乒乓球运动员的大脑功能非对称性及功能耦合性研究
2015-11-01郭志平李安民王积福
郭志平,李安民,王积福
大脑半球特化是人类大脑组织的主要特征之一,左半球主要负责言语功能,右半球主要负责空间信息加工[1],这种大脑组织特征是否会受到经验的影响及其机制问题受到研究者的广泛关注,运动员群体为了解经验对大脑功能组织特性的影响提供了良好的契机。
在体育运动,特别是对抗性的运动项目中,运动员通常需要对瞬息万变的运动情景进行快速处理,继而迅速地做出恰当的反应,因此视觉空间信息的处理能力对运动员而言至关重要。早期的生理研究表明,经过长期的运动训练,运动员右半球视觉-空间能力提高,他们的大脑功能非对称性变得越发显著[2]。随着科学技术的进步和研究方法的革新,脑电技术被应用于运动员大脑功能特性的研究中,其中不少是以脑电特定频段能量变化或相干性为指标的。通常,alpha频段(8~13 Hz)能量的增加反映了皮层激活水平的降低[3]。在认知或注意任务中,alpha频段能量的增加往往在任务无关皮层区域出现,与半球内抑制相联系[4-6],而左半球颞叶alpha频段能量的上升可能表明左半球言语认知加工的抑制。theta频段(4~7 Hz)反映了通过远距离皮层交流进行的神经环路整合,以及自上而下的注意加工过程[7]。
对射击、射箭以及高尔夫选手的研究表明,运动员在自主运动动作的准备期间,与大脑右半球相比,左半球(主要是颞叶区域)显示出alpha频段能量的上升[8-12]。与低水平运动员相比,高水平运动员在动作准备、动作识别和动作表象过程中显示出更高的左半球alpha频段能量[9,13-14]。但是,这种运动员左侧大脑alpha频段能量更高的结论与另一些研究结果相矛盾。如有研究发现,运动成绩与双侧或主要是与右侧alpha频段的高能量相联系[15-17]。此外,对运动动作的识别判断以及手部自主运动的脑电研究表明,运动员比非运动员的alpha频段事件相关去同步化幅值更低,即能量更高[18-19]。
除了比较alpha频段的能量变化,研究者也运用脑电相干性分析方法对运动员的大脑功能组织特性进行探索。研究发现,与低水平选手相比,高水平选手在动作准备[20]、动作表象[13]和动作识别[21]时表现出左侧大脑的低相干性,特别是左侧颞叶(T3)和运动前区(FZ)之间。但是,并非所有研究都支持运动员左侧大脑具有低相干性的结论。如C.DEL PERCIO等[22]采用脑电相干性分析方法对气手枪选手的研究表明,在准备射击期间,高水平选手的双侧顶颞叶、顶枕叶脑区低频alpha、高频alpha频段的相干性高于非运动员。此外,C.BABILONI等[23]对高尔夫专家选手的相干分析发现,与失败推杆相比,成功推杆时在双侧顶额叶以及顶中央脑区的半球内低频alpha相干性更高。
总之,已有研究丰富了对运动经验之于大脑功能特性影响的认识,然而回顾前人的研究发现,关于运动员在任务完成过程中特定脑电频段能量变化或是脑电相干性高低,结论并不一致。此外,现有研究均是针对运动员在动作准备、动作识别或动作表象时的大脑功能特性,鲜有研究探讨运动员在运动经验相关以及经验无关的一般认知过程中的大脑功能特性,正如M.W.VOSS等[24]有关运动与认知关系的元分析所指出的那样,未来研究应该使用高水平的认知任务和不同的视觉注意测试。