油污土壤中脂肪酶产生菌的筛选及产酶条件优化
2015-10-28曲威等
曲威等
摘要:为了获得脂肪酶产生菌,进而获得高酶活菌株,试验采集东营胜利油田被废油长期污染的土壤,通过罗丹明B平板法进行菌株分离筛选,并采用橄榄油乳化法测定脂肪酶酶活。经16S rRNA鉴定,确定为铜绿假单胞菌属,命名为Pseudomonas aeruginosa S8。并对该菌株的摇床培养产酶条件进行了初步研究,采用单因素试验和正交试验对S8菌株产脂肪酶的条件进行优化,得到最佳发酵培养基与条件为葡萄糖8 g/L,酵母粉8 g/L,硫酸铵6 g/L,花生油 20 g/L,起始pH 7.0,接种量9%,温度30 ℃,发酵培养时间72 h,为工业化生产提供了出发菌株。
关键词:脂肪酶;铜绿假单胞菌;筛选;鉴定;优化
中图分类号:TQ925+.6;Q556+.1 文献标识码:A 文章编号:0439-8114(2015)16-4002-05
DOI:10.14088/j.cnki.issn0439-8114.2015.16.044
Screening of Lipase-Producing Strain in Oil Field and Optimization
of the Lipase Producing Conditions
QU Wei,SHAO Lei,SONG Li-fen,XU Rong-xue
(Yantai Academy of China Agricultural University,Yantai 264670, Shandong. China)
Abstract:In order to obtain lipase-producing strain of the bacteria,thus obtain high activity strains,the strain was screened from long waste-oil-pollutedsoil near Shenglin oil field in Dongying through Rhodamine B plate method and the lipase activity was determined with olive oil emulsification method. A strain S8 producing lipase was screened, which was identified as Pseudomonas aeruginosa by the identification