干48h,并用粉碎机粉碎。另外,在养殖生物放养、收获时分别留取适量养殖生物样品,在养殖过程中留存一定量的对虾饲料样品,梭子蟹饵料兰蛤样品,60℃下烘干后粉碎,用氧弹仪(PARR-1281,美国)分别测定沉积物、养殖生物样品、对虾饲料样品以及梭子蟹饵料生物样品能量含量。
1.5 能量折算系数和转化效率公式
将所有投入和产出物质均转化为能值(MJ)。其中,配合饲料、兰蛤、凡纳滨对虾、三疣梭子蟹和菲律宾蛤仔的能量折算系数(单位kg干重的养殖生物所含能量MJ)见表2。养殖系统能量转化效率(引自文献[14])按以下公式计算:
光能利用率=初级生产力(MJ)/太阳辐射能(MJ)×%100;
光合能的转化效率=生物净产出能(MJ)/太阳辐射能(MJ)×100%;
饲料能转化效率=生物净产出能(MJ)/饲料能(MJ)×100%;
总能量转化效率=生物产出能(MJ)/总输入能(MJ)×100%;
单位净产量耗饲料能=饲料能(MJ)/生物净产出量(kg);
单位净产量耗总能=总投入能(MJ)/生物净产出量(kg);
总输入能=输入生物能(MJ)+饲料能(MJ)+初级生产力(MJ);
总投入能=输入生物能(MJ)+饲料能(MJ)。

表2 不同处理投入与产出物质的能量折算系数Table 2Conversion coefficients of energy for input and output materials in different treatments
1.6 数据处理与分析
所有数据用spss17.0软件对数据进行分析,采用单因子方差(ANOVA)和(Duncan)多重比较进行分析处理,以P<0.05作为差异显著性水平。
2 结果
2.1 不同处理养殖动物收获情况
不同处理养殖动物收获情况见表3。可以看出,三疣 梭子蟹产量以CSB4最高,为(426.91±63.01)g/m2,CSB1最低,与CB、CS差异不显著(P>0.05);凡纳滨对虾产量以CSB3处理最高,为(420.94±63.88)g/m2,显著高于其它各处理组(P<0.05),CSB2处理最低,为(81.18±23.47)g/m2;菲律宾蛤仔产量总体随放养密度增加而增加,其中以CSB4处理最高,为(121.90±23.77)g/m2,显著高于其它各处理组(P<0.05),CSB1处理最低,为(23.83±7.49)g/m2。
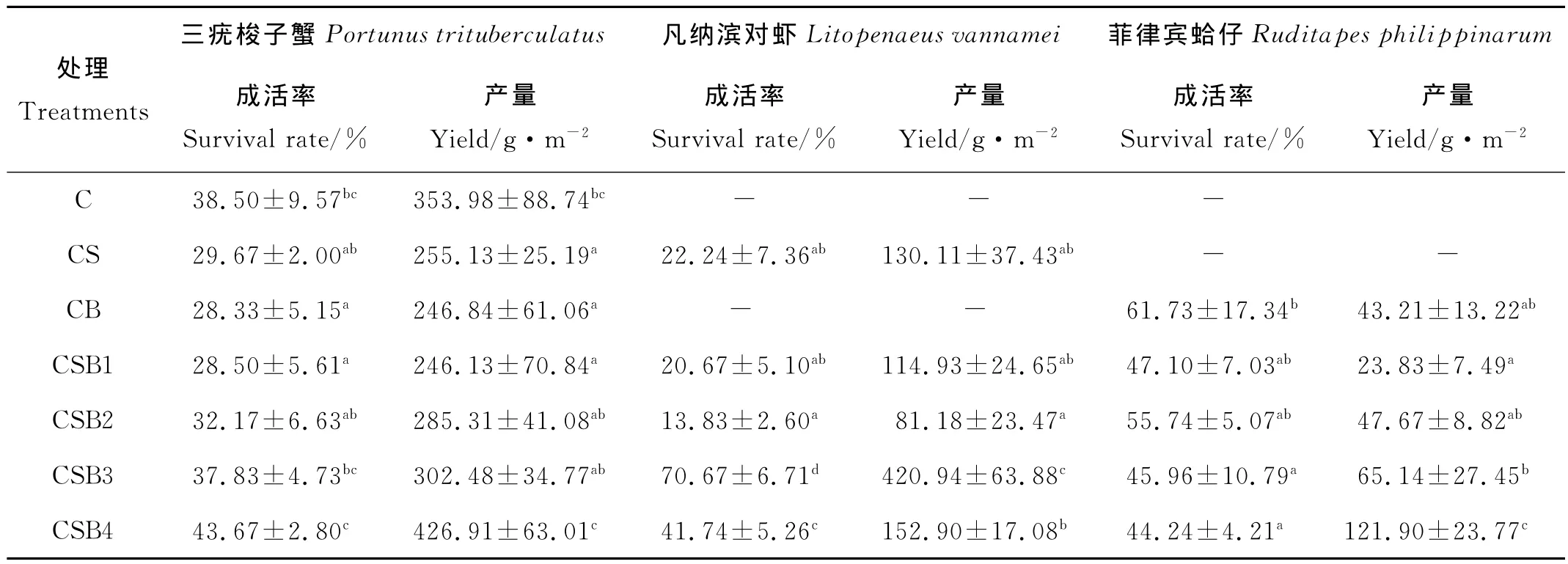
表3 不同处理养殖动物的收获情况Table 3 Harvest information of cultured animals in different treatments
2.2 各处理初级生产力和光能利用率
各处理初级生产力和光能利用率见表4。研究期间,整个系统接受的总太阳能为1 550MJ/m2。各处理初级生产力总体变动在2.54~5.81MJ/m2之间,以梭子蟹单养处理最高,显著高于CB、CSB1、CSB2和CSB4处理(P<0.05)。各处理光能利用率变化在0.16%~0.38%之间,总体趋势与初级生产力相似,以梭子蟹单养对照最高,显著高于CS、CB、CSB1和CSB2处理(P<0.05)。