因此,本研究运用脑电技术,以经验相关和经验无关图形作为刺激材料,对乒乓球运动员在图形识别过程中的大脑功能非对称性及耦合性进行研究,以探讨下面2个问题:(1)运动经验与脑功能非对称性之间的关系;(2)乒乓球运动员在经验相关和无关图形识别过程中不同脑区之间的功能耦合,研究假设如下。(1)与非运动员相比,乒乓球运动员在经验相关图形识别过程中显示出更高的右侧颞叶激活,更少的左侧颞叶激活;在经验无关图形识别过程中无差异。差异通过左右颞叶8~13 Hz alpha能量差值(T8减去T7 alpha能量)反映,差值为正则表示左侧颞叶激活高于右侧,差值为负责表示右侧颞叶激活高于左侧,这也作为运动员在经验相关图形识别过程中言语分析多少的反映指标。(2)与非运动员相比,乒乓球运动员在经验相关图形识别过程中显示出更高的T8FZtheta(4~7 Hz)相干,更低的T7FZtheta相干;在经验无关图形识别过程中无差异。暗示乒乓球运动员更多地依赖于视觉-空间动作加工,而非言语分析加工,并且这种特性仅限于对运动经验相关图形的识别。
1 研究对象与方法
1.1 被试
共有28名男性右利手被试参加本试验,其中来自上海体育学院乒乓球训练专业的乒乓球运动员14名,运动等级为二级及以上,平均年龄为(19.64±1.50)岁,平均训练年限为(11.00±2.50)年;其余14名为上海体育学院运动科学学院、经济管理学院和体育新闻传播与外语学院的普通大学生,平均年龄为(21.50±1.83)岁,均没有乒乓球运动的训练经历。在试验前,被试熟悉本研究的目的、试验要求、方法及流程,自愿参加试验并签署知情同意书,试验完成后给予一定的报酬。
1.2 试验设备
用的E-Prime2.0心理试验编程软件编制和播放试验程序。德国Brain Products公司生产的64导事件相关电位测试系统,以及用于运行试验程序,记录和采集行为、脑电数据的2台DELL计算机,计算机的分辨率为1 024×768,刷新率为100 Hz。
1.3 试验材料
“Type Token”模型[25]认为,对于熟悉的物体,个体会形成特殊的表征形式,包括“Type”和“Token”2种分离的表征水平。“Type”提供物体的原型信息,主要包括轮廓(outline)和三维(three-dimension)信息。“Token”储存物体的朝向(orientation)和颜色(color)信息,并且可以与“Type”进行捆绑(bind)储存。如果长时间接触某物体,个体就能将“Type”和“Token”信息简化保存,形成捆绑式的特殊表征形式。对于乒乓球运动员来说,迅速判断出击球点位置并立即做出恰当的反应是极其关键的。本研究依据“Type Token”模型和乒乓球项目特点设计了2类刺激材料。(1)运动经验相关刺激材料——带缺口的圆,圆形提供乒乓球的轮廓信息,而缺口朝向则提供击球点方向的信息。圆的缺口角度为15°,缺口位置有4种,按顺时针方向计算,第1个缺口的正中位置对应45°(右上方),其余缺口的正中位置依次对应135°、225°和315°。该刺激的轮廓与朝向组合对运动员来说是一种运动经验相关的刺激物,可以用来启动经验相关认知加工过程[26]。(2)运动经验无关刺激材料——带缺口的十字星,该刺激材料虽然在朝向和轮廓捆绑方式上与经验相关刺激物相同,但在轮廓上与乒乓球运动专项情境无关,以此破坏已建立好的“Type Token”捆绑模式,对乒乓球运动员来说是一种经验无关的一般性刺激物。