of 16S rRNA. The fermentation conditions of a lipase from S8 were determined by the single-factor experiment and orthogonal design. The result showed that the optimal medium composition for lipase production was as follows: glucose 8 g/L,yeast extract 8 g/L, (NH4)SO4 6 g/L, peanut oil 20 g/L. The optimal fermentation conditions were pH of 7.0, fermentation temperature of 30 ℃, inoculation amount of 9%, and fermentation time of 72 h, which provided foundation for the further study of lipase production fermentation.
Key words: lipase; pseudomonas aeruginosa; screening; identification; optimization
脂肪酶(lipase,EC 3.1.1.3)是一类广泛存在于微生物和动植物细胞中的水解酶,能在油水界面催化甘油三酯生成甘油及脂肪酸[1],因其在非水相条件下催化反应可逆,可用于催化醇解、酯化和酯交换反应[2]。生物来源的脂肪酶由于反应条件缓和,对环境污染小,成本低,被广泛应用于食品、制革、饲料、洗涤和油脂水解等工业领域[3,4],并且在生物柴油的制备、地沟油的回收利用等方面有不可替代的应用价值[5]。
在自然界中,脂肪酶主要存在于动物胰脏、植物种子和微生物中,由于微生物种类多、繁殖快,并且所产生的脂肪酶一般都是分泌性的胞外酶,比动物脂肪酶酶解作用的pH和温度范围更宽[6],适合于工业化大生产和获得高纯度样品,因此微生物脂肪酶是工业用脂肪酶的重要来源。
本研究从山东东营胜利油田附近被废油长期污染的土壤中成功筛选出一株产脂肪酶效果较好的菌株,经过鉴定,确定为铜绿假单胞菌,命名为Pseudomonas aeruginosa S8,并对其产酶的培养条件进行优化,得到该菌产酶的最佳发酵条件,以期为脂肪酶的生产提供优良酶源。
1 材料与方法
1.1 材料