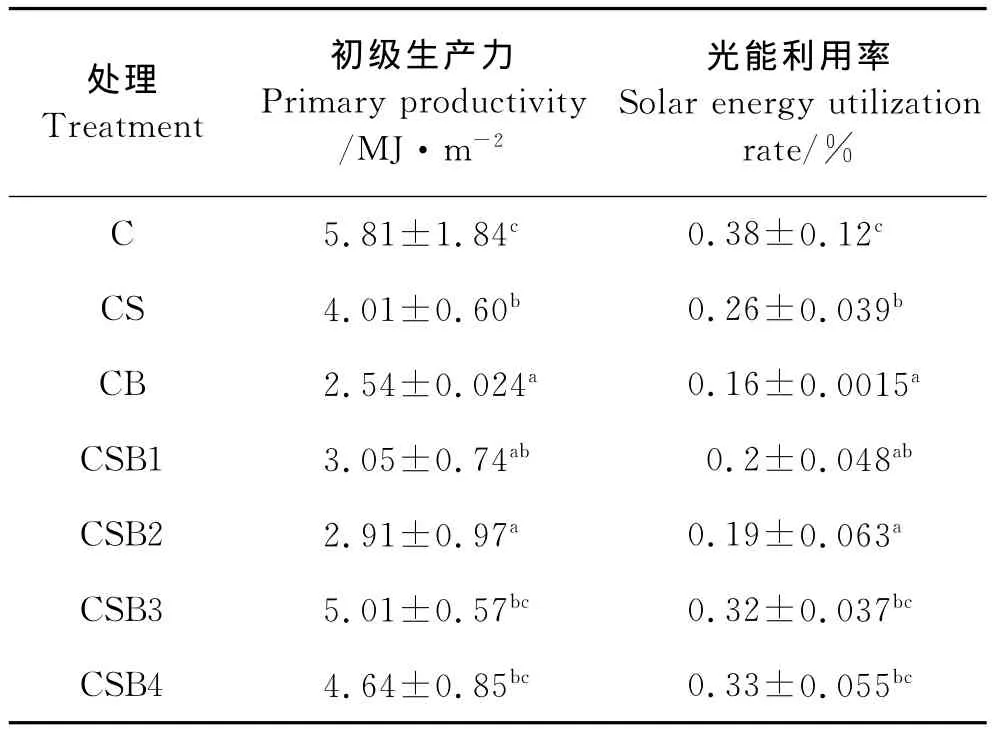
表4 不同处理的初级生产力和光能利用率Table 4 Primary productivity and solar energy utilization rates of different treatments
2.3 不同处理的能量收支和能量转化效率
2.3.1 不同处理的能量输入与输出 各处理能量输入情况见表5。各处理能量总输入量在130.33~412.60MJ之间,以CSB3处理最多,CB处理为最少,显著低于CS、CSB3和CSB4处理(P<0.05)。在总输入项目中,初级生产力与对虾饲料均占较大比例。初级生产力能量输入量最大的是梭子蟹单养,约占总输入量的67.72%,而CB处理则最低。饲料能在投入的总能量中所占比例在31.30%~69.39%之间,以CSB3处理最高,其对虾产量也最高。由养殖生物放养投入的生物能在总输入能量中所占比例较小,大约占总投入能量的0.11%~0.69%。
各处理能量输出情况如表6所示,主要项目为养殖动物的收获。各处理的能量输出量在20.47~71.53 MJ之间,以CSB3处理最大,CB处理最小。梭子蟹的能量产出中,以CSB4处理产量最高,其次是蟹单养对照,而CS、CB和CSB1处理显著小于CSB4、蟹单养对照(P<0.05)。梭子蟹的能量输出量在其总输出量中的所占比例分别为100.00%、58.00%、92.92%、57.05%、67.72%、32.62%和61.25%;凡纳滨对虾能量产量以CSB3处理最高,显著高于其它处理(P<0.05),而CSB2则为最低;菲律宾蛤仔的产出能量则以CSB4处理最高,CSB1处理最低。
2.3.2 各处理底泥沉积能量 各处理沉积物质量、能量以及沉积物能量在输入系统总能量中所占的比例见表7。实验期间,各处理沉积物质量变化于2.07~8.47 kg/m2之间,以CSB3处理为最高,CBS2处理为最低;沉积物能量在1.71~5.43MJ/m2之间,以CSB3处理最高,与CSB4处理差异不显著(P>0.05),但显著高于其它处理(P<0.05);各处理能量折算系数,即单位沉积物能值则以梭子蟹单养最高,显著高于CS、CB和CBS1处理;各处理沉积能量占输入总能量比例变化于25.57%~33.47%,各处理之间没有显著差异(P>0.05)。

表5 各处理的能量输入Table 5 The inputs of energy in different treatments /MJ

表6 各处理的能量输出Table 6 The outputs of energy in different treatments /MJ
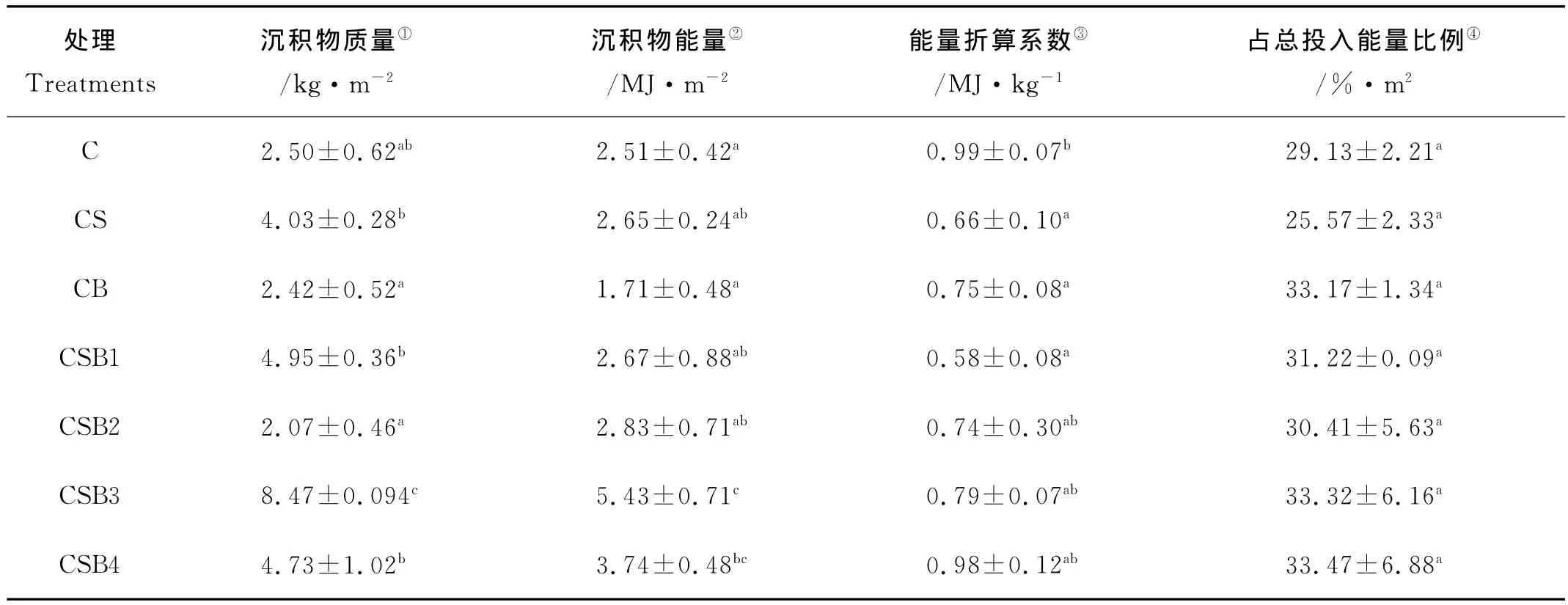