1.4 试验程序
在某高校运动心理研究中心脑电实验室完成试验,实验室环境舒适、隔音,且光线柔和。试验前,被试需要洗干净头发并吹干,熟悉实验室的环境,填写被试基本情况调查表,关闭手机及其他便携式通讯设备。
试验是在只有主试和被试的独立环境中进行的。开始前,向被试介绍试验要求:选舒服的坐姿坐好并在试验过程中尽量使全身放松;将头放在面前的U型托上以保持头部稳定,U型托离显示器95 cm,双眼与显示器屏幕的中心处在同一条水平线上,在正式开始试验后,需要尽量保持头部和身体稳定;将双手舒服地搁置在小键盘的数字键上,并告知被试屏幕上将呈现的试验指导语以及需要进行的简单按键操作反应。
试验程序设置了练习和正式测试2个阶段。为了消除操作熟练性因素对试验结果的影响,在正式开始试验前,让被试进行一定次数的练习,当被试的按键反应时趋于稳定后进入正式试验。
试验任务为选择反应任务,刺激呈现时会出现包含轮廓类型相同,但缺口方向不同的4个图形的图片,靶刺激为缺口正中位置对应135°(右下方)的圆或十字星,靶刺激只会是4个图形中的一个,不会同时出现2个、3个或4个靶刺激,其他位置缺口的圆或十字星为非靶刺激。在试验中,被试需要又快又准地辨别所呈现出来的刺激中是否包含靶刺激,如果出现了包含圆靶刺激的图片,便使用右手食指按“1”键进行反应,如果出现的是包含十字星靶刺激的图片,则用右手中指按“3”键进行反应,非靶刺激不作反应。圆、十字星靶刺激,以及圆、十字星非靶刺激各呈现72次,共288次,呈现方式是随机的,试验时间大约为 15 min(见图 1)。
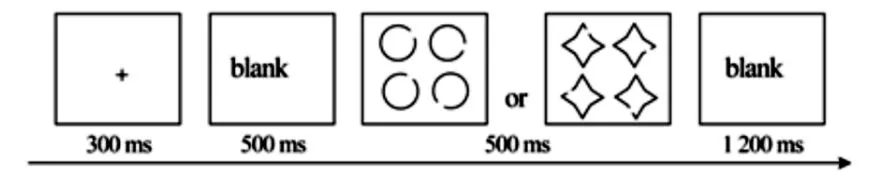
图1 试验任务流程图
1.5 脑电的记录
使用德国Brain Product的事件相关电位记录系统记录脑电数据(带宽0.01~100 Hz,采样率500 Hz),采用国际标准10-20扩展系统64导电极帽。在AFz和Fz电极点之间安放电极参考点,在Pz和Oz之间安放接地参考点,在右眼外侧和左眼下方分别安放水平眼电(HEOG)和垂直眼电(VEOG)电极点,电极阻抗保持在5 kΩ以下。
1.6 数据的处理与分析
试验获得的行为数据(反应时、反应正确率)采用E-prime 2.0软件记录并进行初步的筛选和处理,而后运用spss16.0剔除超过3个标准差的数据,再对不同组别被试的反应时、正确率进行重复测量方差分析。
脑电数据的处理采用的是脑电数据分析软件Analyzer 2.0,使用双耳乳突的平均电位作为参考,去除眨眼伪迹,排除噪音干扰以及其他伪迹,滤波范围设置为0.01~35 Hz,去除50 Hz市电干扰,将波幅大于的当作伪迹自动去除。用于提取alpha频段能量和计算theta频段相干性的时间窗口为0~900 ms,刺激呈现时间点为0 ms。剔除7名脑电伪迹较多的被试,最终纳入统计分析的运动员组和非运动员组各14名被试。
为了检验前面的假设(在经验相关图形识别过程中,与非运动员相比,乒乓球运动员显示出更高的右侧颞叶激活,更少的左侧颞叶激活,T8-FZtheta相干性高,T7-FZtheta相干性低,但二者在经验无关图形识别过程中无上述差异),采用重复测量方差分析对alpha频段能量、theta频段相干性数据进行分析。在分析过程中,数据的方差齐性评估采用Mauchlcy球型检验,对于不满足球型检验的统计量运用Greenhouse-Geisser法进行自由度的矫正。