土样:采自东营胜利油田被废油严重污染的土壤,共16份。
富集培养基:酵母粉2 g/L,橄榄油10 g/L,KH2PO4 3 g/L,Na2HPO4 1 g/L,MgSO4·7H2O 0.5 g/L,pH 7.0。
筛选培养基:酵母粉 2 g/L,橄榄油乳化液 20 g/L,(NH4)2SO4 2 g/L,NaCl 0.05 g/L,KH2PO4 0.15 g/L,Na2HPO4 0.35 g/L,琼脂 2 g/L,罗丹明 B 0.005 g/L(过滤除菌),pH 7.0。
橄榄油与聚乙烯醇(体积分数为2%)以1∶3(体积比)的比例混合,搅动乳化5 min,即成乳白色聚乙烯醇橄榄油乳化液[7]。
种子培养基:蛋白胨2 g/L,葡萄糖 2 g/L,酵母粉 2 g/L,NaCl 3 g/L,K2HPO4 1 g/L,pH 7.0。
发酵培养基:蛋白胨2 g/L,葡萄糖 2 g/L,(NH4)2SO4 1 g/L,MgSO4·7H2O 1 g/L,KH2PO4 1 g/L,pH 7.0。
碳源筛选培养基:酵母粉2 g/L,(NH4)2SO4 2 g/L,NaCl 0.05 g/L,KH2PO4 0.15 g/L,Na2HPO4 0.35 g/L,pH 7.0,分别加入蔗糖、葡萄糖、可溶性淀粉、乳糖、麦芽糖作为碳源。
氮源筛选培养基:葡萄糖 2 g/L,NaCl 0.05 g/L,KH2PO4 0.15 g/L,Na2HPO4 0.35 g/L,pH 7.0,分别加入酵母粉、蛋白胨、尿素、(NH4)2SO4、KNO3作为氮源。
1.2 试验方法
1.2.1 产脂肪酶菌种筛选
1)富集培养。每种土样称量5.0 g,加入20 mL无菌水,振荡制成土壤悬液。取每种土壤悬液各5 mL加入装有30 mL富集培养基的三角瓶中,将三角瓶置于30 ℃摇床振荡培养3 d。富集3次。
2)初筛培养。将过滤灭菌的罗丹明B加入到已灭菌的初筛培养基中,倒平板。从富集培养的菌液中取1 mL菌液,用去离子水进行系列梯度稀释后涂布在平板中,30 ℃培养3 d后,在350 nm紫外光下观察,产生脂肪酶的菌株周围会出现荧光圈,且这种荧光圈越大,脂肪酶活越高。依据培养平板上形成的荧光圈大小进行菌种筛选。
3)复筛培养。挑取初筛培养基上菌落周围变色圈大的菌落划线分纯。分离出单菌落后斜面保存,将分纯后的菌株转接到装有30 mL种子培养基的瓶中,30 ℃180 r/min培养24 h,然后以5% 的接种量将菌液加入到50 mL的发酵培养基中,30 ℃180 r/min培养72 h,离心得到上清液进行酶活测定。