表7 各处理的沉积能量Table 7 Sedimented energy in different treatments
2.3.3 各处理的能量转化效率 各处理的能量利用、消耗和转化效率情况见表8。可以看出,光合能转化效率通体变动在0.57%~1.81%之间,以CBS3处理为最高,CBS4居其次,均显著高于其它处理(P<0.05),而CB处理和梭子蟹单养相对较低;各处理饲料能转化效率在21.23%~40.03%之间,其中最高为梭子蟹单养,为40.03%,但与CSB4和CB处理差异不大(P>0.05);总能量转化效率在13.69%~19.25%之间,各处理比较,以CSB4处理为最高,为19.25%,CSB3处理其次,为18.68%,CSB4、CSB3与除CSB2外的各处理组之间差异显著(P<0.05),其它处理之间未见显著差异(P>0.05);单位净产量耗饲料能在7.92~16.77 MJ·kg-1之间,以梭子蟹单养为最低,与CB和CSB4差异不大(P>0.05),但显著低于其它处理(P<0.05);不同处理单位净产量耗总能在7.96~16.86 MJ·kg-1,与单位净产量耗饲料能趋势一致,以梭子蟹单养为最低,与CB和CSB4差异不大(P>0.05),但显著低于其它处理(P<0.05)。综合以上各能量指标,可以看出CSB4处理在光合能转化效率和总能量转化效率处于较佳的位置,而饲料能转化效率、单位净产量耗饲料能和单位净产量耗总能等指标则与梭子蟹单养相差不大,为本实验中选出的最优搭配养殖模式。

表8 各处理能量的利用和转化效率Table 8Energy conversion coefficients in different treatments
3 讨论
建立高效,高产,稳定的池塘生态系统是相关领域研究者们一直以来的追求目标,对池塘生态系统中能量转换效率的研究是建立养殖资源高效利用养殖方式的核心问题之一[29]。而浮游植物通过光合作用将太阳能固定为生物能,是池塘中能量的主要输入部分之一,是生态系统中食物链或食物网的基础[30],无论对池塘中养殖生物的产量还是养殖水体中生态系统结构的建设都影响重大。由于不同的养殖系统之间因各种因素导致的差异,不同养殖系统中浮游植物初级生产力往往差异较大。在本研究中,各养殖系统接受的太阳辐射总能为1 550MJ/m2,而浮游植物初级生产力大小在2.54~5.81MJ/m2之间,光能利用率变化在0.16%~0.38%之间。与董贯仓[15]研究相比,这一数值要高于其对虾单养及虾贝混养处理,与虾藻混养处理相比则相差不大。在淡水养殖系统相关研究中,宋颀等[10]对混养鲢鱼(Hypophythalmichthysmolitrix)的草鱼(Ctenopharyngodonidellus)混养生态系统研究获得的光能利用率为0.25%,与本研究结果相似。但与李吉方等[14]盐碱地养殖池塘0.29%~0.62%、康春晓等[30]的草鲢养殖系统 0.46% 以及吴乃薇等[29]青鱼(Mylopharyngodonpiceus)养殖系统0.61%~0.96%的光能利用率数据相比,本研究结果处于较低水平。