在评估左右半球8~13 Hzalpha频段能量差异时,以T8-T7 alpha能量差值为因变量,以组别(运动员、非运动员)为组间变量,刺激类型(圆、十字星)为组内变量,对alpha频段能量的差异进行2(组别)×2(刺激类型)的重复测量方差分析。在评估左右半球的功能耦合情况时,以组别(运动员、非运动员)为组间变量,以脑区(T7-FZ、T8-FZ)为组内变量,分别对圆和十字星识别过程中的脑电相干性进行2(组别)×2(脑区)的重复测量方差分析。
2 结 果
2.1 行为数据结果
对正确率的方差分析结果显示,组别和刺激类型的交互效应、组别的主效应均不显著(P>0.09),刺激类型的主效应显著(P<0.01),圆的正确率显著高于十字星。对反应时的方差分析结果则表明,组别与刺激类型的交互效应显著(F(1,26)=4.917,P=0.036,η2=0.159)。随后进行的简单效应分析发现,在对圆的反应中,运动员的反应时要显著小于非运动员(P=0.039),此外,运动员对圆的反应显著快于十字星(P=0.001)(见表1)。表明,运动员在经验相关图形识别过程中具有速度优势,但是这种优势并未迁移到经验无关的图形识别中。
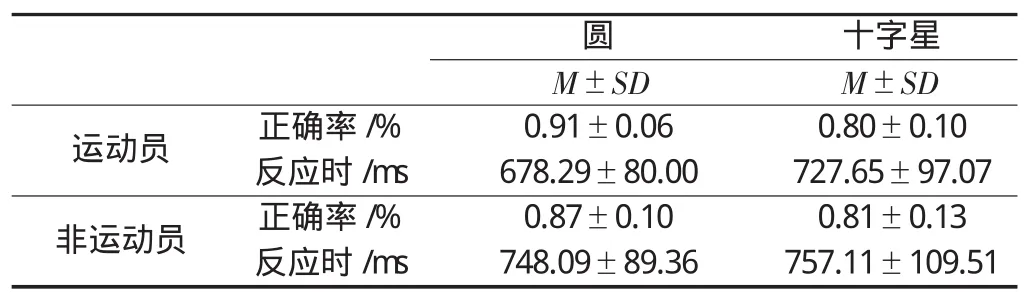
表1 运动员与非运动员在图形识别过程中的正确率和反应时(n=14)
2.2 脑电结果
2.2.1 运动员与非运动员在图形识别过程中的alpha频段能量对T8-T7alpha频段的能量差异进行的方差分析结果表明,组别和刺激类型的交互效应显著(F(1,26)=4.524,P=0.043,η2=0.148)(见图2)。简单效应分析发现,运动员在对圆(-0.12±0.65)进行反应时的左右侧颞叶alpha频段的能量差值显著大于十字星(-0.91±0.98)(P=0.031),非运动员则无上述差异(-0.93±1.23圆,-0.68±1.39十字星,P>0.05);在对圆进行识别时,运动员(-0.12±0.65)的左右颞叶alpha频段能量差值显著大于非运动员(-0.93±1.23)(P=0.039),在对十字星的反应中,alpha频段能量没有显示出组间差异(-0.91±0.98运动员,-0.68±1.39非运动员,P>0.05)。此外,鉴于T8-T7的差值为负,表明运动员与非运动员在图形识别过程中均为右侧颞叶激活比左侧高,在大脑半球的非对称性程度上,运动员要低于非运动员。

图2 运动员与非运动员左右颞叶alpha频段能量差
2.2.2 运动员与非运动员在图形识别过程中的脑电相干性 运动员与非运动员在圆形识别过程中脑电相干性重复测量方差分析显示,组别与脑区的交互效应显著(F(1,26)=11.247,P=0.002,η2=0.302)(见图3)。简单效应分析发现,运动员的右半球(0.23±0.19)相干性显著高于左半球(0.04±0.06)(P<0.000),非运动员左右半球相干性无差异(0.07±0.11右,0.10±0.10左,P>0.05);在右半球相干性上,运动员(0.23±0.19)显著高于非运动员(0.07±0.11)(P=0.019),而左半球相干性无组间差异(0.04±0.06运动员,0.10±0.10非运动员,P>0.05)。对十字星形识别过程中的脑电相干性不存在组间及半球差异(0.05±0.10运动员右,0.07±0.12非运动员右,0.10±0.22运动员左,0.12±0.25非运动员左,P>0.05)。