1.2.2 酶活的测定 脂肪酶酶活的测定采用经典的聚乙烯醇橄榄油乳化液方法[8]。酶活力单位定义:30 ℃条件下,10 min油脂水解反应,每分钟催化脂肪水解产生1 μmol脂肪酸的脂肪酶量定义为一个脂肪酶国际单位(U/mL)。
1.2.3 菌株的分子生物学鉴定 菌株基因组DNA提取的方法参考文献[9]。16S rRNA PCR引物序列为F:5-AGAGTTTGATCCTGGCTCAG-3′,R:5′- GGTTACCTTGTTACGACTT -3[引物由上生工生物工程(上海)股份有限公司合成]。PCR扩增体系(50 μL):2 × PCR Taq M 25 μL;引物F(10 μmol/L)2 μL;引物R (10 μmol/L)2 μL;DNA模板4 μL;ddH2O 17 μL。PCR反应循环体系为:94 ℃ 预变性3 min;94 ℃变性1 min,50 ℃退火30 s,72 ℃延伸2.5 min,30个循环;72 ℃ 延伸10 min。反应结束后,取10 μL PCR产物用1%的琼脂糖凝胶电泳检测。将PCR产物送至生工生物工程(上海)股份有限公司测序,并将测得的序列拼接完成后登陆到NCBI BLAST中进行同源性比对。
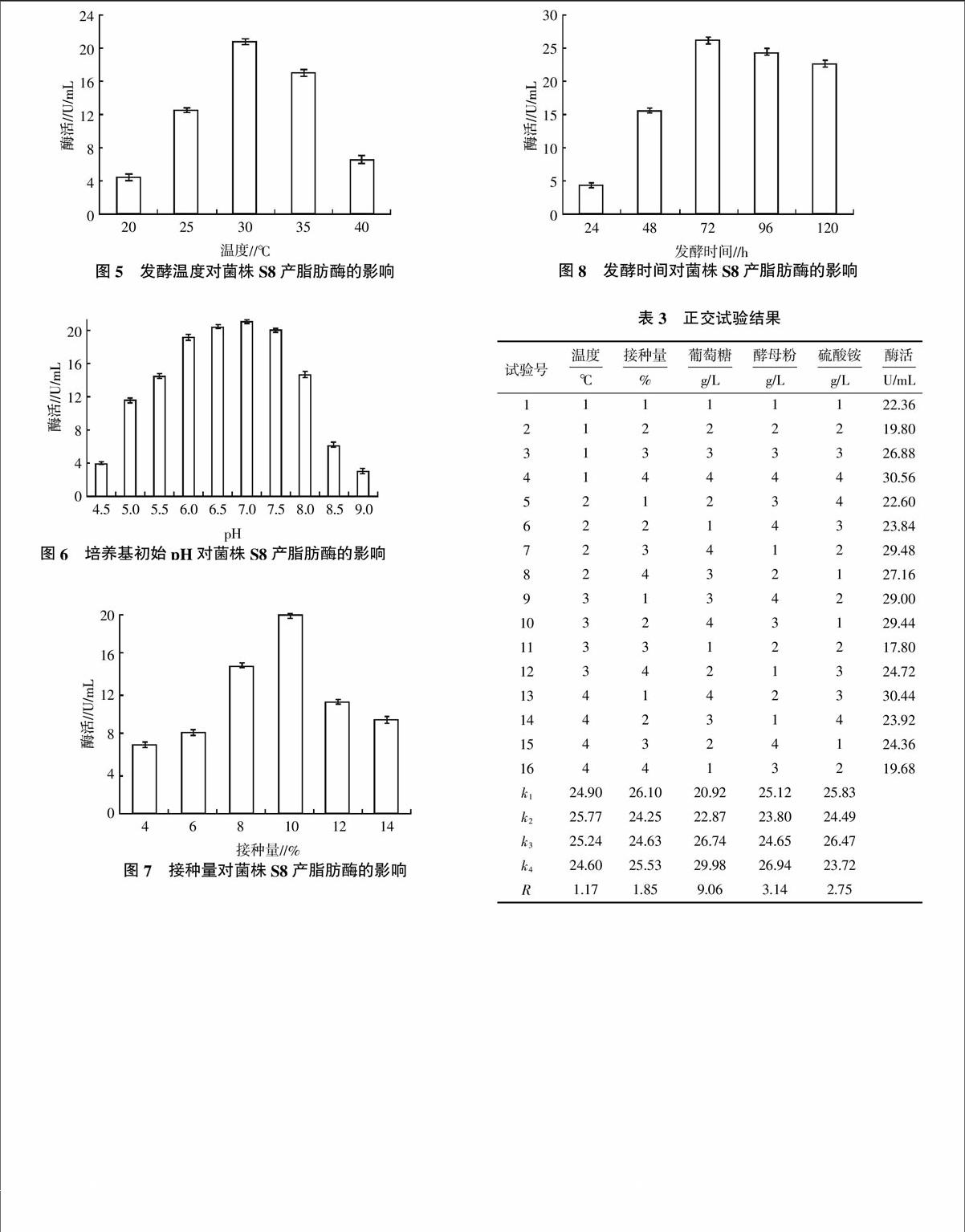
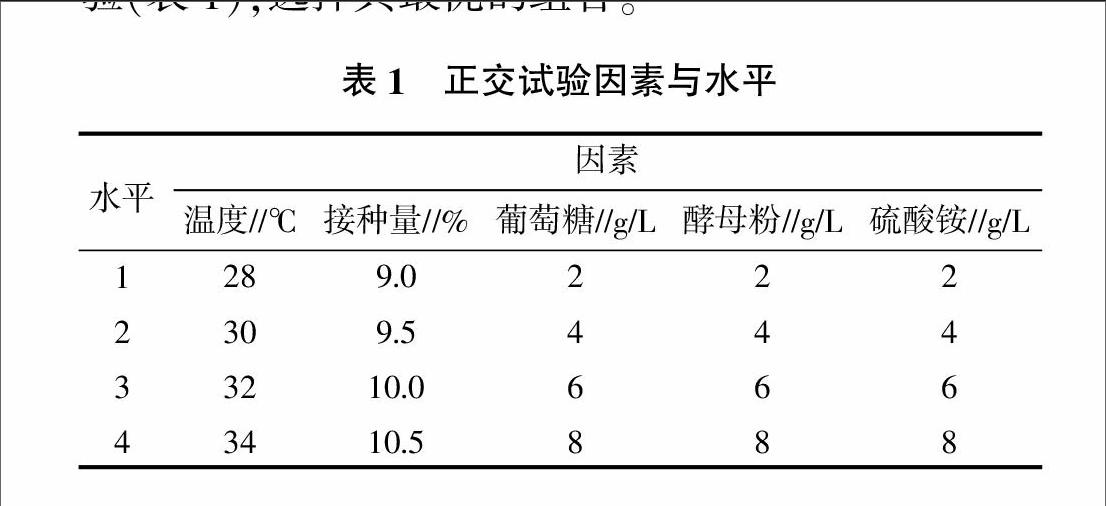
1.2.4 产酶条件参数优化 设计单因素试验,分别以培养基中的碳源、氮源、诱导剂和发酵条件中的温度、初始pH、接种量、发酵时间为因素,考察发酵条件对产酶的影响,选择对产脂肪酶菌株影响较大的5个因素,对其进行5因素4水平的正交试验(表1),选择其最优的组合。
2 结果与分析
2.1 菌种筛选
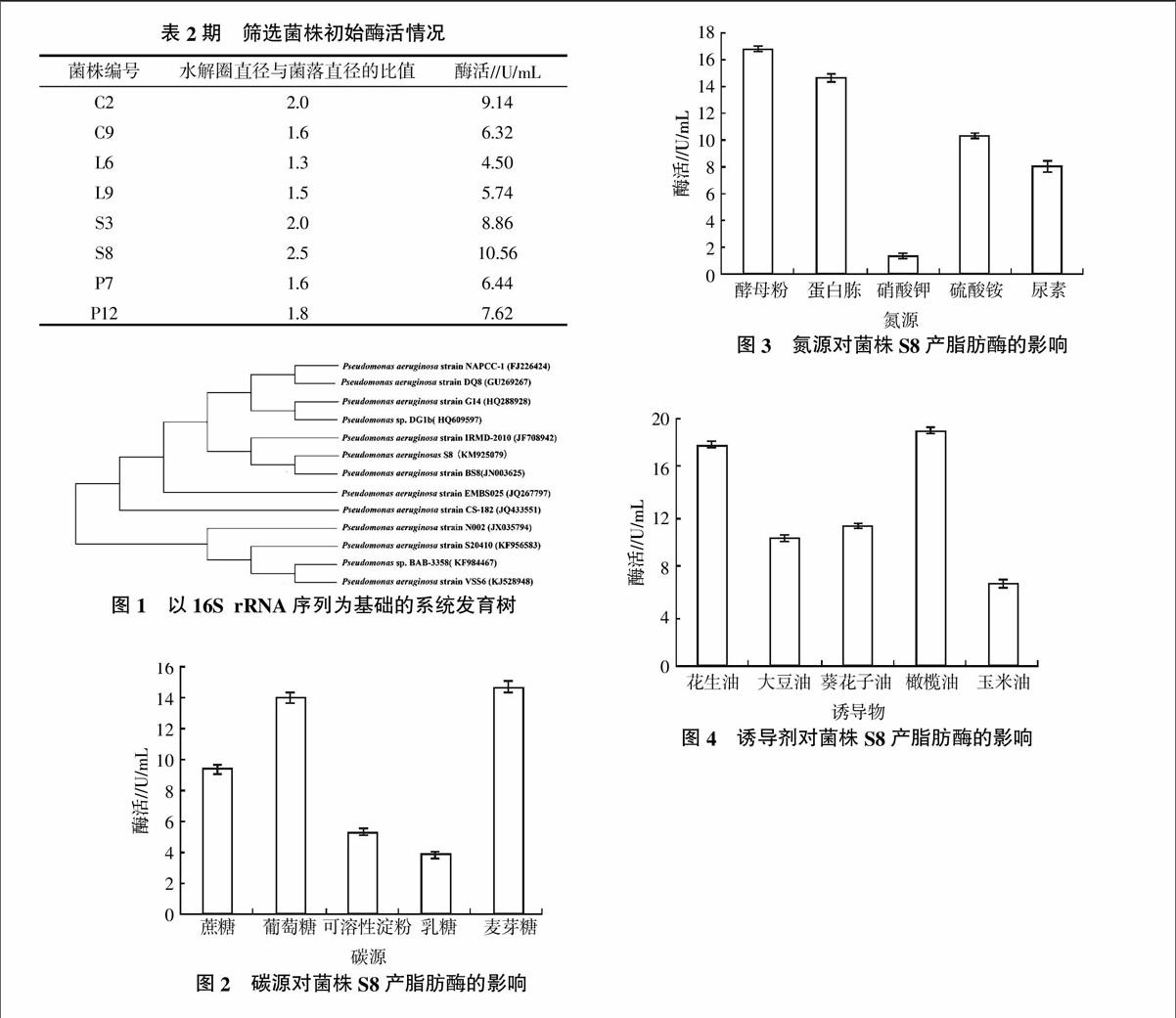
利用富集培养从被废油长期污染的16份土样中分离筛选得到87株能在初筛培养基中生长、有罗丹明B反应的菌株,挑取变色圈直径与菌落直径之比(Hc)大于1的28株菌落进行摇瓶发酵复筛,通过测定脂肪酶活,最终筛选到胞外脂肪酶活较高的8个菌株(表2),大部分初筛得到的菌株脂肪酶酶活较低,而菌株S8的初始酶活可达到10.56 U/mL,远高于其他菌株,有很好的研究价值。
2.2 产脂肪酶菌S8的分子生物学鉴定