在本研究中,输入的能量包括初级生产力、饵料和苗种三部分,各处理之间存在较大的差异。初级生产力和饵料投入是系统能量输入的最主要部分,其中,初级生产力占输入总能的比例在30.34%~67.72%之间,梭子蟹单养对照最高,显著高于其它处理(P<0.05);饵料占输入总能的比例在31.30%~69.39%之间。而苗种比例较低,仅占总输入的0.11%~0.69%。生物净产出能是养殖系统能量输出最受关注的部分,主要由系统的养殖动物产量决定。在本研究中,生物净产出能占总输入能的比例,也就是总能量转化效率,在13.69%~19.25%之间,以CSB3最高,CSB4处理其次,显著高于其它处理(P<0.05)。在输入能量被养殖生物利用的同时,实验过程中未被养殖生物完全摄食的生物饵料兰蛤和虾饲料会沉积在池底,加上养殖生物的排泄物以及死亡的浮游生物,沉积到池塘底部的能量成了本养殖系统能量输出的一个重要方面。在本研究中,各处理底泥总沉积能量占总输入能量25.57%~33.47%,各处理之间未见显著差异(P>0.05)。本研究中底泥沉积能量在1.71~5.43MJ/m2之间,低于宋颀等[10]草鱼混养系统 4.71~15.08 MJ/m2的沉积能量值,但高于董贯仓等[15]对虾混养系统0.53~1.30MJ/m2沉积能量值,推测原因,可能与养殖生物、水体环境以及系统的构成等存在的不同有关。
滤食性贝类主要以浮游植物、微生物和有机碎屑等为食,滤水率和摄食量都很大。现有研究表明,它不仅具有很强的生物沉积作用[31-33],还具有促进周边区域沉积物矿化作用的能力[33-35],因此可以加速养殖系统悬浮物质的沉降、加速有机质的循环利用。但关于其对水体浮游植物的影响,还存在一定的争议。部分研究表明,菲律宾蛤仔通过滤食养殖水体中的细小颗粒,减少水体中浮游植物的生物量[32,36],然而另有研究认为,贝类排泄和底层沉积物矿化作用再生的营养盐可以成为浮游植物初级生产的重要基础[37],营养盐再生可以促进初级生产力的发展[38],因此对处于同一生态环境中的浮游植物具有促生长作用[39]。本研究设置了不同菲律宾蛤仔密度处理,可以看出,贝类放养密度较高的CSB3和CSB4处理并未因贝类密度放养较大而降低浮游植物生物量,使得光能利用率降低;相反,在光能利用率较低的CB和CSB2处理,尽管贝类放养密度不高,但光能利用率却处于较低水平,即浮游植物的生物量较低。因此,CSB3和CSB4处理菲律宾蛤仔放养密度没有超过其可以对浮游植物造成很大摄食压力的阈值,浮游植物在满足其营养供应的情况下可能仍具有一定的生长空间。