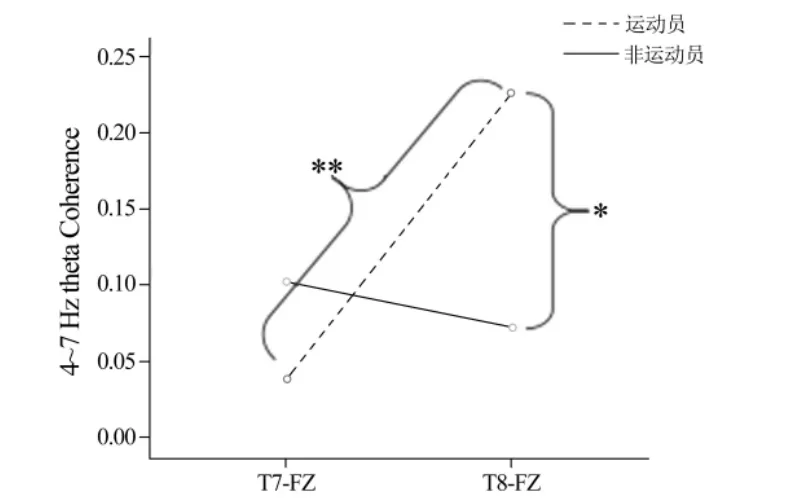
图3 运动员和非运动员左右颞叶与运动前区的theta频段相干性
3 讨 论
运动前区与运动区、颞叶与顶叶皮层之间有着广泛的生理连接[27],在心理运动任务以及动作技能学习中,这些连接可能反映了动作计划和躯体感觉以及视觉运动整合[28-29]。大脑颞叶功能非对称性以及颞叶与运动前区之间的信息交流可能与运动员的动作效率有关[13]。
alpha频段(8~13 Hz)反映了与高水平运动员熟练认知-运动表现相联系的皮层神经同步化机制,是了解专家大脑功能组织特征的窗口[9,19,22]。alpha频段能量的增加被认为是皮层激活降低的指标[3],反映了加工抑制、选择性皮层信息加工或皮层空闲[30],而alpha频段能量的降低是皮层激活上升的反映,表示皮层信息加工。
在alpha频段能量的变化上,与非运动员相比,假设运动员在经验相关图形识别中显示出更高的右侧颞叶激活,更少的左侧颞叶激活;在经验无关图形的识别过程中无差异。研究结果与假设相反,运动员在对圆的反应中,左右颞叶alpha频段能量差显著大于非运动员,而且运动员在对圆进行反应时的左右颞叶alpha频段能量差显著大于十字星;在对十字星的识别过程中,无组间及半球差异,与假设一致。即与非运动员相比,运动员的皮层激活以及左右半球的非对称程度均较低,而且这种差异只出现在经验相关图形识别中。该结果得到了相关研究[15-17]的支持,运动成绩与双侧或主要是与右侧alpha频段的高能量相联系。此外,对体操动作的识别研究表明,体操运动员的右侧颞叶和枕叶区域高频alpha频段能量在体操动作识别中更高[18],研究者认为,这种alpha频段高能量(低激活)是专家大脑神经效率加工的体现,即在完成相同任务过程中,专家大脑的皮层激活可能具有空间集中特性[31],只需耗费相对较少的皮层资源。乒乓球运动员通过常年的运动训练,对乒乓球及其击球点位置的判断已经达到非常熟练的程度,可能形成捆绑式的表征方式,在面对相似情景(缺口圆的判断)的时候,是基于感觉信息与长时记忆中的信息进行比较,并与记忆中的模式进行匹配,从而做出判断。该过程的完成对于运动员而言几乎是自动化的,速度快,而且耗费的皮层资源相对较少。因此,乒乓球运动员对圆的识别反应速度相对更快,皮层激活水平以及左右大脑半球的非对称程度均较低,右侧激活高于左侧,而且这种差异只出现在经验相关图形识别中,可能源于运动员具有良好的视觉-空间注意技巧[12]。在经验相关图形识别过程中,只需相对较少的皮层资源,主要是右半球的视觉空间资源,支持了专家大脑的神经效率假说[32]。此外,较低的半球非对称性可能反映了运动员具有更好的认知功能,具体表现在操作思维、认知和信息加工方面[33]。