对菌株S8进行16S rRNA分子鉴定,采用通用引物进行PCR扩增后,获得约1.4 kb的片段。将测序得到的序列提交至GenBank获得登录号:KM925079,通过BLAST与NCBI GenBank 数据库中已报道序列进行同源性比对,结果显示菌株S8与铜绿假单胞菌属(Pseudomonas aeruginosa)亲缘关系最近,同源性达到99%,构建系统发育树(图1)。
2.3 单因素试验结果
2.3.1 碳源对菌株S8产脂肪酶的影响 碳源是微生物生长的重要成分,对菌体的代谢产酶有重要影响。将菌株S8在添加浓度为0.2%的不同碳源的培养基中培养,分别测定发酵液脂肪酶酶活(图2)。由图2可知,以麦芽糖和葡萄糖为碳源时该菌株产酶能力较高,由于成本问题,选用葡萄糖作为最佳碳源。
2.3.2 氮源对菌株S8产脂肪酶的影响 以葡萄糖为碳源,将菌株S8在添加0.2%的不同氮源的培养基中培养,分别测定发酵液脂肪酶酶活(图3)。由图3可知,菌株在5种氮源发酵培养基中产脂肪酶酶活大小依次为酵母粉>蛋白胨>硫酸铵>尿素>硝酸钾,表明菌株S8对有机氮的利用优于无机氮,对铵态氮的利用优于硝态氮。因此,菌株S8的最佳有机氮源为酵母粉,无机氮源为硫酸铵。
2.3.3 诱导物对菌株S8产脂肪酶的影响 由于大多数水解酶类是诱导酶,很多微生物产生的脂肪酶也多属于诱导酶,因此在上述碳源和氮源的基础上添加20 g/L的不同种类的油脂作为脂肪酶的诱导剂,以考察油脂是否能有效的促进菌株S8产脂肪酶。由图4可知,20 g/L的橄榄油对菌株产酶最有利,最大酶活可以达到19.02 U/mL,而花生油的诱导效果仅次于橄榄油,从生产成本以及购买途径上考虑,选择花生油作为菌株S8产脂肪酶的诱导物。
2.3.4 发酵温度对菌株S8产脂肪酶的影响 培养温度对发酵来说是极其重要的,温度对菌体的生长和脂肪酶的产生的影响是各种因素的综合表现。调节发酵温度分别为20、25、30、35、40 ℃,研究不同发酵温度对菌株S8发酵产酶能力的影响(图5)。由图5可知,随着发酵温度的升高,脂肪酶的酶活增大,当温度达到30 ℃时,脂肪酶酶活最大,超过30 ℃酶活有所下降。因此,适宜菌株S8产脂肪酶的发酵温度为30 ℃。
2.3.5 初始pH对菌株S8产脂肪酶的影响 发酵培养基起始pH对微生物生长有着密切的关系,用稀盐酸或稀氢氧化钠将发酵培养基调至不同的pH,发酵培养后测定脂肪酶酶活,结果表明,初始pH控制在6.0~7.5较为合适,当pH在7.0时效果最佳(图6)。
2.3.6 接种量对菌株S8产脂肪酶的影响 将发酵培养基稀释至107个/mL,按发酵液体积的4%、6%、8%、10%、12%和14%接种于发酵培养基中,发酵结束后测定脂肪酶酶活(图7)。由图7可知,脂肪酶的酶活以10%接种量为最大,这是因为过高接种量使接种的细胞数目增多,发酵培养基中的营养相对不足,细胞的生长受到抑制,影响了脂肪酶的产生。