池塘里合理混养滤食性贝类,不仅可以增加系统的物种多样性、分布空间的层次性,还可提高各种营养物的利用率,这已被以往的很多研究所确认[14,40]。而从系统能量流动角度分析,本研究中菲律宾蛤仔的滤食作用、生物沉积作用、加速向水体释放营养盐这一系列过程可能形成了养殖水体中新的能量循环途径,从而有效提高了系统的能量利用与转化效率。例如,CSB3和CSB4处理的光合能转化效率以及总能量转化效率均要显著高于梭子蟹单养对照。不过本研究中混养菲律宾蛤仔对于人工投入的饲料能转化效率提升作用不大,这一点与董贯仓等[15]对对虾混养系统的相关研究有所不同。这种差别是否与养殖的种类与混养结构的差异有关,尚待进一步研究。尽管蟹、虾、贝混养是养殖实践中常见的混养模式,但梭子蟹、菲律宾蛤仔和对虾均为底层生活种类,从提高对水体空间利用的互补性、空间多样性以及对营养盐利用考虑,适当混养中上层养殖动物(例如鱼类等)和大型藻类,可能是下一步研究需要关注的问题。
综合各能量指标分析可以看出,CSB4处理的总能量转化效率在各处理组中处于最高水平,光合能转化效率仅次于CSB3组,且均显著高于梭子蟹单养组(P<0.05)。而饲料能转化效率、单位净产量耗饲料能和单位净产量耗总能等指标则与梭子蟹单养对照相差不大(P>0.05);而CSB3处理虽然在饲料能转化效率、单位净产量耗饲料能和单位净产量耗总能等方面显著低于单养对照(P<0.05),但其光合能利用率在各处理组中最高而且总能量利用率也显著高于单养对照组(P<0.05),总产出量则显著高于其它处理(P<0.05)。因此,CSB3和CSB4处理为本研究各处理中的最优处理,即最佳三元混养结构,其中三疣梭子蟹、凡纳滨对虾和菲律宾蛤仔放养密度分别为45、6和30~60ind/m2。不过,本研究中蟹虾贝三元混养结构优化是在课题组蟹虾二元混养结构优化[41]的结果基础上进行的,而且菲律宾蛤仔放养密度最高仅为60ind/m2,因此,蟹、虾、贝混养是否还有更好的搭配方式和比例,可能还需进一步深入研究。
[1]王大鹏,韦嫔媛.对虾池混养的生态学原理及现状 [J].广西水产科技,2008(1):36-40.
[2]刘从力,刘世禄.我国海水养殖业发展现状与可持续发展问题[J].黄渤海海洋,2001,19(3):100-105.
[3]黄晓,杨延保等.养殖对虾病害生态防治的探讨 [J].生态科学,1999,18(2):24-30.
[4]张哲,穆峰等.太平洋牡蛎(Crassostreagigas)对虾池水质的影响[J].中国海洋大学学报,2002,32(增刊):24-29.
[5]Prince T C.Bivalve grazing,nutrient cycling and phytoplankton dynamics in an estuarine ecosystem[D].Wageningen:University of Wageningen,1996:151.
[6]Black K S,David D M.Measurement of the erosion potential of cohesive marine sediment:A review of current in situ technology[J].Journal of Marine Environmental Engineering,1997,4:43-83.