脑电相干性被认为是对脑功能连接所必须的网络构成和整合的测量[34],低脑电相干性与脑血流量降低有关[35],高相干性与信息交换[36]以及脑区之间的功能耦合、功能协调性[37-38]有关。在脑电相干性上,与非运动员相比,假设乒乓球运动员在经验相关图形识别过程中显示出更高的T8FZtheta相干,更低的T7FZtheta相干;在经验无关图形的识别过程中,无上述差异。研究结果与假设一致,在对圆进行识别时,运动员的右半球相干性显著高于左半球,非运动员左右半球相干性无差异;在右半球相干性上,运动员显著高于非运动员,而左半球相干性无组间差异;对十字星进行识别时,脑电相干性不存在组间及半球差异。该结果得到有关射击、高尔夫研究[22-23]的支持,研究者认为,运动员在任务完成过程中的高相干性反映了其良好的大脑功能耦合和更有效的大脑皮质功能。鉴于乒乓球运动员长时间接触乒乓球,可能将形状(圆形)和方向(缺口方向)这2个水平的信息进行了简化保存,形成了捆绑式的特殊表征,结合theta频段的功能含义,即它反映了通过远距离皮层交流进行的神经环路整合以及自上而下的注意加工过程[4,7]。运动员在圆的识别过程中,右侧颞叶与运动前区之间显示出高相干性,可能表明了乒乓球运动员是依赖于已建立好的内部模式来调节信息的匹配和执行,相关脑区具有良好的功能耦合,各脑区能更加协同一致地活动,启动了自上而下的加工,具有更加有效的大脑皮质功能。而非运动员颞叶与运动前区之间的低相干性可能源于他们对形状方向信息没有形成捆绑式的内部记忆表征,相关脑区的活动还不够协调一致,需要更多地依赖环境中的信息线索,启动了自下而上的加工,而不是启动记忆来应对当前的任务。此外,运动员的高相干性只是显示在右侧脑区而非左侧脑区、经验相关而非经验无关图形识别过程中,结合大脑功能组织特征,即左半球主要负责言语功能而右半球主要负责空间信息加工[1],左侧颞叶主要与言语分析和语言功能相联系,而右侧颞叶与视觉-空间和整合加工相联系[39]。此研究结果可能表明,在图形识别过程中,乒乓球运动员比非运动员更多地依赖于视觉-空间动作加工,而非言语分析加工,并且上述特性并未迁移到经验无关图形的识别过程中。
总体上看,无论是行为还是脑电指标都表明,运动员的优势反应仅限于对运动经验相关图形的识别,该结果与“狭窄迁移假说”(narrow transfer hypothesis)的观点一致,即特定领域的专家在其专长领域内具有优异的认知加工能力,但在其专长领域外则不尽然[40]。
4 结 论
运动员的反应速度快于非运动员,运动员右侧颞叶激活以及颞叶-运动前区之间的相干性显著高于左侧,与非运动员相比,运动员在图形识别过程中显示出较低的右侧颞叶激活,较高的T8FZtheta相干,并且上述差异只出现在经验相关图形识别中。该结果可能表明,运动员的反应更快,他们拥有良好的视觉-空间注意技巧,在经验相关图形识别过程中更多地依赖于视觉-空间加工,而非言语分析加工,需要的皮层资源相对较少,具有信息加工的神经高效性,而且上述特性不具有迁移性。
[1]HERVE P Y,ZAGO L,PETIT L,etal.Revisiting human hemispheric specializationwith neuroimaging[J].Trends Cogn Sci,2013,17(2):69-80.
[2]NIKOLAENKO N N,MIKHEEV M M,AFANAS'EV S V.Changes of motor and sensory asymmetries in highly trained athletes[J].Journal of Evolutionary Biochemistry and Physiology,2001,37(3):273-279.