2.3.7 发酵时间对菌株S8产脂肪酶的影响 在以葡萄糖为碳源、酵母粉和硫酸铵为氮源、花生油为诱导物、初始pH为7.0、接种量为10%、发酵温度为30 ℃的条件下进行发酵,分别于24、48、72、96、120 h取发酵液测定脂肪酶酶活(图8)。由图8可知,发酵时间为72 h时,脂肪酶酶活最大,可达到26.2 U/mL。之后发酵时间再增加,由于菌体密度不断增大,发酵培养基中营养成分减少,发酵代谢产物增多,菌株生长进入平稳期,不利于脂肪酶的产生,因此,选择发酵时间为72 h。
2.4 正交试验优化结果
在单因素试验基础上,选取碳源葡萄糖、有机氮源酵母粉、无机氮源硫酸铵、温度和接种量5个因素,每个因素4个水平进行正交试验,以获得最有利菌株S8产脂肪酶的发酵参数。由正交试验结果(表3)可知,不同的试验因子对菌株S8发酵产脂肪酶影响次序为葡萄糖>酵母粉>硫酸铵>接种量>温度,最佳的发酵培养条件为葡萄糖8 g/L、酵母粉8 g/L、硫酸铵6 g/L、温度为30 ℃、接种量为9.0%。
3 小结与讨论
近年来,由于石化能源的储量逐年减少以及造成的污染日益严重,可再生能源受到人们的关注,尤其生物柴油以其低排放、可直接用于现有柴油机、无需进行结构的改造而备受青睐[10],目前生产生物柴油的方法很多,但用生物酶催化合成生物柴油具有反应条件温和、醇用量小、后处理简单、无污染物排放,并且原料油中的FFA能完全转化成甲酯等优点[11]。利用脂肪酶生产生物柴油在国内还处于初步研究阶段,而国外一些国家已经形成规模生产[12,13]。
本研究从东营胜利油田附近的土壤中成功分离出一株脂肪酶产生菌,经16S rRNA序列分析和系统发育分析鉴定,确定为铜绿假单胞菌,命名为Pseudomonas aeruginosa S8。利用单因素和正交试验方法确定了产脂肪酶的最佳发酵培养基为葡萄糖8 g/L、 酵母粉8 g/L、硫酸铵6 g/L,以花生油为诱导物;最佳发酵条件为起始pH 7.0、接种量9%、温度30 ℃、发酵时间72 h。通过筛选得到的脂肪酶菌株可作为今后工业化生产的出发菌株,并为脂肪酶基因的克隆、工程菌的构建奠定基础。
参考文献:
[1] CHO S S,PARK D J,SIMKHADA J R,et al.A neutral lipase applicable in biodiesel production from a newly isolated Streptomyces sp.CS326[J]. Bioprocess Biosyst Eng,2012,35(1-2):227-234.