[7]Widdows J,Brinsley M D.A Benthic Annular Flume for In Situ Measurement of Suspension Feeding/Biodeposition Rates and Erosion Potential of Intertidal Cohesive Sediments [J].Estuarine,Coastal and Shelf Science,1998,46:27-38.
[8]Haven D S and Morales-Alamo R.Bio-deposition as a factor in sedimentation of fine suspended solids in estuaries[J].Geological society of America,Memoir,1972,133:121-130.
[9]Hatcher A,Grant J,Schofield B.Effects of suspended mussel culture(Mytilusspp.)on sedimentation,benthic respiration and sediment nutrient dynamics in a coastal bay [J].Marine ecology process series,1994,115:219-235.
[10]宋颀,田相利,董双林,等.草鱼混养生态系统能量收支的研究[J].中国海洋大学学报:自然科学版,2011,41(10):45-51.
[11]Li S.Energy structure and efficiency in a typical Chinese integrated fish farm [J].Aquaculture,1987,65:105-118.
[12]雷惠僧,谭玉钧,葛光华.河埒水产养殖综合养鱼复合生态系统的初步研究 [J].水产科技情报,1983(3):10-14.
[13]李吉方,董双林,文良印.盐碱地池塘不同养殖模式的能量利用比较 [J].中国水产科学,2003,10(2):143-147.
[14]包杰,田相利,董双林,等.对虾、青蛤和江蓠混养的能量收支及转化效率研究 [J].中国海洋大学学报:自然科学版,2006,36:27-32.
[15]董贯仓,田相利,董双林,等.几种虾、贝、藻混养模式能量收支及转化效率的研究 [J].中国海洋大学学报:自然科学版,2007,37(6):899-906.
[16]Balasubramnian S,Pappathi R,Raj S P.An energy budget and efficiency of sewage-feed fish ponds[J].Bioresourses Technology,1995,52:145-150.