[3]ALLEN JJ,COAN J A,NAZARIAN M.Issues and assumptions on the road from raw signals to metrics of frontal EEGasymmetry in emotion[J].Biol Psychol,2004,67(1-2):183-218.
[4]ASTRID VON STEIN,SARNTHEIN J.Different frequencies for different scales of cortical integration:from local gamma to long range alpha/theta synchronization[J].International Journal of Psychophysiology,2000,38(2000):301-313.
[5]KLIMESCH W.Evoked alpha and early access to the knowledge system:the P1 inhibition timinghypothesis[J].Brain Res,2011,1408:52-71.
[6]KLIMESCH W.alpha-band oscillations,attention,and controlled access tostored information[J].Trends Cogn Sci,2012,16(12):606-617.
[7]SAUSENG P,KLIMESCH W,SCHABUSM,etal.Fronto-parietal EEG coherence in theta and upper alpha reflect central executive functions of workingmemory[J].Int JPsychophysiol,2005,57(2):97-103.
[8]HILLMANCH,JANELLE CM,APPARIESRJ,etal.An electrocortical comparison of executed and rejected shots in skilled marksmen[J].Biological Psychology,2000,52:71-83.
[9]KERICK S E,DOUGLASS L W,HATFIELD B D.Cerebral cortical adaptationsassociated with visuomotor practice[J].Med Sci Sports Exerc,2004,36(1):118-129.
[10]JANELLE C M,HILLMAN C H,APPARIES R J,etal.Expertise Differences in Cortical Activation and Gaze Behavior During Rifle Shooting[J].Journal of Sport&Exercise Psychology,2000,22(2):167-182.
[11]LOZE G M,COLLINS D,HOLMES P S.Pre-shot EEG alpha-power reactivity during expert air-pistol shooting:a comparison of best and worstshots[J].JSports Sci,2001,19(9):727-733.
[12]SALAZAR W,LANDERSD M,PETRUZZELLO SJ,etal.Hemispheric asymmetry,cardiac response,and performancein elite archers[J].Res Q Exerc Sport,1990,61(4):351-359.
[13]WOLFS,BROLZE,KEUNEPM,etal.Motorskillfailureorflow-experience?Functional brain asymmetry and brain connectivity in elite and amateur tabletennisplayers[J].Biol Psychol,2015,105:95-105.
[14]HAUFLER A J,SANTA MARIA D L,SPALDING T W,etal.Neurocognitiveactivity duringaself-paced visuospatial task:comparative EEG profiles in marksmen and novice shooters[J].Biological Psychology,2000,53:131-160.
[15]BABILONIC,DELPERCIOC,IACOBONIM,etal.Golf putt outcomes are predicted by sensorimotor cerebral EEG rhythms[J].J Physiol,2008,586(1):131-139.
[16]DEL PERCIO C,BRANCUCCI A,BERGAMI F,etal.Cortical alpha rhythms are correlated with body sway during quiet open-eyes standing in athletes:a high-resolution EEGstudy[J].Neuroimage,2007,36(3):822-829.
[17]CREWS D J,LANDERS D M.Electroencephalographic measures of attentional patternsprior tothegolf putt[J].Medicine&Science in Sports&Exercise,1993,25(1):116-126.
[18]BABILONIC,DEL PERCIOC,ROSSINIPM,etal.Judgment of actions in experts:ahigh-resolution EEGstudy in elite athletes[J].Neuroimage,2009,45(2):512-521.
[19]BABILONIC,MARZANON,INFARINATOF,etal.“Neural efficiency”of experts'brain during judgment of actions:a high-resolution EEG study in elite and amateur karateathletes[J].Behav Brain Res,2010,207(2):466-475.
[20]DEENY S P,HILLMAN C H,JANELLE C M,etal.Cortico-cortical communication and superior performance in skilled marksmen:an EEG coherence analysis[J].Journal of Sport&Exercise Psychology,2003,25:188-204.
[21]王丽岩,李安民,王洪彪.乒乓球运动员动作识别时的脑电相干性分析[J].体育科学,2013,33(05):31-40.