[2] TAN T,LU J,NIE K,et al.Biodiesel production with immobilized lipase:A review[J]. Biotechnol Adv,2010,28(5):628-634.
[3] SHARMA R,CHISTI Y,BANERJEE U C.Production,purification,characterization, and applications oflipases[J]. Biotechnol Adv,2001,19(8):627-662.
[4] 彭立风,赵汝淇,谭天伟.微生物脂肪酶的应用[J].食品与发酵工业,2000,26(3):68-73.
[5] 张振乾,官春云.可催化生产生物柴油脂肪酶产生菌的筛选及其培养条件研究[J].中国油脂,2009,34(1):41-45.
[6] EISYED H,HAMED R R, KANTOUCH A. Enzyme-based feltproofing of wool[J].AATCC Rev,2002,2(1):25-28.
[7] 肖海群.脂肪酶产生菌的选育及其发酵条件的研究[D].南昌:南昌大学,2007.
[8] ABRAMIC M, LESCIC I,KORICA T,et al. Purification and properties of extracellular lipase from Sreptomyces rimosus[J]. Enzyme Microbial Technol,1999,25(6):522-529.
[9] 郄晓莎.低温脂肪酶高产菌株的筛选、鉴定及其发酵产酶条件的优化[D].河北保定:河北农业大学,2007.
[10] BEALL G H, PINCHNEY L R. Nanophase glass ceramics[J]. J Am Chem Soc,1999,82(1):5-16.
[11] 姜 楠,张 正.生物柴油的现状与发展前景[J].世界农业,2005(3):11-12.
[12] 李宁杨,孙佩慧,胡基埂,等.脂肪酶催化制备生物柴油的研究进展[J].中国生物工程杂志,2008,28(10):136-140.
[13] 舒正玉,杨江科,黄 瑛.生物柴油生产用脂肪酶资源及研发现状[J].湖北农业科学,2007,46(6):1027-1030.