[17]周一兵,刘亚军.虾池生态系能量收支和流动的初步分析 [J].生态学报,2000,20(5):474-481.
[18]翟雪梅,张志南.虾池生态系能流结构分析 [J].青岛海洋大学学报,1998,28(2):275-282.
[19]游奎.对虾工程化养殖系统重要元素及能量收支 [D].青岛:中国科学院研究生院,2005.
[20]田相利,董双林,王芳.不同温度对中国对虾生长及能量收支的影响 [J].应用生态学报,2004,15(4):678-682.
[21]张继红,方建光,金显仕,等.低温对菲律宾蛤仔能量收支的影响 [J].水产学报,2002,26(5):423-427.
[22]张硕,董双林.饵料和盐度对中国对虾幼虾能量收支的影响 [J].大连水产学院学报,2002,17(3):227-233.
[23]王兴强,曹梅,马甡,等.密度对凡纳滨对虾存活生长和能量收支的影响 [J].中国农学通报,2005,21(8):409-412.
[24]裴宇.凡纳滨对虾与金钱鱼混养对池塘生态因子及能量收支的影响 [D].湛江:广东海洋大学,2011.
[25]李德尚,杨红生.一种池塘陆基实验围隔 [J].青岛海洋大学学报,1998,28(2):199-204.
[26]王骥.浮游植物的初级生产力与黑白瓶测氧法 [J].淡水渔业1980,3:24-28.
[27]张天文.对虾高位池精养模式和生态养殖模式中碳流通特征的解析一Ecopath with Ecosim在养殖生态系统中应用的初步研究[D].青岛:中国海洋大学,2011.
[28]刘思俭,曾淑芳.江蓠在不同水层中的光合作用与生长 [J].水产学报,1982,6(1):59-64.
[29]康春晓,雷慧僧,谭玉钧.以草鱼,鲢鱼为主养鱼的池塘能量转换效率初探 [J].水产科技情报,1990,2:47-49.
[30]吴乃薇,边文冀,姚宏禄.主养青鱼池塘生态系统能量转换率的研究 [J].应用生态学报,1992,3(4):333-338.
[31]周毅,杨红生,张福绥.海水双壳贝类的生物沉积及其生态效应[J].海洋科学,2003,27(1):23-26.
[32]Nakamura M,Yamamuro M,Ishikawa M,et al.Role of the bivalveCorbiculaJaponicain the nitrogen cycle in a mesohaline lagoon[J].Mar Biol,1988(99):369-374.
[33]Kautsky N,Evans S.Role of biodeposition byMytilusedulisin the circulation of matter and nutrients in a Baltic coastal ecosystem [J].Mar Ecol Prog Ser,1987(38):201-212.
[34]Kaspar H F,Gillespie P A,Boyer I C,et al.Effects of mussel aquaculture on the nitrogen cycle and benthic communities in Kenepru Sounds[J].New Zealand,Mar Biol,1985(85):127-136.
[35]Gilbert F,Souchu P,Bianchi M,et al.Influence of shellfish farming activities on nitrification,nitrate reduction to ammonium and denitrification at the water-sediment interface of the Thau lagoon,France[J].Mar Ecol Prog Ser,1997(151):143-153.
[36]项福亭,曲维功,张益额,等.庙岛海峡以东浅海养殖结构调整的研究 [J].齐鲁渔业,1996,13(2):1-4.
[37]souchu P,vaquer A,collos Y,et al.Influence of shellfish farming activities on the biogeochemical composition of the water column in Thau lagoon[J].Mar Ecol Prog Ser,2001,218:141-152.
[38]Dame R T,Libes S.Oyster reefs and nutrient retention in tidal creeks.Journal of Experimental[J].Marine Biology and Ecology,1993,171(2):251-258.
[39]董双林,潘克厚,Uwe Brockmann.海水养殖对沿岸生态环境影响研究的进展 [J].青岛海洋大学学报,2000,30(4):575-582.
[40]田相利,李德尚,阎希翥,等.对虾池封闭式三元综合养殖的实验研究 [J].中国水产科学,1999,6(4):49-54.
[41]董佳.三疣梭子蟹(Portunustrituberculatus)池塘混养系统结构优化和氮磷收支的实验研究 [D].青岛:中国海洋大学,2013.