[22]DEL PERCIO C,IACOBONI M,LIZIO R,etal.Functional coupling of parietal alpha rhythms is enhanced in athletes before visuomotor performance:a coherence electroencephalographic study[J].Neuroscience,2011,175:198-211.
[23]DEL PERCIO C,INFARINATO F,MARZANO N,etal.Reactivity of alpha rhythms to eyes opening is lower in athletes than non-athletes:a high-resolution EEG study[J].Int J Psychophysiol,2011,82(3):240-247.
[24]VOSSM W,KRAMERA F,BASAK C,etal.Are expert athletes'expert'in thecognitivelaboratory?A meta-analytic review of cognition and sport expertise[J].Applied Cognitive Psychology,2010,24(6):812-826.
[25]ZIMMER H D,ECKER U K.Remembering perceptual features unequallybound in object and episodic tokens:Neural mechanismsand their electrophysiological correlates[J].Neurosci Biobehav Rev,2010,34(7):1066-1079.
[26]张大为.Go/Nogo任务下运动经验对抑制控制影响的电生理证据[D].上海:上海体育学院,2014.
[27]FRIEDERICIA D,GIERHANSM.Thelanguagenetwork[J].Curr Opin Neurobiol,2013,23(2):250-254.
[28]HANAKAWA T.Rostral premotor cortex as agateway between motor and cognitivenetworks[J].Neurosci Res,2011,70(2):144-154.
[29]HARDWICK R M,ROTTSCHY C,MIALL R C,etal.A quantitative meta-analysis and review of motor learning in the human brain[J].Neuroimage,2013,67:283-297.
[30]PFURTSCHELLER G,LOPES D A,SILVA F H.Event-related EEG/MEG synchronization and desynchronization:basic principles[J].Clinical Neurophysiology,1999,110:1842-1857.
[31]DEL PERCIOC,ROSSINIPM,MARZANON,etal.Is there a“neural efficiency”in athletes?A high-resolution EEG study[J].Neuroimage,2008,42(4):1544-1553.
[32]DEL PERCIO C,BABILONI C,BERTOLLO M,etal.Visuo-attentional and sensorimotor alpha rhythms are related to visuo-motor performance in athletes[J].Hum Brain Mapp,2009b,30(11):3527-3540.
[33]GEORGIY K,LESIA K.Functional Brain Asymmetry and Cognitive Functions in Elite Wrestlers[J].International Journal of Wrestling Science,2014,4(1):26-34.
[34]NUNEZ P L,WINGEIER B M,SILBERSTEIN R B.Spatial-temporal structures of human alpha rhythms:theory,microcurrent sources,multiscalemeasurements,and global bindingof local networks[J].Human Brain Mapping,2001,13:125-164.
[35]LEUCHTER A F,NUTON T F,COOK I A,etal.Changes in brain functional connectivity in alzheimer-type and multi-infarct dementia[J].Brain,1992,115:1543-1561.
[36]ANDREW C,PFURTSCHELLER G.Lack of bilateral coherence of post-movement central beta oscillations in the human electroencephalogram[J].Neuroscience Letters,1999,273:89-92.
[37]GERLOFF G,RICHARD J,HADLEY J,etal.Functional coupling and regional activation of human cortical motor areas during simple,internally paced and externally paced finger movements[J].Brain,1998,121:1513-1531.
[38]GEVINSA,SMITH M E,LEONG H,etal.Monitoring working memory load duringcomputer-based taskswith EEGpattern recognition methods[J].Human Factors,1998,40(1):71-91.
[39]PARK JL,FAIRWEATHER M M,DONALDSON D I.Making the case for mobile cognition:EEGand sports performance[J].Neurosci Biobehav Rev,2015,52:117-130.
[40]FURLEY P,MEMMERT D.Studyingcognitive adaptationsin the field of sport:broad or narrow transfer?A comment on Allen,Fioratou,and McGeorge(2011)[J].Perceptual and Motor Skills,2011,113(2):481-488.
